
Abstract
In this study we isolated 109 Enterococcus faecalis from chicken faecal samples in 6 provinces of China to investigate the prevalence and transmission mechanism of the bacitracin resistance locus bcrABDR in E. faecalis. Thirty-seven bcrABDR-positive E. faecalis were detected with 26 different PFGE clusters. The MLST of 14 positive strains belonged to ST16 and we also detected three new sequence types. S1-PFGE analysis indicated that the locus was located on plasmids presenting different sizes, with the most prevalent size being ~50 kb (13/37). Sequence analysis revealed that 17 out of the 37 strains harbored a 5400-bp central region, in which locus bcrABDR was bracketed by two ISEnfa1 of the same orientation. Two types of bcrABDR alleles, differing in around 10% of their sequence were found. In silico analysis showed that bcrABDR is present in a variety of bacteria including the chicken commensal Enterococcus cecorum. Our results indicate that the use of bacitracin at farms might trigger the emergence and spread of the bacitracin resistance determinant bcrABDR among human bacterial pathogens. The finding of bcrABDR in the chicken commensal E. cecorum indicates that farm animals microbiota can be an important reservoir of resistance genes with relevance for human health.
Similar content being viewed by others


Introduction
Bacitracin is a peptide antimicrobial that inhibits the cell wall biosynthesis by binding to the lipid undecaprenyl pyrophosphate (UPP) and impeding its dephosphorylation. UPP is the precursor of the undecaprenol monophosphate that serves as a membrane-associated carrier for sugar-peptide units in the peptidoglycan synthesis. After completion of the cycle, UPP is dephosphorylated to form undecaprenol monophosphate (UP) to start another synthetic cycle1. By binding UPP, bacitracin reduces the amount of available UP, stopping the synthesis of peptidoglycan, which finally leads to bacterial death.
It has been reported that bacitracin can be an effective antibiotic to treat patients who are infected with vancomycin-resistant enterococci (VRE)2,3. Nevertheless, this antibiotic is also used as animal growth promoter in China, and this wide use of the bacitracin for non-therapeutic procedures might compromise its utility for human health. Indeed, some recent reports have shown that high-level bacitracin resistance (MIC ≥ 256 μg/ml) in enterococci is becoming frequent and widespread4. Several bacitracin resistance mechanisms have been described in Gram-negative and Gram-positive bacteria5,6. In E. faecalis, a primary resistance mechanism has been reported to be the presence of the bcrABDR cluster, composed by the bcrABD operon and its regulatory gene bcrR7. The bcrABD operon encodes a putative heterodimeric ATP-binding cassette (ABC) transporter BcrAB that has been proposed to mediate the active efflux of bacitracin8. In addition, BcrD is a putative undecaprenol kinase that would convert UPP to UP. Overproduction of the undecaprenol kinase increases the levels of UP, which can overcome the low levels of UPP when the pathway is blocked by bacitracin9. Finally, BcrR has been identified as a bacitracin able sensor to regulate the expression of the operon bcrABD10.
In E. faecalis, the bcrABDR cluster has been sporadically detected11, but little is known about its prevalence and genetic environment. Our aim is to investigate the prevalence of the bcrABDR cluster in E. faecalis isolates obtained from animal samples from six different provinces in China in the aim of determining whether or not the wide use of bacitracin in farms may render a selection of strains carrying this resistance determinant, which might compromise the use of bacitracin in human therapy. In addition, we have explored the different hierarchical levels (from genes to clones) at which selection of resistance may operate in the aim of defining the elements involved in the allodemic12 spread of this resistance determinant, and its consequences for human health.
Results
The bacitracin resistance locus bcrABDR is widespread in Enterococcus faecalis isolates from chickens
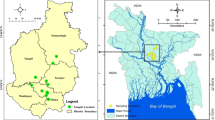
109 E. faecalis from 11 farms in 6 provinces of China (Table S1) were studied to determine the incidence and mechanisms of bacitracin resistance associated to farming activities. Among them, 37 (33.9%) strains displayed high-level resistance (MIC ≥ 256 μg/ml) to bacitracin. PCR analysis demonstrated that all the 37 isolates carried bcrABDR. In addition to presenting bacitracin resistance, all the bcrABDR-positive isolates were resistant to erythromycin, tetracycline, and streptomycin. Additionally, most of them were resistant to florfenicol and kanamycin (Fig. 1).These data indicate that bacitracin resistance, associated to the presence of bcrABDR is highly prevalent in E. faecalis strains isolated from chicken at farms in China.

The information of bcrABDR-positive strain.
The Figure shows epidemiological, phylogenetical and antibiotic resistance information of the strains analyzed in the current article. Strain EF21, EF24 and EF176 carry the locus bcrABDR highly similarity with the respect gene reported in plasmid pTW9 (NC_014726); ST683, ST684 and ST 685 were new ST types summation in this study; The size for locus bcrABDR positive plasmid is indicated by underline; The size for conjugative plasmid is indicated by black. NA, not available; UN, Hunan; GX, Guangxi; GD, Guangdong; ZJ Zhejiang; HN, Hainan; BAC, bacitracin; STR, streptomycin; ERY, erythromycin; TET, Tetracycline; FFC, florfenicol; KAN, kanamycin.
The acquisition of bcrABDR is polyclonal in E. faecalis
The 37 bcrABDR-positive E. faecalis isolates displayed different PFGE patterns and belonged to 26 clusters designated A to Z (Fig. 1). Cluster A, F, N, S contained 8, 2, 3, and 2 isolates respectively, and the other clusters contained a single strain each. MLST results showed a diversity of STs of the 37 isolates, with 14 strains belonging to ST 16. We also detected three new ST types, including ST683, ST684 and ST685 (Fig. 1).
Different transferrable plasmids are involved in the spread of bcrABDR in E. faecalis
Seven plasmids carrying bcrABDR were successfully transferred to a susceptible recipient strain through conjugation experiments. When compared with the recipient strain, the bacitracin MICs of the seven transconjugants increased at least 8-fold. In addition to bacitracin, co-transfer of resistance, erythromycin, streptomycin and kanamycin, were also observed in these seven transconjugants. Further, five of them conferred as well resistance to tetracycline and two transferred florfenicol resistance (Table S3). Altogether, these results highlight that bcrABDR-mediated bacitracin resistance is present in multi-resistance plasmids, supporting that co-selection by one or another of the antibiotics to which the plasmids confer resistance might be in the basis of their high prevalence.
To further characterize the plasmids carrying the bcrABDR operon, S1-PFGE followed by Southern blot using a bcrB-specific probe was performed for all the isolates containing bcrABDR. As shown in (Fig. S1), only a single plasmid hybridized with the bcrB-specific probe for each of the analyzed strains. The plasmids harboring the bcrABDR operon presented sizes ranging from 40 kb to 147 kb, with ~50kb being the most the most frequent size (13/37) (Figs 1 and S1).
To gain further insight into the mechanisms mediating bcrABDR spread, one of the plasmids (pEF123) was fully sequenced as described in methods. As shown in (Fig. S2), bcrABDR is surrounded in pEF123 by two ISEnfa1 insertion sequences, a structure also observed for the bcrABDR-containing plasmids pXD5 and pTW9. Notably, several other plasmids that do not contain the bcrABDR operon also presented ISEnfa1, indicating that this IS might be critical elements for the acquisition of genetic material and the evolution of E. faecalis plasmids.
The genetic environment surrounding bcrABDR is not always the same in conjugative plasmids
The regions surrounding the bcrABDR operon were determined by primer walking. Three different genetic environments (designated as type I–III) were detected in the 7 transconjugants. In the type I genetic organization, the locus bcrABDR was flanked by two copies of the insertion sequence ISEnfa1 located in the same orientation (JEF105, JEF123) and the locus bcrABDR was oriented in the opposite direction of ISEnfa1. To determine the stability of this structure, inverse PCR was performed using the primersP13/P14. Sequence analysis of the PCR production revealed that it contained one intact ISEnfa1, supporting that bcrABDR might form a circular intermediate through the recombination of the two copies of ISEnfa1. Figure 2 The inverse PCR was also performed on all the bcrABDR-positive strains, and 17 of them gave a positive result. Together with previously published works13, our results further support that the formation of circular intermediates may mediate the transfer of bcrABDR. In the type II organization, ISEnfa1 was only located upstream the locus bcrABDR (JEF7, JEF8, JEF127 and JEF129) without any ISEnfa1 located downstream. In the type III organization we could not find the insertion sequence ISEnfa1 neither upstream nor downstream bcrABDR (JEF174). Although it is tempting to propose that these results suggest a route for the evolution and spread of bcrABDR from bacteria not harboring ISEnfa1 towards strain carrying one ISEnfa1 copy, ending in those that carry two ISEnfa1 copies, in which the circular intermediate can be found, the opposite situation might also happens if loss of ISEnfa1 occur through abortive recombination events.

Genetic environment of the locus bcrABDR on the plasmid JEF123.
The genetic environment of bcrABDR from the E. faecalis EF123 analyzed in the study is shown in comparison with E. faecalis TX0645 genomic, E. faecalis B302 genomic, Streptococcus pyogenes NGAS322 genomic, Enterococcus faecium plasmid pXD5, Staphylococcus aureus NRS2 ctg7180000000010 genomic sequences. The positions and orientations of the genes are indicated by arrows, with the direction of transcription shown by the arrowhead. Regions of homology are shaded in gray. The whole ISEnfa1-bcrABDR-ISEnfa1 segment is 5400 bp in length. The 8 bp direct target site duplication sequence (5-TCTTTTCT-3) is boxed. The black arrows indicated the positions and orientations of the inverse PCR primers P13/P14.
The origin of plasmid-encoded bcrABDR is polyphyletic in E. faecalis
From the 37 bcrABDR-positive isolates, the four genes in the locus could be amplified just in three of the isolates with the primers previously described. For the other 34 isolates the genes bcrA and bcrD could not be detected using these primers, so new primers for these genes were designed according to the sequence of the plasmid pEF123 determined in this study by using these novel primers (Table S2), a new allelic variant of the bcrABDR operon was found. The amino acid sequences of BcrA, BcrB, BcrD and BcrR in the two alleles presented similarities of 90.8%, 88.9%, 88.3% and 94.1% respectively. Further, the Type II allele presents at least three different subtypes that were similar among them, but displayed some few consistent different SNPs (Fig. 3). Independent Blast searches using the two different allelic forms of the bcrABDR operon were performed. By means of these approaches, we found that both types of sequences are present at the NCBI database. The firstly described bcrABDR allele (hereafter dubbed as Type I), was found and identity of 99% in pTW9 and pJM01, whereas the bcrABDR sequence present in pEF123 (Type II), with a 99% identity was found in the plasmids pXD5 and R17. It has been discussed that the low variability of antibiotic resistance genes found in the mobile gene elements of bacterial pathogens might be the consequence of a founder effect14. The first resistance gene to spread will prevail and impede the entrance of other resistance elements rendering the same phenotype. Here we detect that two bcrABDR alleles (or maybe more if we consider the Type II subtypes as different alleles) are present in bacitracin resistant E. faecalis indicating that in this case at least two and likely four, different events might account for the acquisition of resistance. This situation may happen when there exists a close ecological relationship between the donor and the recipient of the resistance element, which allows these transfer events to occur frequently and being efficiently selected, likely as the consequence of bacitracin use in chicken farms.

Phylogenetic relationship of bcrABDR clusters analyzed in the present work.
The figure shows the phylogenetic relationships among the different bcrABDR alleles studied in the present work. As shown, two major branches (Type I and Type II) can be distinguished, with the Type II allele presenting also three different subtypes. This population structure indicates that the acquisition of bcrABDR by E. faecalis has a polyphyletic origin. Each black line in the Figure represents a SNP in comparison with the consensous bcrABDR sequence.
Enterococcus cecorum can be a reservoir of bacitracin resistance in chicken
To ascertain the distribution of bcrABDR, the sequence of the cluster and the surrounding ISEnfa1 was searched at NCBI DNA database. In addition to the aforementioned plasmids, Type II bcrABDR was found to be present in the genome of Streptocccus pyogenes NGAS322 as well as in the genomes of different strains of E. cecorum (a chicken commensal with some virulent members) isolates15,16,17, in occasions surrounded by ISEnfa1, in other occasions without IS present around. In addition, the genomes of E. cecorum also contain upstream and downstream sequences nearly identical to those found in pEF123 and other bcrABDR-containing plasmids. All of these will be powerful to improve Enterococcus cecorum can be a reservoir of bacitracin resistance in chicken. Although the whole-genome sequence methods used in this study did not allow distinguishing between chromosomal and plasmidic DNA. Since a route of transmission from E. cecorum towards E. faecalis or vice versa cannot be tracked, we cannot ascertain which is the origin of bcrABDR; however our results indicate that the commensal chicken microbiota can be a reservoir of transferrable bacitracin resistance that may end in relevant human pathogens as E. faecalis.
Discussion
In this study, we investigated the prevalence of the locus bcrABDR in E. faecalis isolated from 6 provinces in China. Our study showed that resistance to bacitracin, associated to the presence of plasmids containing the bcrABDR cluster was highly prevalent and associated with resistance to other antimicrobials, especially erythromycin, streptomycin, kanamycin, florfenicol and tetracycline. The polyclonal phenomenon result in E. faecalis indicated that different clones are involved in the spread of bacitracin resistant E. faecalis isolates containing the bcrABDR cluster. Persistence and co-selection of the bcrABDR cluster could be promoted under antibiotic selective pressure, when these classes of antimicrobials were used either as feed additives or to cure or prevent bacterial infections in animals18. When looking at the different elements, from genes to clones that may be involved in the spread of this resistance, we found that at least two different alleles of the bcrABDR cluster are present in different plasmids, suggesting the existence of multiple events of acquisition (and fixation) of this resistance determinant by E. faecalis from its (still unknown) original host. After these acquisition events, the spread of bcrABDR has been guided by its incorporation in different plasmids, which are also spread among different clonal complexes.
The inspection of the bcrABDR flanking regions showed three different organizations, in most cases, two intact ISEnfa1 flanked the bcrABDR cluster and inverse PCR shows that they can form bcrABDR-containing circular intermediates through homologous recombination, which suggests that IS-mediated recombination may account for the transfer of the bcrABDR cluster between different genetic platforms. Interestingly the structure ISEnfa1-bcrABDR-ISEnfa1 is also observed in other bacterial species, which can further suggest that the form of the circular intermediate is the main manner for the transmission of the locus bcrABDR (Fig. 2). For the other two types of genetic environments, in which bcrABDR is not flanked by ISEnfa1 the mechanisms involved in its acquisition and spread are not known at the moment.
All these results indicated that multiple levels of selection regulate the spread of bcrABDR among E. faecalis populations. The fact that this operon presented in other species13,19 shows that inter-species transmission is involved in the spread of this element. In this regard, it is particularly relevant that the bcrABDR operon is found in E. cecorum, which is the dominant enteric commensal of adult chicken and contributes to the gut microbiota of several avian species. This indicated that the commensal microbiota of farming animals can be a reservoir of antibiotic resistance that can be transferred to human pathogens. Indeed, MLST typing revealed that 14 of 37 bcrABDR-positive strains belonged to the high-risk clone ST16, which has been reported to be associated with the prevalence of the multi-resistance gene cfr in E. faecalis and VRE20,21 and is commonly found in animals, humans, and the environment22. The presence of ST16 in both poultry and faecal samples of healthy humans had been observed in Portugal, Denmark, New Zealand, Japan, Thailand, and the USA23 a situation that might suggests that E. faecalis can spread between animals and humans through the food chain. Finally, it is important to highlight that PCR-based epidemiological analyses must be taken into consideration that allelic forms of a given determinants can be present in the population, which are not detected by using the standard conditions (as in the case of Type II allele of bcrABDR. These situations may underscore the estimations on the prevalence of some resistance genes in bacterial populations).
Materials and Methods
Bacterial strains and susceptibility testing
A total of 109 E. faecalis were isolated from 181 faecal swabs of healthy chickens. The samples were collected from 11 farms in six provinces of China (Guangdong, Hainan, Guangxi, Hunan, Jiangxi and Zhejiang) in May 2014 (Table S1). The susceptibilities of bcrABDR-positive isolates to bacitracin and to other antimicrobials (tetracycline, gentamycin, kanamycin, erythromycin, streptomycin, and florfenicol) were determined by the agar dilution method following the guidelines of the Clinical and Laboratory Standards Institute (CLSI)24,25. E. faecalis ATCC 29212 was used as a quality control strain. Bacterial DNA was extracted using a DNA extraction kit (Omega, USA) following the manufacturer’s instructions. The presence of bcrABDR was determined by PCR and further sequencing of the amplicons. Primer walking was performed to analyze more in detail the different bcrABDR and surrounding structures in the selected strains. The primers used in this study were presented in Table S2.
Molecular typing
Pulsed-field gel electrophoresis (PFGE) was performed using the CHEF-MAPPER System (Bio-Rad Laboratories) as described previously26 to analyze the clonality of the E. faecalis strains carrying bcrABDR. PFGE clusters were analyzed with Bio Numerics (Applied Maths, Sint-Martens-Latem, Belgium). Dendrograms were generated using the Dice similarity coefficient, and the analogical values to categorize identical PFGE types cut-off was fixed at 90%. Further determination of clonality was performed by Multi Locus Sequence Typing (MLST), following the protocol described at http://efaecalis.mlst.net.
Conjugation and transformation analysis
Conjugation and transformation experiments of the bcrABDR positive strains were performed as described previously, using E. faecalis JH2-2 (Rifr) as the recipient strain27,28. Putative transconjugants and transformants were selected on brain heart infusion agar containing 25 μg/ml fusidic acid, 50 μg/ml rifampicin and 100 μg/ml bacitracin. bcrABDR positive transconjugants and transformants were detected by PCR, using the primers described in Table S2.
Plasmid characterization
To estimate the size of the plasmids harboring bcrABDR in the bcrABDR-positive strains (37 isolates and 7 transconjugants), whole-cell DNA was separated by PFGE after treatment with S1 nuclease (TaKaRa). Afterwards, the plasmids carrying bcrABDR were identified by Southern blot using a bcrB probe.
A bcrB-positive plasmid from transconjugant JEF123 was extracted using the Qiagen plasmid maxi kit (Qiagen, Valencia, CA). Sequencing was performed using the Miseq platform (Illumina, San Diego, CA) with a 500-bp paired-end library, and assembled by SOAPdenovo29. The gaps between the contigs were closed by PCR and respective amplicons were sequenced. Gene prediction and annotation were performed using the RAST tools30. The sequence comparison and map generation was performed using BLAST (http://blast.ncbi.nlm.nih.gov) and BRIG31.
Nucleotide sequence accession number
The nucleotide sequence of the plasmid pEF123 containing bcrABDR, has been deposited in GenBank under the accession number KX579977.
Additional Information
How to cite this article: Chen, M.-Y. et al.Multilevel selection of bcrABDR-mediated bacitracin resistance in Enterococcus faecalis from chicken farms. Sci. Rep. 6, 34895; doi: 10.1038/srep34895 (2016).
References
Chalker, A. F. et al.The bacA gene, which determines bacitracin susceptibility in Streptococcus pneumoniae and Staphylococcus aureus, is also required for virulence. Microbiology 146 (Pt 7), 1547–1553, 0.1099/00221287-146-7-1547 (2000).
Mondy, K. E., Shannon, W. & Mundy, L. M. Evaluation of zinc bacitracin capsules versus placebo for enteric eradication of vancomycin-resistant Enterococcus faecium. Clin Infect Dis 33, 473–476, 10.1086/321895 (2001).
Tran, T. T. et al.Oral Bacitracin: A Consideration for Suppression of Intestinal Vancomycin-Resistant Enterococci (VRE) and for VRE Bacteremia From an Apparent Gastrointestinal Tract Source. Clin Infect Dis 60, 1726–1728, 10.1093/cid/civ130 (2015).
Soares-Santos, V., Barreto, A. S. & Semedo-Lemsaddek, T. Characterization of Enterococci from Food and Food-Related Settings. J Food Prot 78, 1320–1326, 10.4315/0362-028X.JFP-14-419 (2015).
El Ghachi, M., Bouhss, A., Blanot, D. & Mengin-Lecreulx, D. The bacA gene of Escherichia coli encodes an undecaprenyl pyrophosphate phosphatase activity. J Biol Chem 279, 30106–30113, 10.1074/jbc.M401701200 (2004).
Dintner, S. et al.Coevolution of ABC Transporters and Two-Component Regulatory Systems as Resistance Modules against Antimicrobial Peptides in Firmicutes Bacteria. J. Bacteriology 193, 3851–3862, Doi 10.1128/Jb.05175-11 (2011).
Manson, J. M., Keis, S., Smith, J. M. & Cook, G. M. Acquired bacitracin resistance in Enterococcus faecalis is mediated by an ABC transporter and a novel regulatory protein, BcrR. Antimicrob Agents Chemother 48, 3743–3748, 10.1128/AAC.48.10.3743-3748.2004 (2004).
Gebhard, S. et al.Identification and characterization of a bacitracin resistance network in Enterococcus faecalis. Antimicrob Agents Chemother 58, 1425–1433, 10.1128/AAC.02111-13 (2014).
Shaaly, A., Kalamorz, F., Gebhard, S. & Cook, G. M. Undecaprenyl pyrophosphate phosphatase confers low-level resistance to bacitracin in Enterococcus faecalis. J. Antimicrob Chemother 68, 1583–1593, Doi 10.1093/Jac/Dkt048 (2013).
Gebhard, S., Gaballa, A., Helmann, J. D. & Cook, G. M. Direct stimulus perception and transcription activation by a membrane-bound DNA binding protein. Mol Microbiol 73, 482–491, 10.1111/j.1365-2958.2009.06787.x (2009).
Matos, R., Pinto, V. V., Ruivo, M. & Lopes, M. D. S. Study on the dissemination of the bcrABDR cluster in Enterococcus spp. reveals that the BcrAB transporter is sufficient to confer high-level bacitracin resistance. Int J Antimicrob Agents. 34, 142–147, DOI 10.1016/j.ijantimicag.2009.02.008 (2009).
Baquero, F., Coque, T. M. & Canton, R. Allodemics. Lancet Infect Dis 2, 591–592 (2002).
Wang, X. M. et al.Characterization of a multidrug resistance plasmid from Enterococcus faecium that harbours a mobilized bcrABDR locus. J Antimicrob Chemother 70, 609–611, 10.1093/jac/dku416 (2015).
Martinez, J. L. Bottlenecks in the transferability of antibiotic resistance from natural ecosystems to human bacterial pathogens. Front Microbiol 2, 265, 10.3389/fmicb.2011.00265 (2011).
Dolka, B., Chrobak-Chmiel, D., Makrai, L. & Szeleszczuk, P. Phenotypic and genotypic characterization of Enterococcus cecorum strains associated with infections in poultry. BMC Vet Res 12, 129, 10.1186/s12917-016-0761-1 (2016).
Dolka, B. et al.Draft Genome Sequences of Two Commensal Enterococcus cecorum Strains Isolated from Chickens in Belgium. Genome Announc 3, 10.1128/genomeA.01108-15 (2015).
Borst, L. B., Suyemoto, M. M., Scholl, E. H., Fuller, F. J. & Barnes, H. J. Comparative genomic analysis identifies divergent genomic features of pathogenic Enterococcus cecorum including a type IC CRISPR-Cas system, a capsule locus, an epa-like locus, and putative host tissue binding proteins. PLoS One 10, e0121294, 10.1371/journal.pone.0121294 (2015).
Thumu, S. C. & Halami, P. M. Acquired resistance to macrolide-lincosamide-streptogramin antibiotics in lactic Acid bacteria of food origin. Indian J Microbiol 52, 530–537, 10.1007/s12088-012-0296-5 (2012).
Han, X. et al.Functional analysis of a bacitracin resistance determinant located on ICECp1, a novel Tn916-like element from a conjugative plasmid in Clostridium perfringens. Antimicrob Agents Chemother 59, 6855–6865, 10.1128/AAC.01643-15 (2015).
Diaz, L. et al.Transferable plasmid-mediated resistance to linezolid due to cfr in a human clinical isolate of Enterococcus faecalis. Antimicrob Agents Chemother 56, 3917–3922, 10.1128/AAC.00419-12 (2012).
Kudo, M., Nomura, T., Yomoda, S., Tanimoto, K. & Tomita, H. Nosocomial infection caused by vancomycin-susceptible multidrug-resistant Enterococcus faecalis over a long period in a university hospital in Japan. Microbiol Immunol 58, 607–614, 10.1111/1348-0421.12190 (2014).
Novais, C. et al.Spread of multidrug-resistant Enterococcus to animals and humans: an underestimated role for the pig farm environment. J Antimicrob Chemother 68, 2746–2754, 10.1093/jac/dkt289 (2013).
Freitas, A. R., Novais, C., Ruiz-Garbajosa, P., Coque, T. M. & Peixe, L. Clonal expansion within clonal complex 2 and spread of vancomycin-resistant plasmids among different genetic lineages of Enterococcus faecalis from Portugal. J Antimicrob Chemother 63, 1104–1111, 10.1093/jac/dkp103 (2009).
Clinical and Laboratory Standards Institute.Performance Standards for Antimicrobial Susceptibility Testing; 23rd Informational Supplement M100–S23. CLSI, Wayne, PA, USA 2013a.
Clinical and Laboratory Standards Institute.Performance Standards for Antimicrobial Disk and Dilution Susceptibility Tests for Bacteria Isolated From Animals; Approved Standard-Fourth Edition. CLSI document VET01-A4. CLSI, Wayne, PA, USA, 2013b.
Kim, S. Y. et al.Characterization of Enterococcus spp. from human and animal feces using 16S rRNA sequences, the esp gene, and PFGE for microbial source tracking in Korea. Environ Sci Technol 44, 3423–3428, 10.1021/es903282p (2010).
Huys, G., D’Haene, K., Collard, J. M. & Swings, J. Prevalence and molecular characterization of tetracycline resistance in Enterococcus isolates from food. Appl Environ Microbiol 70, 1555–1562 (2004).
Liu, Y. et al.First report of the multidrug resistance gene cfr in Enterococcus faecalis of animal origin. Antimicrob Agents Chemother 56, 1650–1654, 10.1128/AAC.06091-11 (2012).
Luo, R. et al.SOAPdenovo2: an empirically improved memory-efficient short-read de novo assembler. Gigascience 1, 18, 10.1186/2047-217X-1-18 (2012).
Aziz, R. K. et al.The RAST Server: rapid annotations using subsystems technology. BMC Genomics 9, 75, 10.1186/1471-2164-9-75 (2008).
Alikhan, N. F., Petty, N. K., Ben Zakour, N. L. & Beatson, S. A. BLAST Ring Image Generator (BRIG): simple prokaryote genome comparisons. BMC Genomics 12, 402, 10.1186/1471-2164-12-402 (2011).
Acknowledgements
This study was supported by the Program for Changjiang Scholars and Innovative Research Team in University of Ministry of Education of China (Grant No. IRT13063), National Natural Science Foundation and Natural Science Foundation of Guangdong Province, China (Grant No. U1201214), the National Natural Science Foundation (Grant No. 31272609).
Author information
Authors and Affiliations
Contributions
M.C. performed experiments, analyzed the data and wrote the manuscript; H.L., J.D. and R.W. performed experiments. F.L. and J.M. analyzed the data and edited the manuscript; J.S. designed the experiments, analyzed the data, and edited the manuscript. X.L. and Y.L. coordinated the whole project.
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Electronic supplementary material
Rights and permissions
This work is licensed under a Creative Commons Attribution 4.0 International License. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in the credit line; if the material is not included under the Creative Commons license, users will need to obtain permission from the license holder to reproduce the material. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/
About this article

Cite this article
Chen, MY., Lira, F., Liang, HQ. et al. Multilevel selection of bcrABDR-mediated bacitracin resistance in Enterococcus faecalis from chicken farms. Sci Rep 6, 34895 (2016). https://doi.org/10.1038/srep34895
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/srep34895
This article is cited by
-
Longitudinal study on the effects of growth-promoting and therapeutic antibiotics on the dynamics of chicken cloacal and litter microbiomes and resistomes
Microbiome (2021)
-
Defining and combating antibiotic resistance from One Health and Global Health perspectives
Nature Microbiology (2019)
-
Prevalence, resistance pattern, and molecular characterization of Staphylococcus aureus isolates from healthy animals and sick populations in Henan Province, China
Gut Pathogens (2018)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
