
Abstract
To investigate the physiological responses of plants to high root-zone temperature (HT, 35 °C) stress mitigated by exogenous glutathione (GSH), cucumber (Cucumis sativus L.) seedlings were exposed to HT with or without GSH treatment for 4 days and following with 4 days of recovery. Plant physiological variables, growth, and gene expression related to antioxidant enzymes and Calvin cycle were quantified. The results showed that HT significantly decreased GSH content, the ratio of reduced to oxidized glutathione (GSH/GSSG), chlorophyll content, photosynthesis and related gene expression, shoot height, stem diameter, as well as dry weight. The exogenous GSH treatment clearly lessened the HT stress by increasing the above variables. Meanwhile, HT significantly increased soluble protein content, proline and malondialdehyde (MDA) content as well as O2•− production rate, the gene expression and activities of antioxidant enzymes. The GSH treatment remarkably improved soluble protein content, proline content, antioxidant enzymes activities, and antioxidant enzymes related gene expression, and reduced the MDA content and O2•− production rate compared to no GSH treatment in the HT condition. Our results suggest that exogenous GSH enhances cucumber seedling tolerance of HT stress by modulating the photosynthesis, antioxidant and osmolytes systems to improve physiological adaptation.
Similar content being viewed by others

Introduction
Global warming is generally predicted to have a negative effect on plant growth and productivity due to the damaging effect of high temperature on plant development1,2. Under heat stress conditions, plants are subject to physical changes in their environments3 as well as reduction in performance of plant cell functions, including enzyme activity, membrane fluidity, protein complexes formation, chlorophyll synthesis, photosynthesis, respiration, and redox state4,5. Considering that global temperature continues to increase and there is an urgent need for the development of adaptation strategies to maintain agricultural yields, a better understanding of the influences of the high temperature on plant physiology and growth is required.
Cucumber (Cucumis sativus L.) is a thermophilic species which grows well in warmer condition, however it is intolerant to high temperature, especially in its root zone6. In order to fulfill the highly increasing demand of vegetables such as cucumber, greenhouse vegetable production systems have been rapidly developed in recent decades in China7. The greenhouse temperature can easily reach and exceed to high and intolerable level (above 38 °C) during the summer8. Opening the greenhouse windows is the common approach for reduction of the air temperature but not soil temperature, especially for soil which is covered with black films. Temperature in the root-zone is often above 30 °C, or even higher than 35 °C in hot seasons. High root-zone temperature (HT) may negatively affect plant physiology, morphology and growth9,10. But a comprehensive investigation of the physiological basis underlying cucumber plant responses to the HT stress is still lacking.
Photosynthesis is an important biological process that is highly sensitive to high temperature stress and is often inhibited before other cell functions are impaired11. The main reason might be that high temperature seriously suppresses the activities of ribulose-1,5-bisphosphate carboxylase/oxygenase (Rubisco) large subunit (RBCL), Rubisco small subunit (RBCS) and other related enzymes12, and leads to impairment of chlorophyll biosynthesis13. It has been reported that biotic and abiotic stress often induces the overproduction of reactive oxygen species (ROS) such as the superoxide radical (O2•−) and hydrogen peroxide (H2O2)14. Accumulation of ROS can damage lipids, proteins, nucleic acids, and carbohydrates, leading to destruction of plant cells15. ROS are scavenged by plant antioxidant defense systems which include non-enzymatic antioxidants (such as ascorbic acid, AsA; glutathione, GSH) and antioxidant enzymes (such as superoxide dismutase, SOD; catalase, CAT; ascorbate peroxidase, APX; glutathione reductase, GR; guaiacol peroxidase, G-POD)16,17. Proline accumulation has been reported during various stress conditions, including drought18 and heat19. Proline is important in cellular homeostasis (cell proliferation or cell death) and can act as a signaling molecule to modulate mitochondrial functions and trigger specific genes expression, which is essential for plant recovery from stresses20.
GSH and associated redox status have been reported to play an important role in cellular signaling pathways involved in many physiologic processes in plants under both optimal and stress conditions21. GSH is able to provide stress protection in several ways22,23. Reduced GSH acts as an antioxidant and is directly involved in reducing most ROS24. The ratio of reduced to oxidized glutathione (GSH/GSSG) redox couple is an indicator of the cellular redox status5,25. Some studies indicated that maintaining a high level of GSH or GSH/GSSG is important for the response to different stresses, such as drought tolerance in mung bean26, low temperature tolerance in loquat27, and isoproturon toxicity in wheat28. However, the role of GSH or GSH/GSSG levels in response to HT remains unknown.
In this study, a glasshouse experiment was conducted to test the HT and exogenous GSH effects on cucumber plant physiology and growth. We hypothesized that applying exogenous GSH to roots may increase heat tolerance by triggering the enzyme activities and gene expression level involved in photosynthesis and stress responses, as well as cellular redox homeostasis maintaining in plants. The specific objectives of this study were: (1) to investigate the effects of the HT on plant physiological variables; (2) to test whether exogenous GSH application can effectively enhance plant tolerance; and (3) to illustrate the potential adaption mechanism in HT stress. The information generated in this study could improve our understanding of HT tolerance of cucumber plants and would be useful for greenhouse and protected vegetation production system management.
Results
Effects of HT and exogenous GSH on plant growth
Compared to normal temperature treatment (NT), plant height, stem diameter, shoot fresh weight, and shoot dry weight of the HT treatment were significantly reduced by 14.8%, 8.1%, 38.1% and 32.6%, respectively, in the HT stress and by 24.0%, 12.6%, 33.5%, 37.7%, respectively, in the recovery period (Table 1; Fig. 1). High root-zone temperature plus exogenous GSH treatment (HT+GSH) significantly increased plant height, stem diameter, shoot fresh weight, and dry weight by 23.2%, 7.3%, 56.8% and 20.7% in the HT stress and by 29.2%, 7.7%, 32.0% and 25.7% in the recovery period, compared to the HT only treatment. Shoot water content was significantly decreased by 1.0% in the HT treatment compared to the NT treatment, however, there was no significant difference between them during the recovery period. HT+GSH treatment significantly increased the shoot water content by 2.7% in contrast with the HT treatment but showed no difference in the recovery period. There were no significant differences for any of the variables measured between the NT treatment and normal temperature plus exogenous GSH treatment (NT+GSH) during and after the HT stress. Shoot dry weight significantly decreased and shoot water content significantly increased for the HT+GSH treatment under the HT stress, and stem diameter, shoot fresh weight, and shoot dry weight were clearly decreased in the recovery period compared to the NT treatment.

Photographs of cucumber seedlings under 4 treatments, 4 days after the high root-zone temperature treatment, demonstrating effects of exogenous GSH on the response to high root-zone temperature stress.
Effects of HT and exogenous GSH on photosynthetic tolerance
In the present study, changes in the light and CO2 response curves of leaf photosynthetic rate (Pn) for different treatments were studied during and after the HT treatment (Fig. 2). Compared to the NT treatment, the light response curves showed that cucumbers experienced the strongest stress under the HT treatment, with Pn of only 6 μmol CO2 m−2s−1 under 2000 μmol m−2s−1 irradiance. The HT+GSH treatment enhanced Pn by a factor of 2.3 compared to HT. We derived the maximum leaf photosynthetic rate (Pmax), quantum yield (α), and the leaf photosynthetic rate under no irradiance (P0) from the light response curves using the rectangular hyperbolic equation29. Compared to the NT treatment, the HT treatment significantly decreased P0 (from 2.415 (NT) to 1.033 μmol CO2 m−2s−1 (HT)), Pmax (from 26.61 (NT) to 8.58 μmol CO2 m−2s−1 (HT)) and α (from 0.0794 (NT) to 0.0194 (HT)) (Table 2). HT+GSH significantly increased P0 (2.174 μmol CO2 m−2s−1), Pmax (19.20 μmol CO2 m−2s−1) and α (0.0556) compared to the HT treatment.

Changes of leaf photosynthetic rate (Pn) to light response curve and CO2 response curve for different treatments 4 days after the high root-zone temperature treatment, and 4 days recovery.
Data are means of three biological replications.
The CO2 response curve under the HT treatment showed the lowest Pn value among the treatments. The HT+GSH treatment had higher Pn than HT across CO2 levels (Fig. 2). In the HT treatment, we found that the maximum velocity of RuBP carboxylation by Rubisco (Vcmax) and the maximum potential rate of electron transport contributing to RuBP regeneration (Jmax) also dropped significantly compared to the NT treatment (from 47.30 (NT) to 20.42 μmol m−2s−1 (HT) and from 126.63 (NT) to 45.51 μmol m−2s−1 (HT), respectively). Similar to the light responses, the HT+GSH treatment increased the values to 40.73 μmol m−2s−1 for Vcmax and 97.81 μmol m−2s−1 for Jmax (Table 2).
In the recovery period, cucumber Pn was enhanced based on the light and CO2 response curves, and Pn in the HT+GSH treatment was increased more than in the HT treatment during the HT stress. As a result, there were no differences in P0 and α under all treatments, and Pmax, Vcmax and Jmax were slightly decreased under the HT treatment compared to the NT treatment. The HT+GSH treatment still slightly increased Pmax, Vcmax and Jmax compared to the HT treatment. There were no significant differences in P0, Pmax, α, Vcmax, and Jmax between the NT and NT+GSH in both treatment and recovery periods (Fig. 2 and Table 2).
Effects of HT and exogenous GSH on chlorophyll, carotenoid, soluble protein, and proline content
Compared to the NT treatment, the HT treatment decreased leaf chlorophyll and carotenoid content significantly by 5.9% and 4.8%, respectively (Table 3). Soluble protein and proline content were increased by 15.4% and 24.0% respectively under the HT treatment. The HT+GSH treatment increased chlorophyll, soluble protein, and especially proline content, which was stimulated by 51.8% compared to the HT treatment. There were no differences in soluble protein or proline content between the NT+GSH and NT treatments. Chlorophyll and carotenoid contents were enhanced significantly under the NT+GSH treatment relative to the NT treatment.
Effects of HT and exogenous GSH on antioxidant enzymes activities, MDA content, O2•− production rate, and GSH redox homeostasis
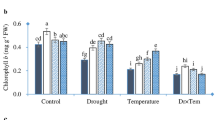
The activities of CAT, APX, G-POD, SOD and GR were stimulated by 13.0%, 25.2%, 35.4%, 16.6% and 14.4%, respectively, under the HT treatment, and by 20.3%, 40.1%, 54.2%, 7.8% and 28.2%, respectively, under the HT+GSH treatment compared to the NT treatment (Fig. 3). The HT+GSH treatment significantly increased the activities of APX, G-POD, and GR, but had no obvious effect on CAT and SOD activity relative to the HT treatment. Although the antioxidant enzyme activities under the NT+GSH treatment were slightly increased, there was no significant difference between the NT+GSH and NT treatments for of the variables except GR.

Effects of GSH application on catalase (CAT), ascorbate peroxidase (APX), superoxide dismutase (SOD), guaiacol peroxidase (G-POD) and glutathione reductase (GR) activities and their transcription levels changes in cucumber plants 4 days after the high root-zone temperature treatment.
Data represent mean ± SD (n = 3). Different letters indicate significant differences at P < 0.05 based on the Least Significant Difference test.
Malondialdehyde (MDA) and O2•−production were significantly increased by 60.6% and 79.9%, respectively, under the HT treatment, and by 23.2% and 30.2%, respectively, under the HT+GSH treatment, compared to the NT treatment (Fig. 4). There was no obvious difference in MDA content and O2•− production between the NT and NT+GSH treatments.

Effects of GSH application on MDA content and O2•− production rate of cucumber 4 days after the high root-zone temperature treatment.
Data represent the mean ± SD (n = 3). Different letters indicate significant differences at P < 0.05 based on the Least Significant Difference test.
The HT treatment remarkably decreased the level of GSH content and GSH/GSSG ratio in cucumber leaves compared to the NT treatment (Fig. 5). The HT+GSH treatment significantly enhanced the GSH content relative to the HT treatment, but had no significant difference compared to the NT or NT+GSH treatment.

Effects of GSH application on GSH content and GSH/GSSG of cucumber 4 days after the high root-zone temperature treatment.
Data represent the mean ± SD (n = 3). Different letters indicate significant differences at P < 0.05 based on the Least Significant Difference test.
Effects of HT and exogenous GSH on gene expression related to photosynthesis and antioxidant enzymes
The expression level of all the Calvin cycle enzyme genes investigated here were significantly up-regulated during the HT stress under the HT+GSH treatment compared to the HT treatment (except for fructose-1,6-bisphosphatase (FBPase) for which the increase was not significant) (Fig. 6), but all of the genes, to some extent, were down-regulated compared to the NT treatment. The gene expressions of RBCS, sedoheptulose-1,7-bisphosphatase (SBPase), and FBPase were significantly increased under the NT+GSH treatment compared to the NT treatment.

GSH application induced up-regulation of transcript levels for RCBL, RCBS, RCA, SBPase, FBPase and FBPaldolase 4 days after the high root-zone temperature treatment.
Data represent the mean ± SD (n = 3). Different letters indicate significant differences at P < 0.05 based on the Least Significant Difference test.
The transcription levels of antioxidant enzyme genes such as CAT, cAPX (cytoplasm ascorbate peroxidase), G-POD, Cu/Zn-SOD and GR were significantly increased under the HT treatment relative to the NT treatment (Fig. 3). HT+GSH considerably enhanced the expressions of CAT, cAPX, G-POD, and GR, with GR being induced the most compared to the HT treatment. Compared to the NT treatment, the expressions of CAT, cAPX, and GR were significantly increased under the NT+GSH treatment.
Discussion
As predicted, we found that the HT treatment significantly reduced plant height, stem diameter, shoot fresh weight, shoot dry weight, and shoot water content of cucumber, similar to some previous studies (e.g., Wahid et al.30). However, these reductions in plant growth were significantly alleviated by the addition of exogenous GSH (Table 1). The production of GSH under the HT stress is likely a critical mechanism for stress tolerance5,31 and has positive effects on plant growth and yield32.
Photosynthesis of cucumber plants was depressed under the HT treatment based on the light and CO2 response curves, which could be due to the reduction of chlorophyll content and carotenoid content at high temperatures that impaired the biosynthesis of photosynthetic pigments33. The application of exogenous GSH relieved the negative effects of the HT treatment on photosynthesis, not only during the HT treatment but also in the recovery period (Fig. 2). The HT+GSH treatment significantly increased chlorophyll and carotenoid content consistent with the results of previous studies27,34. The reduction of photosynthetic capacity was often accompanied by decreases in Pmax and α29, Vcmax and Jmax35. Inactivation or loss of Rubisco would reduce Vcmax while a reduction in Jmax is associated with the diminution of key regulatory enzymes such as SBPase and FBPase in the Calvin cycle36,37. Our results showed that Pmax, α, Vcmax and Jmax were significantly decreased in the HT treatment and increased again in the HT+GSH treatment, which suggested that the enzymes involved in RuBP carboxylation by Rubisco and RuBP regeneration limit photosynthesis under the HT stress. A similar result was found by Li et al.12 who revealed that the activity of enzymes involved in CO2 assimilation is higher at cucumber optimal growth temperature. In addition, expressions of Calvin cycle enzyme genes such as RBCL and RBCS were significantly enhanced under the HT+GSH treatment, indicating that up-regulation of Calvin cycle enzymes at the translational level could positively influence CO2 assimilation38. This is consistent with previous studies that found transcription factors play an important role in biotic and abiotic stress responses30,39. Jiang et al.24 also reported positive relationships between the expressions of Calvin cycle enzyme genes and the GSH/GSSG ratio, and an important role of reducing redox state on the stability of Calvin cycle enzymes.
Our results showed that the HT treatment increased MDA content and O2•− production rate in cucumber leaves, which was significantly decreased in the HT+GSH plants (Fig. 4). This is in accordance with previous findings that an increase in MDA level due to lipid peroxidation is one of the most common markers of cellular oxidative damage under the heat stress40. A higher GSH level reduces oxidative stress induced by heat stress32. Plants typically accumulate ROS under stress conditions. Tolerance to high temperature stress in plants is often associated with an increase in antioxidant enzyme activities41. In this study, we found that exogenous GSH increased the activities of CAT, APX, G-POD and GR under the HT treatment, which is consistent with the study of Nahar et al.32 who reported that oxidative damage effects are reduced by the GSH treatment during heat stress. Furthermore, transcription regulation of the antioxidant response (cAPX, G-POD and GR) was significantly increased under the HT+GSH treatment, which suggests that antioxidant adjustment in supra-optimal temperature is very important for plant adaptation to heat stress5. Huang et al.19 found that glucose application increases the transcriptions of some antioxidant enzymes under heat stress. The up-regulation of these genes contributed to an enhanced adaptation to high temperature stress in cucumber roots. Similarly, an exogenous GSH-induced improvement of activities of antioxidant enzymes reduces oxidative stress under various abiotic stress conditions26,27,28. Therefore, exogenous GSH regulates the accumulation of ROS and decreases membrane lipid peroxidation in the heat-stressed cucumbers through inducing antioxidant enzymes.
It is well established that proline accumulates in many plant species in response to environmental stress20. In this study, we found that proline was increased significantly under the HT treatment and even more when imposed in the HT+GSH treatment. These results indicated that the more accumulation of proline, the more stress tolerant for the plants. The reason could be that proline is an ROS scavenger and a molecular chaperone stabilizing the structure of proteins, thus helps protect cells from damage caused by the stress20. Similar results are reported in other plants such as mung bean32.
The accumulation of soluble protein, especially for the heat shock proteins during heat stress, contributes to stress tolerance in plants30. Our study demonstrated that the HT+GSH treatment significantly increased the soluble protein content of leaves under the HT stress, which is in agreement with a previous study that found protein protection plays an important role in maintaining high heat resistance in cucumber5.
The amelioration of exogenous GSH on the growth of cucumber under the HT treatment was effective (Fig. 1). The level of GSH has been shown to correlate with the adaptation of plants to extremes of temperature stresses15 and plants with a higher GSH/GSSG ratio possess higher stress-tolerance characteristics26. In this study, the HT+GSH treatment increased GSH accumulation and GSH/GSSG ratio compared to the HT treatment (Fig. 5). GR often plays an important role in the protection of plants from temperature stresses by preventing the oxidation of enzymes and membranes42. Indeed, higher GR activity was found with the application of exogenous GSH (Fig. 3), which may contribute to the high level of GSH and high GSH/GSSG ratio in the HT+GSH treatment compared to the HT treatment. Exogenous GSH application with drought26 and isoproturon toxicity28 also reduces these stresses by increasing GSH content and GSH/GSSG ratio.
In conclusion, we found that the stress of HT on cucumber seedlings was mostly relieved by the exogenous supplementation of GSH. Several mechanisms might be involved, including: (1) Maintaining high leaf water content, chlorophyll content, carotenoid content, and high activity of the Calvin cycle with the GSH application improved cucumber plant tolerance to the HT treatment, and led to high photosynthesis. GSH maintained osmotic balance by regulating soluble protein content and proline content thus improved the water status of plant leaves to resist the stress; (2) Exogenous GSH effectively eliminated HT-induced oxidative damage not only by increasing activities of antioxidant enzymes but also by improving transcription regulation of the antioxidant responses; and (3) Increases in GSH, enhanced GSH/GSSG ratio, or both, might be an obligatory event in the modulation of redox potential necessary for plants to adapt to the HT stress (Fig. 7). Such exogenous GSH to enhance HT tolerance mechanisms could be further exploited to improve our understanding of stress tolerance and the agricultural production especially in hot seasons.

A framework of the effects of exogenous GSH on the responses of high root-zone temperature stress to physiological variables, antioxidant enzymes, and gene expressions of cucumber seedlings.
Red arrow indicates the effects of HT treatment compared to the NT treatment. Blue arrow indicates the effects of HT+GSH treatment compared to the HT treatment. Up arrow means increased effect and down arrow means decreased effect.
Materials and methods
Plant material and treatments
The cucumber variety used in this study was Chunqiuwang NO. 2 selected by the Horticultural Research Institute of Shanghai Academy of Agricultural Sciences, China. Cucumber seeds were sown in Grodan blocks (10 cm × 10 cm × 6.5 cm) on February 12, 2015 in a well heated glasshouse. The seedlings were watered with half-strength Enshi nutrient solution43. The temperature in the glasshouse was maintained at 25 °C during the day and 18 °C at night. Plants grew under the glasshouse natural light. When the second true leaf was fully expanded, 4 seedlings were transplanted into each plastic container (27 cm × 40 cm × 12 cm). Nutrient solution was maintained full in the container, and a ventilation pump ensured the solution had enough oxygen. Meanwhile, a heater stick was fixed at the bottom of the container to control the solution temperature. The HT treatments were started when the fourth true leaf of plants was fully expanded. The treatments lasted 4 days, and measurements continued for 4 more days to capture the recovery period. The biggest leaves of each treatment were harvested after 4 days HT treatment, and the samples were frozen immediately in liquid nitrogen and stored at −80 °C for further analysis.
The experiment used a completely randomized design with three replications (plastic containers) for each treatment. Four treatments were included in this experiment: (1) normal temperature (NT): plants shoots grew in the normal glasshouse temperature, no heating for roots zone (about 22 °C with a range of 20–25 °C); (2) normal temperature plus exogenous GSH treatment (NT+GSH): added 0.25 mM GSH in nutrient solution one day before the treatments and maintained at 0.25 mM GSH in the nutrient solution during the treatment period; (3) high root-zone temperature treatment (HT): root-zone was heated to 35 ± 1 °C by heater stick; and (4) high root-zone temperature plus exogenous GSH treatment (HT+GSH): root-zone temperature was raised to 35 ± 1 °C and 0.25 mM GSH in nutrient solution was maintained.
Growth and shoot water content measurements
Plant height, stem diameter, shoot fresh weight were measured after 4 days of treatments and 4 days of recovery. Meanwhile, shoots of each treatment were harvested for the measurement of fresh weight, and then placed in the oven at 80 °C for 3 days to measure the dry mass. Shoot water content was calculated as (plant fresh weight - dry weight)/plant fresh weight. Five replicates were performed for each treatment.
Measurements of light response curve and CO2 response curve
Light and CO2 response curves were measured using a LI-6400 Portable Photosynthesis System (Li-Cor Inc., Lincoln, NE, USA) on the middle fully developed leaves of cucumber seedling after 4 days of treatments and 4 days of recovery. Irradiance levels were set at 0, 20, 50, 100, 200, 300, 500, 800, 1000, 1500 and 2000 μmol photon m−2s−1 for light response curve measurement. CO2 concentration was set at 400 μmol mol−1 with air temperature and relative humidity set at the greenhouse conditions. Leaves were allowed to acclimate to each irradiance level for about 2 min before reading and irradiance was increased successively from 0 to 2000 μmolm−2s−1.
CO2 concentrations were set at 0, 50, 100, 200, 400, 600, 800, 1000, 1200, 1500 and 2000 μmol mol−1 and irradiance levels were set at 1000 μmol m−2s−1 for CO2 response curve measurements. Leaves were allowed to acclimate to each CO2 level for about 2 min before reading. The first CO2 concentration was set at 400 μmol mol−1 which approached the greenhouse CO2 concentration and increased to 2000 μmol mol−1 and then successively decreased from 2000 to 0 μmol mol−1.
These light response curves and CO2 response curves were measured on each of the two leaves and replicated three times.
Measurements of chlorophyll content and carotenoid content
Leaf tissues (0.5 g) supernatant was extracted with 80% v/v acetone and absorbance was measured with a UV–visible spectrophotometer at 663, 645, and 470 nm. Total chlorophyll and carotenoid contents were calculated following the procedure described by Lichtenthaler and Wellburn44.
Measurements of O2•− producing rate in leaf extracts and MDA content
The O2•− production rate was measured by analyzing nitrite formation from hydroxylamine in the presence of O2•− 45. Each 0.5 g of frozen leaf segment was homogenized with 3 mL of 65 mM potassium phosphate buffer (pH 7.8) and centrifuged at 5,000 g for 10 min. The incubation mixture contained 0.9 mL of 65 mM phosphate buffer (pH7.8), 0.1 mL of 10 mM hydroxylamine hydrochloride, and 1 mL of the supernatant. After incubation at 25 °C for 20 min, 17 mM sulfanilamide and 7 mM R-naphthylamine were added to the incubation mixture. Ethylether in the same volume was added and centrifuged at 1,500 g for 5 min. The absorbance in the aqueous solution was read at 530 nm.
MDA content was determined according to Hodges et al.46. Leaf samples of 0.3 g were ground with 3 mL ice-cold 25 mM HEPES buffer (pH 7.8) containing 0.2 mM EDTA and 2% PVP. The homogenates were centrifuged at 4 °C for 20 min at 12,000 g, and the resulting supernatants were used for MDA analysis. Samples were mixed with 10% TCA containing 0.65% 2-thiobarbituric acid (TBA) and heated at 95 °C for 25 min. MDA content was calculated by correcting for compounds, other than MDA, that absorb at 532 and 600 nm, by subtracting the absorbance at 532 and 600 nm of a solution containing plant extract incubated without TBA from an identical solution containing TBA.
Measurements of proline and soluble protein
To determine the free proline level, 0.5 g of leaf sample from each group was homogenized in 3% (w/v) 5-sulfosalicylic acid, after which the homogenate was filtered through filter paper47. The mixture was heated at 100 °C for 1 h in a water bath after the addition of ninhydrin acid and glacial acetic acid. The reaction was then stopped in an ice bath. The mixture was extracted with toluene, and the absorbance of the fraction with toluene aspired from the liquid phase was read at 520 nm. The proline concentration was determined using a calibration curve48.
For biochemical assays, each 0.5 g sample of leaf material was homogenized in 3 mL 25 mM HEPES buffer (pH 7.8) containing 0.2 mM EDTA and 2% (w/v) polyvinylpyrrolidon. The homogenate was centrifuged for 20 min at 12,000 g and the supernatant obtained was used for enzyme analysis. All operations were performed at 0–4 °C. An aliquot of the extract was used to determine protein content according to Bradford49, using bovine serum albumin as the standard.
Glutathione assays
For the measurement of GSH and GSSG, plant leaf tissue (0.2 g) was homogenized in 2 mL of 2% metaphosphoric acid containing 2 mM EDTA and centrifuged at 4 °C for 10 min at 14,000 g. After neutralization with 0.5 M phosphate buffer (7.5), 0.1 mL of the supernatant was added to a reaction mixture containing 0.2 mM NADPH, 100 mM phosphate buffer (pH 7.5), 5 mM EDTA, and 0.6 mM 5,5′dithio-bis (2-nitrobenzoic acid). The reaction was started by adding 3 U of GR and was monitored by measuring the change in absorbance at 412 nm for 1 min. For the GSSG assay, GSH was masked by adding 20 μL of 2-vinylpyridine to the neutralized supernatant, whereas 20 μL of water was added for the total GSH assay. GSH level was obtained by subtracting the GSSG levels from the total level50.
Antioxidant enzyme activity assay
For the enzyme assays, 0.3 g of leaf sample was ground in 3 mL of ice-cold 25 mM HEPES buffer (pH 7.8) containing 0.2 mM EDTA, 2 mM AsA, and 2% PVP. The homogenates were centrifuged at 4 °C for 20 min at 12,000 g, and the supernatants were used for the determination of enzymatic activity. SOD activity was measured in a reaction mixture containing 50 mM phosphate buffer (pH 7.8), 0.1 mM EDTA, 13 mM methionine, 75 μM nitroblue tetrazolium (NBT), 2 μM riboflavin, and 50 μl enzyme aliquot51. One unit of SOD activity was defined as the amount of enzyme required to cause a 50% inhibition of the rate of p-nitro blue tetrazolium chloride reduction at 560 nm. The method of Cakmak & Marschner52, with some modifications, was used to determine the activity of G-POD. The reaction mixture contained 25 mM phosphate buffer (pH 7.0), 0.05% guaiacol, 1.0 mM H2O2 and 100 μl enzyme extract. The increase in absorbance at 470 nm caused by guaiacol oxidation (E = 26.6 mM cm−1) was used to determine the G-POD activity. CAT was assayed as described by Durner & Klessing53, and the activity was determined as a decrease in the absorbance at 240 nm for 1 min following the decomposition of H2O2. APX was measured according to Nakano & Asada54 by monitoring the rate of ascorbate oxidation at 290 nm. GR activity was measured according to Halliwell & Foyer55 based on the rate of decrease in the absorbance of NADPH at 340 nm.
RNA extraction and RT-PCR for gene expression analysis
To determine the effect of HT stress on the transcription levels of Calvin cycle enzyme genes and antioxidant enzyme genes and their changes after GSH application, we tested photosynthetic genes including those encoding ribulose-1,5-bisphosphate carboxylase/oxygenase activase (RCA), RBCL and RBCS involved in CO2 fixation. Other tested genes encode fructose-1,6-bisphosphate aldolase (FBPaldolase), which catalyze the conversion of two triose-3-phosphates into FBP. FBPase and SBPase catalyze the hydrolysis of FBP and SBP to Fru6P and Sed7P, respectively. Meanwhile, the tested antioxidant enzymes genes included Cu/Zn-SOD, G-POD, CAT, cAPX and GR. Total RNA was extracted using an RNA extraction kit (Axgen, Union City, CA) according to the supplier’s instructions. Contaminated DNA was removed with a purifying column. One microgram of total RNA was reverse-transcribed with a ReverTra Ace qPCR RT Kit (Toyobo, Japan) following the supplier’s recommendations. The gene-specific primers used for the amplification were determined on the basis of gene or EST sequences and are listed in Table S1, Supporting Information.
Quantitative real-time polymerase chain reaction (PCR) was performed with an iCycler iQ 96-well real-time PCR Detection System (Bio-Rad, Hercules, CA). PCR products were amplified using the SYBR Green PCR Master Mix (Applied Biosystems, Foster City, CA) in 25 μl of qRT-PCRs. The PCR conditions consisted of denaturation at 95 °C for 3 min, followed by 40 cycles of denaturation at 95 °C for 30 s, annealing at 58 °C for 30 s and extension at 72 °C for 30 s. To minimize sample variations, the mRNA expression of the target gene was normalized relative to the expression of the actin housekeeping gene. A quantification of mRNA levels was performed according to the method of Livak & Schmittgen56.
Statistical analysis
Analysis of variance (ANOVA) was conducted using the Statistical Analysis System (SAS version 9.3) (SAS Institute Inc., Cary, NC). Each value was presented as the mean ± standard deviation (SD), with a minimum of three replicates. Differences between treatment means were tested by the Least Significant Difference (LSD) method at α = 0.05 level of significance. The data were plotted using Origin 7.0 software (Origin Lab, Northampton, MA, USA).
Relationships between the leaf photosynthetic rate (Pn) and irradiance under four treatments were analyzed with a rectangular hyperbolic equation29:

where Pmax is the maximum leaf photosynthetic rate, α is quantum yield, I is irradiance, and P0 is the leaf photosynthetic rate when I = 0.
Relationships between Pn and intercellular CO2 concentration (Ci) of cucumber leaves were also analyzed by CO2 response curve. The maximum velocity of RuBP carboxylation by Rubisco (Vcmax) and the maximum potential rate of electron transport contributing to RuBP regeneration (Jmax) were estimated by non-linear regression techniques, based on the equations of Harley et al.57 and Manter & Kerrigan58.
Additional Information
How to cite this article: Ding, X. et al. Exogenous glutathione improves high root-zone temperature tolerance by modulating photosynthesis, antioxidant and osmolytes systems in cucumber seedlings. Sci. Rep. 6, 35424; doi: 10.1038/srep35424 (2016).
References
Allakhverdiev, S. I. et al. Heat stress: an overview of molecular responses in photosynthesis. Photosynth. Res. 98, 541–550 (2008).
Bita, C. E. & Gerats, T. Plant tolerance to high temperature in a changing environment: scientific fundamentals and production of heat stress-tolerant crops. Front. Plant Sci. 4, 273. 10.3389/fpls.2013.00273 (2013).
Ruelland, E. & Zachowski, A. How plants sense temperature. Environ. Exp. Bot. 69, 225–232 (2010).
Sage, R. F. & Kubien, D. S. The temperature response of C3 and C4 photosynthesis, Plant Cell Environ. 30, 1086–1106 (2007).
Li, H. et al. Growth temperature-induced changes in biomass accumulation, photosynthesis and glutathione redox homeostasis as influenced by hydrogen peroxide in cucumber. Plant Physiol. Biochem. 71, 1–10 (2013).
Tachibana, S. Effect of root temperature on the rate of water and nutrient absorption in cucumber cultivar and figleaf gourd. J. Japan. Soc. Hort. Sci. 55, 461–467 (1987).
Liang, X. S. et al. Effect of optimal daily fertigation on migration of water and salt in soil, root growth and fruit yield of cucumber (Cucumis sativus L.) in solar-greenhouse. PLoS ONE 9(1), e86975, 10.1371/journal.pone.0086975 (2014).
Ding, X. T. et al. Effects of heat shock on photosynthetic properties, antioxidant enzyme activity, and downy mildew of cucumber (Cucumis sativus L.). PLoS One 11(4), e0152429, 10.1371/journal.pone.0152429 (2016).
Malcolm, P. J., Holford, P., Barchia, I. & McGlasson, W. M. High and low root zone temperatures at bud-break reduce growth and influence dry matter partitioning in peach rootstocks. Sci. Hort. 171, 83–90 (2014).
Li, H. et al. Hydrogen peroxide mediates abscisic acid-induced HSP70 accumulation and heat tolerance in grafted cucumber plants. Plant, Cell and Environ. 37, 2768–2780 (2014).
Mathur, S., Agrawal, D. & Jajoo, A. Photosynthesis: response to high temperature stress. J. Photochem. Photobiol. B Biol. 137, 116–126 (2014).
Li, H. et al. The sub/supra-optimal temperature-induced inhibition of photosynthesis and oxidative damage in cucumber leaves are alleviated by grafting onto fig leaf gourd/luffa rootstocks. Physiol. Plantarum 152, 571–584 (2014).
Dutta, S., Mohanty, S. & Tripathy, B. C. Role of temperature stress on chloroplast biogenesis and protein import in pea. Plant Physiol. 150, 1050–1061 (2009).
Sharma, P., Jha, A. B., Dubey, R. S. & Pessarakli, M. Reactive oxygen species, oxidative damage, and antioxidative defense mechanism in plants under stressful conditions. J. Bot. 2012, 1–26 (2012).
Dai, A. H. et al. Cinnamic acid pretreatment enhances heat tolerance of cucumber leaves through modulating antioxidant enzyme activity. Environ. Exp. Bot. 79, 1–10 (2012).
Karimi, R., Ershadi, A., Abdolhossein, R. N. & Khanizadeh, S. Abscisic acid alleviates the deleterious effects of cold stress on ‘Sultana’ grapevine (Vitis vinifera L.) plants by improving the anti-oxidant activity and photosynthetic capacity of leaves. J. Hortic. Sci. Biotechnol. 10.1080/14620316.2016.1162027 (2016).
Gill, S. S. & Tuteja, N. Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant Physiol. Biochem. 48, 909–930 (2010).
Choudhary, N. L., Sairam, R. K. & Tyagi, A. Expression of delta(1)-pyrroline-5-carboxylate synthetase gene during drought in rice (Oryza sativa L.). Ind. J. Biochem. Biophys. 42, 366–370 (2005).
Huang, Y. W. et al. Glucose application protects chloroplast ultrastructure in heat-stressed cucumber leaves through modifying antioxidant enzyme activity. Biol. Plantarum 59, 131–138 (2015).
Szabados, L. & Savouré, A. Proline: a multifunctional amino acid. Trends Plant Sci. 15, 89–97 (2010).
Foyer, C. H. & Noctor, G. Ascorbate and glutathione: the heart of the redox hub. Plant Physiol. 155, 2–18 (2011).
Foyer, C. H. & Noctor, G. Redox homeostasis and antioxidant signaling: a metabolic interface between stress perception and physiological responses. Plant Cell 17, 1866–1875 (2005).
Foyer, C. H. & Noctor, G. Oxidant and antioxidant signaling in plants: a reevaluation of the concept of oxidative stress in a physiological context. Plant Cell Environ. 28, 1056–1071 (2005).
Jiang, Y. P. et al. Cellular glutathione redox homeostasis plays an important role in the brassinosteroid-induced increase in CO2 assimilation in Cucumis sativus. New Phytol. 194, 932–943 (2012).
Zhou, Y. H. et al. Brassinosteroids play a critical role in the regulation of pesticide metabolism in crop plants. Sci. Rep. 5, 10.1038/srep09018 (2015).
Nahar, K., Hasanuzzaman, M., Alam, M. M. & Fujita, M. Glutathione-induced drought stress tolerance in mung bean: coordinated roles of the antioxidant defence and methylglyoxal detoxification systems. AoB Plants 7, plv069, 10.1093/aobpla/plv069 (2015).
Wu, J. C., Sun, S. H., Ke, Y. T., Xie, C. P. & Chen, F. X. Effects of glutathione on chloroplast membrane fluidity and the glutathione circulation system in young loquat fruits under low temperature stress. Acta Hort. 887, 221–225 (2011).
Alla, M. M. N. & Hassan, N. M. Alleviation of isoproturon toxicity to wheat by exogenous application of glutathione. Pestic. Biochem. Physiol. 112, 56–62 (2014).
Hui, D. et al. Canopy radiation- and water-use efficiencies as affected by elevated [CO2]. Glob. Change Biol. 7, 75–91 (2001).
Wahid, A., Gelani, S., Ashraf, M. & Foolad, M. R. Heat tolerance in plants: An overview. Environ. Exp. Bot. 61, 199–223 (2007).
Kocsy, G. et al. Abiotic stress induced changes in glutathione and thioredoxin h levels in maize. Environ. Exp. Bot. 52, 101–112 (2004).
Nahar, K., Hasanuzzaman, M., Alam, M. M. & Fujita, M. Exogenous glutathione confers high temperature stress tolerance in mung bean (Vigna radiata L.) by modulating antioxidant defense and methylglyoxal detoxification system. Environ. Exp. Bot. 112, 44–54 (2015).
Havaux, M. Carotenoids as membrane stabilizers in chloroplasts. Trends Plant Sci. 3, 147–151 (1998).
Wang, F., Chen, F., Cai, Y., Zhang, G. & Wu, F. Modulation of exogenous glutathione in ultrastructure and photosynthetic performance against Cd stress in the two barley genotypes differing in Cd tolerance. Biol. Trace Elem. Res. 144, 1275–1288 (2011).
Zhou, Y. H., Lam, H. M. & Zhang, J. H. Inhibition of photosynthesis and energy dissipation induced by water and high light stresses in rice. J. Exp. Bot. 58, 1207–1217 (2007).
Allen, D. J. et al. An overnight chill induces a delayed inhibition of photosynthesis at midday in mango (Mangifera indica L.). J. Exp. Bot. 51, 1893–1902 (2000).
Ölcer, H., Lloyd, J. C. & Raines, C. A. Photosynthetic capacity is differentially affected by reduction in sedoheptulose-1,7-bisphosphatase activity during leaf development in transgenic tobacco plants. Plant Physiol. 125, 982–989 (2001).
Xia, X. J. et al. Brassinosteroids promote photosynthesis and growth by enhancing activation of Rubisco and expression of photosynthetic genes in Cucumis sativus. Planta 230, 1185–1196 (2009).
Yan, J. et al. De novo transcriptome sequencing and gene expression profiling of spinach (Spinacia oleracea L.) leaves under heat stress. Sci. Rep. 6, 10.1038/srep19473 (2016).
Savicka, M. & Škute, N. Effects of high temperature on malondialdehyde content, superoxide production and growth changes in wheat seedlings (Triticum aestivum L.). Ekologija 56, 26–33 (2010).
Sairam, R. K., Srivastava, G. C. & Saxena, D. C. Increased antioxidant activity under elevated temperature: a mechanism of heat stress tolerance in wheat genotypes. Biol. Plant. 43, 245–251 (2000).
Mohan, J. R., Burke, J. & Orzech, K. Thermal dependence of the apparent Km of glutathione reductases from three plant species. Plant Physiol. 93, 822–824 (1990).
Yu, J. Q. & Matsui, Y. Effects of root exudates of cucumber (Cucumis sativus) and allelochemicals on ion uptake by cucumber seedlings. J. Chem. Ecol. 23, 817–827 (1997).
Lichtenthaler, H. K. & Wellburn, A. R. Determinations of total carotenoids and chlorophylls a and b of leaf extracts in different solvents. Biochem. Soc. Trans. 11, 591–592 (1983).
Elstner, E. F. & Osswald, W. Mechanism of oxygen activation during plant stress. In. Oxygen and Environmental Stress in Plants. P. Roy. Soc. Edinb. B. 102, 131–154 (1994).
Hodges, D. M., DeLong, J. M., Forney, C. F. & Prange, R. K. Improving the thiobarbituric acid-reactive-substances assay for estimating lipid peroxidation in plant tissues containing anthocyanin and other interfering compounds. Planta 207, 604–611 (1999).
Koca, H., Bor, M., Ozdemir, F. & Türkan, I. The effect of salt stress on lipid peroxidation, antioxidative enzymes and proline content of sesame cultivars. Environ. Exp. Bot. 60, 344–351 (2007).
Nostar, O., Ozdemir, F., Bor, M., Turkan, I. & Tosun, N. Combined effects of salt stress and cucurbit downy mildew (Pseudoperospora cubensis Berk. and Curt. Rostov.) infection on growth, physiological traits and antioxidant activity in cucumber (Cucumis sativus L.) seedlings. Physiol. Mol. Plant P. 83, 84–92 (2013).
Bradford, M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 72, 248–254 (1976).
Rao, M. V. & Ormrod, D. P. Ozone exposure decreases UVB sensitivity in a UVB-sensitive flavonoid mutant of Arabidopsis. Photochem. Photobiol. 61, 71–78 (1995).
Giannopolitis, N. & Ries, S. K. Superoxide dismutase I. Occurrence in higher plants. Plant Physiol. 59, 309–414 (1977).
Cakmak, I. & Marschner, H. Magnesium deficiency and high light intensity enhance activities of superoxide dismutase, ascorbate peroxidase, and glutathione reductase in bean leaves. Plant Physiol. 98, 1222–1227 (1992).
Durner, J. & Klessing, D. F. Salicylic acid is amodulator of tobacco and mammalian catalases. J. Biol. Chem. 271, 28492–28502 (1996).
Nakano, Y. & Asada, K. Hydrogen peroxide is scavenged by ascorbate specific peroxidase in spinach chloroplasts. Plant Cell Physiol. 22, 867–880 (1981).
Halliwell, B. & Foyer, C. H. Ascorbic acid, metal-ions and the superoxide radical. Biochem. J. 155, 697–700 (1976).
Livak, K. J. & Schmittgen, T. D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 25, 402–408 (2001).
Harley, P. C., Loreto, F., Marco, G. D. & Sharkey, T. D. Theoretical considerations when estimating the mesophyll conductance to CO2 flux by analysis of the response of photosynthesis to CO2 . Plant Physiol. 98, 1429–1436 (1992).
Manter, D. K. & Kerrigan, J. A/Ci curve analysis across a range of woody plant species: influence of regression analysis parameters and mesophyll conductance. J. Exp. Bot. 55, 2581–2588 (2004).
Acknowledgements
We thank Ms. Ping Wang for her technical assistance in sampling and measurements and Dr. Jennifer Cartwright and Danial Hassani for polishing the English language. This work was supported by the National Key Technology Support Program (2014BAD08B01) and the Agricultural Basic Research of the Shanghai Agriculture Commission (2–1, 2014), and the Developmental Plan of the Shanghai Agriculture Commission (1–42, 2015). The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
Author information
Authors and Affiliations
Contributions
Conceived and designed the experiments: D. Huang and J.Y. Performed the experiments: Y.J. and X.D. Analyzed the data: Y.J. and X.D. Contributed reagents/materials/analysis tools: L.H. and Q.Z. Wrote the paper: X.D., D. Hui and D. Huang.
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Electronic supplementary material
Rights and permissions
This work is licensed under a Creative Commons Attribution 4.0 International License. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in the credit line; if the material is not included under the Creative Commons license, users will need to obtain permission from the license holder to reproduce the material. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/
About this article

Cite this article
Ding, X., Jiang, Y., He, L. et al. Exogenous glutathione improves high root-zone temperature tolerance by modulating photosynthesis, antioxidant and osmolytes systems in cucumber seedlings. Sci Rep 6, 35424 (2016). https://doi.org/10.1038/srep35424
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/srep35424
This article is cited by
-
Exogenous glutathione protected wheat seedling from high temperature and water deficit damages
Scientific Reports (2024)
-
Evaluation of salt and root-zone temperature stresses in leafy vegetables using hydroponics as a clean production cultivation technique in northeastern Brazil
Horticulture, Environment, and Biotechnology (2024)
-
Manipulation of light spectrum is an effective tool to regulate biochemical traits and gene expression in lettuce under different replacement methods of nutrient solution
Scientific Reports (2023)
-
The effects of exogenously applied antioxidants on plant growth and resilience
Phytochemistry Reviews (2023)
-
The Response of Triticum aestivum Treated with Plant Growth Regulators to Acute Day/Night Temperature Rise
Journal of Plant Growth Regulation (2022)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
