
Abstract
Sex hormones fluctuate during the menstrual cycle. Evidence from animal studies suggests similar subtle fluctuations in hippocampal structure, predominantly linked to estrogen. Hippocampal abnormalities have been observed in several neuropsychiatric pathologies with prominent sexual dimorphism. Yet, the potential impact of subtle sex-hormonal fluctuations on human hippocampal structure in health is unclear. We tested the feasibility of longitudinal neuroimaging in conjunction with rigorous menstrual cycle monitoring to evaluate potential changes in hippocampal microstructure associated with physiological sex-hormonal changes. Thirty longitudinal diffusion weighted imaging scans of a single healthy female subject were acquired across two full menstrual cycles. We calculated hippocampal fractional anisotropy (FA), a measure sensitive to changes in microstructural integrity, and investigated potential correlations with estrogen. We observed a significant positive correlation between FA values and estrogen in the hippocampus bilaterally, revealing a peak in FA closely paralleling ovulation. This exploratory, single-subject study demonstrates the feasibility of a longitudinal DWI scanning protocol across the menstrual cycle and is the first to link subtle endogenous hormonal fluctuations to changes in FA in vivo. In light of recent attempts to neurally phenotype single humans, our findings highlight menstrual cycle monitoring in parallel with highly sampled individual neuroimaging data to address fundamental questions about the dynamics of plasticity in the adult brain.
Similar content being viewed by others

Introduction
Accumulating evidence on the effects of estrogen outside of the reproductive system supports an influence of estrogen on brain structure and function1,2,3,4. Several rodent studies have shown strong effects of estrogen on adult neurogenesis5, electrophysiological and structural properties of the brain6, interactions with main neurotransmitter systems7, regulation of gene transcription8,9, and behavior2. Considering the prominent sexual dimorphism seen in many neuropsychiatric disorders, such as major depressive disorder10, endogenous sex hormones may play an essential role in modulating human brain states and mood. Epidemiologic studies report an increased risk for depressed mood when endogenous sex hormone levels fluctuate or decline rapidly from elevated levels, such as during the menopausal transition11,12 or early postpartum13. Changes in hippocampal function and morphology have been reported during depressive episodes in young adulthood14 and these findings have been replicated in different samples15,16. Given the high density of estrogen receptors in the hippocampus17, this region is of particular relevance for exploring whether subtle endogenous sex hormone fluctuations can induce neuroplastic change in humans.
Estrogen has been shown to elicit neurotrophic effects including promoting synaptogenesis and strengthening glia-structure in the rodent hippocampus18. In humans, the influence of estrogen on hippocampal morphology has been investigated by focusing on the changes that occur during puberty, when sex hormone levels rise substantially. During this period, increased estrogen levels have been shown to parallel greater volume in hippocampal regions19,20. Indirect support for estrogen-induced neuroplastic processes in the hippocampus stems from research in Turner syndrome, a condition of severe estrogen deficiency in females that is the result of a complete or partial absence of the X-chromosome21. Neuroimaging findings show reduced hippocampal grey and white matter in the presence of this condition21. Investigating the effect of endogenous estrogen on hippocampal neuroplasticity in humans in vivo is a relatively recent scientific endeavor. Most existing data are either derived from male-to-female comparisons22, cross-sectional studies23, or clinical populations21,24. Within-subject designs are challenging and therefore rare. Yet, such an approach may be more suitable to detect potential effects of subtle estrogen changes in the female brain.
The menstrual cycle provides a natural experimental set-up to investigate whether physiological sex hormonal fluctuations influence brain morphology. Most prior studies exploring the link between estrogen and female brain plasticity have focused on comparisons of women either on or off oral contraception25, sometimes reporting menstrual cycle effects as secondary findings26. Hormonal contraceptive use has been associated with increased grey matter in cortical regions25 and changes in cerebral white matter structure27. During the natural menstrual cycle, Protopopescu and colleagues found grey matter volume changes in the right anterior hippocampus in the high estrogen phase28. However, monitoring of the menstrual cycle in this study was based on self-reports, the authors did not assess sex hormone levels in saliva or blood, and the analysis was limited to two assessments across the menstrual cycle28. These caveats point to the most common challenge scientists face when trying to determine menstrual cycle phase, namely, self-reports are unreliable. It is estimated that only about 40% of women correctly report the time of their last menstrual bleeding when the time of recall was 3 weeks after bleeding cessation or longer29. More accurate monitoring methods include the assessment of body temperature and the detection of sex hormone concentration in blood and urine. However, these methods also have limitations, particularly when they are not applied in combination. Non-invasive daily recording of body temperature is highly influenced by day-to-day variability in the measurement procedure itself, cycle variability and the effects of illness, medication, diet, and fluctuations in sleeping patterns30. Blood sampling at a single time point to assess menstrual cycle phase is insufficient because the menstrual cycle is characterized by high estrogen levels before ovulation (follicular phase) and has a second blunted peak after ovulation and before the onset of menses (luteal phase). Multiple blood sampling time points are thus needed for accurate menstrual cycle monitoring. In addition, confirming ovulation, for instance by assessing lutropin (LH) levels in urine, seems advisable given that an anovulatory cycle can randomly occur in women with otherwise good menstrual cycle health31.
When accurate menstrual cycle tracking is performed, the resulting analysis of ovarian hormone fluctuation and brain structure often yields interesting data: several rodent studies have shown that estrogen has neurotrophic and neuroprotective effects on grey and white matter structure across the estrous cycle32,33.
So far, there is no direct method available to quantify white matter microstructural changes in the human brain in vivo. However, it is possible to assess parameters that are sensitive to alterations in white matter microstructure. Diffusion weighted imaging (DWI) is a MRI technique that measures the directionality of molecular water diffusion34 and enables non-invasive observation of structural reorganization in the human brain35. As axons provide natural barriers for water diffusion, the diffusion profile within a certain brain region can be used to infer local structural properties35. This method has been used successfully to detect white matter changes characteristic of axonal damage and demyelination34 and reorganization associated with functional recovery after brain injury36. Scalar measures of DWI include fractional anisotropy (FA, the most frequently used metric of the diffusion tensor), which quantifies how strongly directional the local tissue is34. A decrease in FA reflects decreased anisotropic diffusion that can indicate white matter damage37,38. Yet, FA is also sensitive to microstructural changes in fiber density and myelination39. Axial diffusion along the primary axis and radial diffusion perpendicular to the primary diffusion axis can be investigated within each FA cluster to specify diffusion directionality in more detail40,41. Both diffusion properties have been associated with different biological underpinnings: radial diffusion (RD) with myelin changes42 and axial diffusion (AD) with axonal damage34. Understanding the relative contribution of these parameters to FA values allows for a more in depth interpretation40,41 of potential estrogen effects on human white structure microstructure across the menstrual cycle.
In this study, we aim to explore the influence of endogenous estrogen on hippocampal white matter measures. Investigating the influence of subtle physiological estrogen fluctuations on structural connectivity in health can reflect on the capacity of the human brain to adapt to the environment. In contrast, treatment and scar effects can bias data obtained from cross-sectional studies in clinical populations. The present work, a heavily sampled longitudinal dataset with detailed information on hormonal status, could prove a powerful means to detect subtle brain changes in synchrony with the menstrual cycle.
Materials and Methods
Participant
We acquired 30 longitudinal scans of a single, 32-year old, Caucasian female with a documented history of regular menstrual cycles (mean = 28 ± 1 day). The subject was physically healthy, with no history of any psychiatric, neurological, or chronic illnesses. The subject was right-handed, of normal weight (BMI 20.2), and free of medication, including no use of contraceptives or antidepressants (i.e., antidepressant-naïve). She was a non-smoker, with no history of alcohol use, drug abuse, pregnancy, or period of breast-feeding. The participant was screened using the structured clinical interview for DSM to rule out any Axis I major mental disorders43 and Axis II personality disorders44. Any potential subclinical manic, depressive and anxiety symptoms were excluded by administration of the Hamilton Rating Scale for Depression (HAM-D)45, the Mood Spectrum self-report (MOODS-SR), and the State-Trait anxiety inventory (STAI). The subject was recruited through the institute’s volunteer database and provided written informed consent to participate. The study protocol was approved by the local ethics board of the University of Leipzig (EK-No.: 246-2009-09112009) and carried out in accordance with the Declaration of Helsinki.
Hormone assays
We collected fasting blood samples to determine serum hormone levels of estrogen (pmol/l), progesterone (nmol/l), and LH (U/l) immediately prior to every MRI scan. Blood samples were analyzed at the Institute for Laboratory Medicine of the University Hospital Leipzig by the fully automated Roche Cobas® system (Roche, Basel, Switzerland). All samples were measured within one plate: intra-assay coefficients of variance (ICV) were within 3.2–6% for estrogen, 2.3–5.2% for progesterone, and 1.6–2% for LH. Ovulation was further confirmed by detecting LH surge, using urine ovulation tests (Diagnostik Nord GmbH hLH-K20 hLH Kassettentest).
MRI data acquisition
MRI images were acquired every second or third day in two separate scanning sessions to cover two full menstrual cycles. Data from scanning session one was collected from January 6, 2012, to February 3, 2012 (16 scans in total) starting with cycle day 15 in the first cycle; to cycle day 21 in the second cycle. Therefore the first scan session bridges two menstrual cycles, covering one full menstrual cycle. The second scanning session spanned from October 31, 2012 to November 30, 2012 (16 scans in total) starting with cycle day 25 until cycle day 3. The time course of scanning according to the subject’s menstrual cycle is depicted in chronological order in Figures 1 and 2(C). Scanning sessions began on a different day of the menstrual cycle and were acquired to different seasons in order to control for potential scanner-drift artifacts and seasonal effects, respectively. Further, every scan was collected at the same time of the day (7.30 am) to control for circadian rhythm effects. Two scans were excluded: one because of missing hormone data and one because of a technical problem with DWI acquisition.

Endogenous ovarian sex hormone fluctuation.
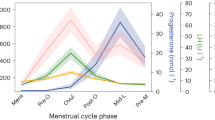
Characteristic patterns of serum estradiol (red line), progesterone (black line) and LH (green line) levels are displayed across two menstrual cycles (follicular phase highlighted in grey). The depicted data represent days of cycle with corresponding z-standardized, single hormone values, matching the single scan time points in chronological order. As expected, estradiol shows a first prominent peak in the periovulatory phase, followed by a second smaller peak in the late luteal phase. LH surges after the peak of estrogen, shortly before ovulation. Progesterone levels are low during the follicular phase and high during the luteal phase.

Estrogen-modulated FA differences and time course across the menstrual cycle in the bilateral hippocampus.
Threshold-Free Cluster Enhancement (TFCE) voxel-wise FA correlation with respective estrogen levels in hippocampal ROI-masks (left (panel A) and right (panel B) hippocampus) are displayed. Red voxels outlined with black, superimposed on respective t-values, correspond to significant FWE-corrected results (p < 0.05). In panel C, FA (left) and RD (right) values from significant clusters, respectively peak voxel, are extracted and plotted versus estrogen levels (in red) across the menstrual cycles assessed.
All images were acquired on a 3-Tesla Magnetom Verio scanner (Siemens, Erlangen, Germany) equipped with a 32-channel head coil. During each scanning session, diffusion data were acquired using a whole-brain, gradient-echo echo-planar imaging (EPI) sequence. The following parameters were used: 72 slices, slice thickness = 1.7 mm, no intersection gap, field of view = 220 mm2, TR = 13.8 s, TE = 100 ms, A/P phase encoding direction, 67 diffusion directions, transversal orientation, fat saturation, b = 0, 1000 s/mm2, bandwidth = 1346 Hz/pixel, GRAPPA parallel imaging and acquisition matrix = 128 × 128. The reconstruction matrix was the same as the acquisition matrix, and voxels were 1.7 mm isotropic. The total DWI scanning time was 16 minutes.
In addition, high-resolution T1-weighted images to test for grey matter structural changes were acquired using a whole-brain, three-dimensional Magnetization-Prepared Rapid Gradient Echo (MPRAGE) sequence. The Alzheimer’s Disease Neuroimaging Initiative (ADNI) standard protocol was used with the following parameters: TI = 900 ms, TR = 2300 ms, TE = 2.98 ms, flip angle = 9°, A/P phase encoding direction, bandwidth = 238 Hz/pixel, image matrix = 256 × 240, 176 partitions, field of view = 256 × 240 × 176 mm3, sagittal orientation, no fat suppression and average voxel size 1 × 1 × 1 mm3. The acquisition time was 9 minutes.
Diffusion data analysis
Diffusion data were analyzed with FSL version 5.0.8 using the FMRIB’s software library (www.FMRIb.ox.ac.uk/fsl). Briefly, the following standard processing pipeline was applied before creating voxel wise maps of diffusion parameters: (1) eddy current correction to correct for gradient-coil distortions and small head motions, (2) removal of non-brain tissue using the Brain Extraction Tool (fractional threshold = 0.25), and (3) local fitting of the diffusion tensor at each voxel using DTIfit (FMRIB’s Diffusion Toolbox v2.0 (FDT)46).
Next, FA images were processed with FSL’s diffusion toolkit47,48. First, all images were nonlinearly aligned to the most representative FA image out of all images and transformed into 1 × 1 × 1 mm MNI152 standard space. Volumetric FA images were minimally smoothed (σ = 1 mm) and subsequently used in permutation-based nonparametric statistical analyses. The nonlinear warp derived from FA co-registration was also used to align the volumetric images of axial diffusivity (AD) and radial diffusivity (RD). This approach was used to examine the relative contribution of AD and RD to potential hormone-dependent changes in FA values post hoc.
Structural data analysis
MPRAGE structural data was analyzed with FSL-VBM49 (http://fsl.fmrib.ox.ac.uk/fsl/fslwiki/FSLVBM), an optimized voxel-based morphometry (VBM) protocol50 carried out with FSL version 5.0.8. First, structural images were reoriented, brain-extracted and grey matter-segmented before being registered to the MNI 152 standard space using non-linear registration (www.fmrib.ox.ac.uk/analysis/techrep). The resulting images were averaged and flipped along the x-axis to create a left-right symmetric, study-specific grey matter template. Second, all native grey matter images were non-linearly registered to this study-specific template and “modulated” to correct for local expansion (or contraction) due to the non-linear component of the spatial transformation. The modulated grey matter images were then smoothed with an isotropic Gaussian kernel with a sigma of 3 mm. Finally, voxel wise general linear model was applied using permutation-based non-parametric testing, correcting for multiple comparisons across space.
Statistical analysis
Based on previous work linking the menstrual cycle to grey matter changes and changes in functional connectivity51 in the hippocampus, we chose this structure as our region of interest (ROI). Hand drawn ROIs for right and left hippocampal white matter were derived from the thresholded (FA > 0.2) mean FA image and hippocampal segmentation of the Harvard-Oxford Subcortical Structural Atlas52 implemented in FSLView (FSL)47. To validate our hand drawn mask, we also generated intensity-based, atlas-defined anatomical masks of the bilateral hippocampus by using the Harvard Oxford Subcortical Structural Atlas, which were thresholded at 0.2 to account for white matter.
We correlated volumetric FA from the bilateral hippocampal ROIs with respective hormone levels using a nonparametric permutation-based approach (5000 permutations). We set the statistical threshold at a voxel-level at p < 0.05, after family-wise error (FWE) correction for multiple comparisons using threshold-free cluster enhancement53. The same approach was used for the structural data using anatomical atlas-defined hippocampal masks from the Harvard Oxford Subcortical Structural Atlas.
Regions identified in the previous step were subsequently used as masks to extract FA, AD and RD values for plotting and for calculating partial correlations with respective hormone values using SPSS Statistics 22 (p < 0.05). These analyses were corrected for set of scan sessions as a covariate of no interest, as both sessions were acquired 9 months apart.
Results
Hormone assessment
Hormone analyses confirmed typical patterns for estrogen, progesterone and LH levels characteristic of the ovulatory, follicular, and luteal phases of the menstrual cycle, across all cycles in the study. A plot of the measured hormone fluctuation across the menstrual cycles can be found in Fig. 1.
Correlations within region of interest
To determine hippocampal microstructural differences related to specific endogenous hormone fluctuations during the menstrual cycle, volumetric FA values in the bilateral hippocampal ROIs were correlated with respective hormone levels. Estrogen levels were significantly correlated with volumetric FA in the bilateral hippocampus (right peak voxel: 22 −32 −7, t = 5.75 p = 0.0016; left peak voxel: −21 −40 1, t = 5.36 p = 0.0232; Fig. 2A,B). No such correlation was found for progesterone.
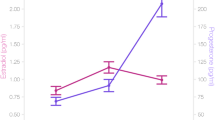
To assess menstrual cycle-related hippocampal grey matter density changes, we performed a permutation-based nonparametric statistical analysis using FSL-VBM49. We found a significant correlation of estrogen levels with grey matter density in the left hippocampus (peak voxel: -20 -28 -8, t = 3.86, p = 0.0118; Fig. 3A), but not in the right hippocampus. To visualize hippocampal grey matter density changes in comparison to hippocampal FA changes across the menstrual cycle, we extracted mean grey matter values of the left hippocampus and plotted it against estrogen level and mean FA values of the left hippocampus (Fig. 3B, for details on how to access this dataset, see supplementary information).

Estrogen-modulated grey matter differences in the left hippocampus.
Threshold-Free Cluster Enhancement (TFCE) voxel-wise grey matter correlation with respective estrogen levels in hippocampal ROI-mask (hair cross on peak voxel; panel A: grey matter changes in left hippocampus) are displayed. Blue voxels, superimposed on respective t-values, correspond to significant FWE-corrected results (p < 0.05). In panel B, bilateral FA and left grey matter values from significant clusters, respectively peak voxel, are extracted and plotted versus estrogen levels (in red) across the menstrual cycles assessed.
Correlations with region of interest extractions
To further assess the relationship between measures of diffusion in the hippocampus and estrogen levels across the menstrual cycle, we extracted mean FA, AD and RD values from significant voxels identified in the previous volumetric FA analysis in the hippocampal ROIs. To visualize scalar measure differences across the menstrual cycles, Fig. 2C includes plots of extracted hippocampal FA and RD values against menstrual cycle days. Estrogen levels were inversely correlated with RD (mean RD, r = −0.460, p = 0.012; Fig. 2C, bottom right) but not with AD (mean AD, r = 0.116, p = 0.547; not shown).
To evaluate a potential time lag in the effects of estrogen on white matter microstructure, we conducted a cross correlation analysis between estrogen levels and FA values to model the relationship of two time series and to display lag-associations at single points in time. This analysis revealed that the largest correlation between FA and estrogen occurred at zero time lag.
Discussion
The current study provides evidence that hippocampal changes parallel fluctuations in endogenous ovarian hormone levels across the menstrual cycle. Specifically, greater FA was significantly associated with increased estrogen levels in bilateral hippocampus. Further, estrogen levels were negatively correlated with RD levels. These diffusion scalar changes in the hippocampus were extended by grey matter signal changes in one hippocampal hemisphere. These multimodal findings suggest significant dynamics in hippocampal structure across the menstrual cycle with a potentially myelin-related process underlying the white matter change.
Our results imply a rapid timeline for menstrual cycle associated structural hippocampal changes in the human brain. The peaks in FA in our data mirror the increases in estrogen, and are most prominent during the estrogen surge shortly before ovulation (Fig. 3, panel C). We found zero time lag in the cross correlation between FA values and estrogen levels, which suggests an immediate effect for high estrogen within a range of 2-3 days. This pattern is consistent with previous grey matter imaging findings that report an acute increase in hippocampal volume from the early follicular phase to the late follicular phase54, as well as the late luteal phase to the late follicular phase in humans28. Evidence from rodent studies also supports a remarkable degree of hippocampal plasticity change in rapid response to estrogen-rises across the estrous cycle6,18.
Several different processes could induce the changes in hippocampal FA that we report here. Since FA quantifies the strength of directionality in local tissue structure34, higher FA values in the hippocampus might point towards a higher directionality in hippocampal white matter tissue. On a cellular level, changes in white matter structure can be related to alterations in fiber organization, myelination, and changes in astrocytes and angiogenesis55. A potential mechanism that has been proposed to underlie white matter changes following learning-experiments is myelination35. In the current study, the increase in FA is primarily driven by a decrease in RD, a scalar measure that has been associated with myelin changes42. In humans, radial diffusivity has been proposed as a surrogate marker for myelin and radial diffusivity measures have successfully been used to characterize the extent of demyelination following traumatic brain injury56. Hence, the increased hippocampal FA and decreased RD associated with high estrogen levels, could reflect microstructural changes (e.g., such as myelination) due to the trophic effects of estrogen on white matter.
We stress that we do not provide any conclusive measurement for the specific biological microstructure that underlies the short-term dynamics of hippocampal changes we observe across the menstrual cycle. However, we have chosen a well-established FA threshold for identifying white matter3,24,27,57 within our hand-drawn hippocampal masks. White matter structure also seems to mediate the relationship between other steroid hormones, such as cortisol, and the hippocampus associated with longitudinal healthy ageing58. This may be a speculative explanation for the stronger association we observe between the steroid hormone estrogen and the diffusion scalar measures more closely associated with white matter structure (FA, RD) compared to both a diffusion scalar measure that has been described as an inverse measure of membrane density, that is both very similar in grey and white matter (MD = mean diffusivity) or the GM findings from our VBM analysis that are limited to one hemisphere (Fig. 3). However, although informative, diffusion-weighted MRI is an indirect measure of structural changes in the brain and the underlying cellular mechanisms remain to be clarified. The hippocampus represents a heterogeneous brain region and both, grey matter volume59 and white matter structure58 have been linked to hippocampal neuroplastic changes in health. The hippocampal changes we find across the menstrual cycle seem to be located at grey-white matter boundaries (Fig. 2) and we cannot exclude that grey matter changes or changes in vasculature contribute to the changes we observe in diffusion scalar measures.
Combining high-resolution human MRI data with histologically validated neuroimaging datasets in rodents can be a useful approach to investigate the changes that occur at the level of the neuronal microstructure. A pioneering study by Sagi and colleagues (2012) compared DWI imaging in humans, with combined DWI imaging/histological analysis in rodents, after each group completed a comparable short-term memory task60. The authors found a significant change in FA, and mean diffusivity, in hippocampal regions 2 hours post-learning in both the human and rodent data. The histological results from the rat study revealed an increase in synaptophysin and brain-derived neurotrophic factor (BDNF), suggesting a structural remodeling process following the learning task. BDNF is involved in long-term potentiation (LTP)60 which is believed to represent a cellular correlate of learning and memory61. Estrogen has been found to increase BDNF expression in the hippocampus62 and an interaction between estrogen and BDNF has been proposed to mediate women’s risk for pathologies characterized by memory-loss63. A recent study reports a positive correlation between BDNF levels in blood serum and hippocampal volume across the menstrual cycle54. Considering these points, one could speculate that estrogen dependent FA changes in the hippocampus across the menstrual cycle could reflect dynamic structural changes comparable to the neuroplasticity changes that have been observed following short-term learning. A multi-modal study-design across rodents and humans could be applied to test this hypothesis in the future.
One limitation of our current study is that we did not include a sequence to specifically quantify potential hormone-related water shifts in the brain that could also account for the FA change. While we acknowledge the possibility that menstrual-cycle related extracellular/intracellular water shifts may contribute to our findings, we consider this unlikely to be the driving mechanism for the following reasons: (1) A substantial change in hydration-status by a strict hydrating/thirsting protocol that has been shown to induce a significant enlargement of ventricular structure and an increase in cerebrospinal fluid (CSF) did not show any significant grey or white matter changes within the hippocampal region64. (2) We did not find any significant results when analyzing CSF changes and estrogen levels across the menstrual cycle in our dataset. (3) The short-time water changes that have been described to be influenced by fluctuating endogenous sex hormones previously65 show a correlation with progesterone, not estrogen. We did not find any significant correlation between progesterone and any of the scalar diffusion measures. In summary, these observations from the literature and our own data support that a shift from extracellular/intracellular water is unlikely to explain the estrogen-associated hippocampal changes we find across the menstrual cycle. We also acknowledge that we did not include any neurocognitive testing in our protocol. Thus, we cannot address whether the potential changes in white matter across the menstrual cycle may relate to behavioral changes. However, grey matter changes across the menstrual cycle were not associated with any significant changes in cognitive abilities in healthy subjects54. Detailed cognitive assessment may be more relevant to clinical populations, such as patients with premenstrual dysphoric disorder. A major aim of our study was to test the feasibility of a longitudinal DWI design across the female menstrual cycle.
In summary, this study is the first to link subtle hormonal fluctuations occurring during the menstrual cycle to changes in FA. Our findings indicate a remarkable degree of hippocampal plasticity in response to estrogen on a very rapid timescale–within days. These data are consistent with evidence from other studies investigating the impact of the menstrual cycle on the human brain28,54,65,66. A noteworthy consistency of neuroplasticity changes during the ovulatory phase, when estrogen levels peak, prevails across study-design, subject- and scan-number. Our study introduces an approach for simultaneously mapping longitudinal characteristics in endogenous hormone and hippocampal dynamics. We acknowledge the limitations from this single subject design. In light of recent attempts to neurally phenotype single humans67,68, our findings highlight menstrual cycle monitoring in parallel with highly sampled individual neuroimaging data to address fundamental questions about the dynamics of plasticity in the adult brain. Similar protocols might be necessary to adequately capture plasticity dynamics when designing future MRI studies that include female participants. The present study represents an important first step towards creating a personalized map of the individual human brain by integrating potential mediators, such as menstrual cycle phase.
Additional Information
How to cite this article: Barth, C. et al. In-vivo Dynamics of the Human Hippocampus across the Menstrual Cycle. Sci. Rep. 6, 32833; doi: 10.1038/srep32833 (2016).
References
Boccardi, M. et al. Effects of hormone therapy on brain morphology of healthy postmenopausal women: a Voxel-based morphometry study. Menopause 13, 584–591, doi: 10.1097/01.gme.0000196811.88505.10 (2006).
Galea, L. A. et al. Endocrine regulation of cognition and neuroplasticity: our pursuit to unveil the complex interaction between hormones, the brain, and behaviour. Can J Exp Psychol 62, 247–260, doi: 10.1037/a0014501 (2008).
Peper, J. S., Hulshoff Pol, H. E., Crone, E. A. & van Honk, J. Sex steroids and brain structure in pubertal boys and girls: a mini-review of neuroimaging studies. Neuroscience 191, 28–37, doi: 10.1016/j.neuroscience.2011.02.014 (2011).
Sacher, J., Neumann, J., Okon-Singer, H., Gotowiec, S. & Villringer, A. Sexual dimorphism in the human brain: evidence from neuroimaging. Magn Reson Imaging 31, 366–375, doi: 10.1016/j.mri.2012.06.007 (2013).
Duarte-Guterman, P., Yagi, S., Chow, C. & Galea, L. A. Hippocampal learning, memory, and neurogenesis: Effects of sex and estrogens across the lifespan in adults. Horm Behav, doi: 10.1016/j.yhbeh.2015.05.024 (2015).
Lee, S. J. & McEwen, B. S. Neurotrophic and neuroprotective actions of estrogens and their therapeutic implications. Annu Rev Pharmacol Toxicol 41, 569–591, doi: 10.1146/annurev.pharmtox.41.1.569 (2001).
Barth, C., Villringer, A. & Sacher, J. Sex hormones affect neurotransmitters and shape the adult female brain during hormonal transition periods. Front Neurosci 9, 37, doi: 10.3389/fnins.2015.00037 (2015).
Shang, Y., Hu, X., DiRenzo, J., Lazar, M. A. & Brown, M. Cofactor dynamics and sufficiency in estrogen receptor-regulated transcription. Cell 103, 843–852 (2000).
Halachmi, S. et al. Estrogen receptor-associated proteins: possible mediators of hormone-induced transcription. Science 264, 1455–1458 (1994).
Kessler, R. C. Epidemiology of women and depression. J Affect Disord 74, 5–13 (2003).
Freeman, E. W., Sammel, M. D., Boorman, D. W. & Zhang, R. Longitudinal pattern of depressive symptoms around natural menopause. JAMA Psychiatry 71, 36–43, doi: 10.1001/jamapsychiatry.2013.2819 (2014).
Freeman, E. W., Sammel, M. D., Lin, H. & Nelson, D. B. Associations of hormones and menopausal status with depressed mood in women with no history of depression. Arch Gen Psychiatry 63, 375–382, doi: 10.1001/archpsyc.63.4.375 (2006).
Munk-Olsen, T., Laursen, T. M., Pedersen, C. B., Mors, O. & Mortensen, P. B. New parents and mental disorders: a population-based register study. JAMA 296, 2582–2589, doi: 10.1001/jama.296.21.2582 (2006).
MacQueen, G. M. et al. Course of illness, hippocampal function, and hippocampal volume in major depression. Proc Natl Acad Sci USA 100, 1387–1392, doi: 10.1073/pnas.0337481100 (2003).
Campbell, S., Marriott, M., Nahmias, C. & MacQueen, G. M. Lower hippocampal volume in patients suffering from depression: a meta-analysis. Am J Psychiatry 161, 598–607, doi: 10.1176/appi.ajp.161.4.598 (2004).
Videbech, P. & Ravnkilde, B. Hippocampal volume and depression: a meta-analysis of MRI studies. Am J Psychiatry 161, 1957–1966, doi: 10.1176/appi.ajp.161.11.1957 (2004).
Mitra, S. W. et al. Immunolocalization of estrogen receptor beta in the mouse brain: comparison with estrogen receptor alpha. Endocrinology 144, 2055–2067, doi: 10.1210/en.2002-221069 (2003).
Barha, C. K. & Galea, L. A. Influence of different estrogens on neuroplasticity and cognition in the hippocampus. Biochim Biophys Acta 1800, 1056–1067, doi: 10.1016/j.bbagen.2010.01.006 (2010).
Neufang, S. et al. Sex differences and the impact of steroid hormones on the developing human brain. Cereb Cortex 19, 464–473, doi: 10.1093/cercor/bhn100 (2009).
Hu, S., Pruessner, J. C., Coupe, P. & Collins, D. L. Volumetric analysis of medial temporal lobe structures in brain development from childhood to adolescence. Neuroimage 74, 276–287, doi: 10.1016/j.neuroimage.2013.02.032 (2013).
Kesler, S. R. et al. Amygdala and hippocampal volumes in Turner syndrome: a high-resolution MRI study of X-monosomy. Neuropsychologia 42, 1971–1978, doi: 10.1016/j.neuropsychologia.2004.04.021 (2004).
Ingalhalikar, M. et al. Sex differences in the structural connectome of the human brain. Proc Natl Acad Sci USA 111, 823–828, doi: 10.1073/pnas.1316909110 (2014).
Goto, M. et al. 3 Tesla MRI detects accelerated hippocampal volume reduction in postmenopausal women. J Magn Reson Imaging 33, 48–53, doi: 10.1002/jmri.22328 (2011).
Kranz, G. S. et al. White matter microstructure in transsexuals and controls investigated by diffusion tensor imaging. J Neurosci 34, 15466–15475, doi: 10.1523/JNEUROSCI.2488-14.2014 (2014).
Pletzer, B. et al. Menstrual cycle and hormonal contraceptive use modulate human brain structure. Brain Res 1348, 55–62, doi: 10.1016/j.brainres.2010.06.019 (2010).
Petersen, N., Kilpatrick, L. A., Goharzad, A. & Cahill, L. Oral contraceptive pill use and menstrual cycle phase are associated with altered resting state functional connectivity. Neuroimage 90, 24–32, doi: 10.1016/j.neuroimage.2013.12.016 (2014).
De Bondt, T. et al. Does the use of hormonal contraceptives cause microstructural changes in cerebral white matter? Preliminary results of a DTI and tractography study. Eur Radiol 23, 57–64, doi: 10.1007/s00330-012-2572-5 (2013).
Protopopescu, X. et al. Hippocampal structural changes across the menstrual cycle. Hippocampus 18, 985–988, doi: 10.1002/hipo.20468 (2008).
Wegienka, G. & Baird, D. D. A comparison of recalled date of last menstrual period with prospectively recorded dates. J Womens Health (Larchmt) 14, 248–252, doi: 10.1089/jwh.2005.14.248 (2005).
Bauman, J. E. Basal body temperature: unreliable method of ovulation detection. Fertil Steril 36, 729–733 (1981).
Coutts, J. R., Adam, A. H. & Fleming, R. The deficient luteal phase may represent an anovulatory cycle. Clin Endocrinol (Oxf) 17, 389–394 (1982).
Pawluski, J. L., Brummelte, S., Barha, C. K., Crozier, T. M. & Galea, L. A. Effects of steroid hormones on neurogenesis in the hippocampus of the adult female rodent during the estrous cycle, pregnancy, lactation and aging. Front Neuroendocrinol 30, 343–357, doi: 10.1016/j.yfrne.2009.03.007 (2009).
Qiu, L. R. et al. Hippocampal volumes differ across the mouse estrous cycle, can change within 24 hours, and associate with cognitive strategies. Neuroimage 83, 593–598, doi: 10.1016/j.neuroimage.2013.06.074 (2013).
Alexander, A. L., Lee, J. E., Lazar, M. & Field, A. S. Diffusion tensor imaging of the brain. Neurotherapeutics 4, 316–329, doi: 10.1016/j.nurt.2007.05.011 (2007).
Taubert, M., Villringer, A. & Ragert, P. Learning-related gray and white matter changes in humans: an update. Neuroscientist 18, 320–325, doi: 10.1177/1073858411419048 (2012).
Raffin, E. & Dyrby, T. B. Diagnostic approach to functional recovery: diffusion-weighted imaging and tractography. Front Neurol Neurosci 32, 26–35, doi: 10.1159/000348818 (2013).
Ling, J. M. et al. Biomarkers of increased diffusion anisotropy in semi-acute mild traumatic brain injury: a longitudinal perspective. Brain 135, 1281–1292, doi: 10.1093/brain/aws073 (2012).
Dinkel, J. et al. Long-term white matter changes after severe traumatic brain injury: a 5-year prospective cohort. AJNR Am J Neuroradiol 35, 23–29, doi: 10.3174/ajnr.A3616 (2014).
Beaulieu, C. The basis of anisotropic water diffusion in the nervous system-a technical review. NMR Biomed 15, 435–455, doi: 10.1002/nbm.782 (2002).
Herting, M. M., Maxwell, E. C., Irvine, C. & Nagel, B. J. The impact of sex, puberty, and hormones on white matter microstructure in adolescents. Cereb Cortex 22, 1979–1992, doi: 10.1093/cercor/bhr246 (2012).
Steele, C. J., Scholz, J., Douaud, G., Johansen-Berg, H. & Penhune, V. B. Structural correlates of skilled performance on a motor sequence task. Front Hum Neurosci 6, 289, doi: 10.3389/fnhum.2012.00289 (2012).
Song, S. K. et al. Dysmyelination revealed through MRI as increased radial (but unchanged axial) diffusion of water. Neuroimage 17, 1429–1436 (2002).
First, M. B. The DSM series and experience with DSM-IV. Psychopathology 35, 67–71, doi: 65121 (2002).
First, M. B. G. M., Spitzer, R. L., Williams, J. B. W. & Benjamin, L. S. Structured Clinical Interview for DSM-IV Axis II Personality Disorders, (SCID-II) American Psychiatric Press, Inc.: Washington, D.C, (1997).
Hamilton, M. A rating scale for depression. J Neurol Neurosurg Psychiatry 23, 56–62 (1960).
Behrens, T. E. et al. Characterization and propagation of uncertainty in diffusion-weighted MR imaging. Magn Reson Med 50, 1077–1088, doi: 10.1002/mrm.10609 (2003).
Smith, S. M. et al. Advances in functional and structural MR image analysis and implementation as FSL. Neuroimage 23 Suppl 1, S208–S219, doi: 10.1016/j.neuroimage.2004.07.051 (2004).
Smith, S. M. et al. Tract-based spatial statistics: voxelwise analysis of multi-subject diffusion data. Neuroimage 31, 1487–1505, doi: 10.1016/j.neuroimage.2006.02.024 (2006).
Douaud, G. et al. Anatomically related grey and white matter abnormalities in adolescent-onset schizophrenia. Brain 130, 2375–2386, doi: 10.1093/brain/awm184 (2007).
Good, C. D. et al. A voxel-based morphometric study of ageing in 465 normal adult human brains. Neuroimage 14, 21–36, doi: 10.1006/nimg.2001.0786 (2001).
Arelin, K. et al. Progesterone mediates brain functional connectivity changes during the menstrual cycle-a pilot resting state MRI study. Front Neurosci 9, 44, doi: 10.3389/fnins.2015.00044 (2015).
Desikan, R. S. et al. An automated labeling system for subdividing the human cerebral cortex on MRI scans into gyral based regions of interest. Neuroimage 31, 968–980, doi: 10.1016/j.neuroimage.2006.01.021 (2006).
Smith, S. M. & Nichols, T. E. Threshold-free cluster enhancement: addressing problems of smoothing, threshold dependence and localisation in cluster inference. Neuroimage 44, 83–98, doi: 10.1016/j.neuroimage.2008.03.061 (2009).
Lisofsky, N. et al. Hippocampal volume and functional connectivity changes during the female menstrual cycle. Neuroimage 118, 154–162, doi: 10.1016/j.neuroimage.2015.06.012 (2015).
Zatorre, R. J., Fields, R. D. & Johansen-Berg, H. Plasticity in gray and white: neuroimaging changes in brain structure during learning. Nat Neurosci 15, 528–536, doi: 10.1038/nn.3045 (2012).
Lin, M., He, H., Schifitto, G. & Zhong, J. Simulation of changes in diffusion related to different pathologies at cellular level after traumatic brain injury. Magn Reson Med. doi: 10.1002/mrm.25816 (2015).
Steele, C. J., Bailey, J. A., Zatorre, R. J. & Penhune, V. B. Early musical training and white-matter plasticity in the corpus callosum: evidence for a sensitive period. J Neurosci 33, 1282–1290, doi: 10.1523/JNEUROSCI.3578-12.2013 (2013).
Cox, S. R. et al. Does white matter structure or hippocampal volume mediate associations between cortisol and cognitive ageing? Psychoneuroendocrinology 62, 129–137, doi: 10.1016/j.psyneuen.2015.08.005 (2015).
Killgore, W. D., Olson, E. A. & Weber, M. Physical exercise habits correlate with gray matter volume of the hippocampus in healthy adult humans. Sci Rep 3, 3457, doi: 10.1038/srep03457 (2013).
Sagi, Y. et al. Learning in the fast lane: new insights into neuroplasticity. Neuron 73, 1195–1203, doi: 10.1016/j.neuron.2012.01.025 (2012).
Foy, M. R. et al. 17beta-estradiol enhances NMDA receptor-mediated EPSPs and long-term potentiation. J Neurophysiol 81, 925–929 (1999).
Zhou, J., Zhang, H., Cohen, R. S. & Pandey, S. C. Effects of estrogen treatment on expression of brain-derived neurotrophic factor and cAMP response element-binding protein expression and phosphorylation in rat amygdaloid and hippocampal structures. Neuroendocrinology 81, 294–310, doi: 10.1159/000088448 (2005).
Sohrabji, F. & Lewis, D. K. Estrogen-BDNF interactions: implications for neurodegenerative diseases. Front Neuroendocrinol 27, 404–414, doi: 10.1016/j.yfrne.2006.09.003 (2006).
Streitburger, D. P. et al. Investigating structural brain changes of dehydration using voxel-based morphometry. PLoS One 7, e44195, doi: 10.1371/journal.pone.0044195 (2012).
Hagemann, G. et al. Changes in brain size during the menstrual cycle. PLoS One 6, e14655, doi: 10.1371/journal.pone.0014655 (2011).
Franke, K., Hagemann, G., Schleussner, E. & Gaser, C. Changes of individual BrainAGE during the course of the menstrual cycle. Neuroimage 115, 1–6, doi: 10.1016/j.neuroimage.2015.04.036 (2015).
Poldrack, R. A. et al. Long-term neural and physiological phenotyping of a single human. Nat Commun 6, 8885, doi: 10.1038/ncomms9885 (2015).
Laumann, T. O. et al. Functional System and Areal Organization of a Highly Sampled Individual Human Brain. Neuron 87, 657–670, doi: 10.1016/j.neuron.2015.06.037 (2015).
Acknowledgements
The study was supported by a research grant from the Society in Science–The Branco Weiss Fellowship to Dr. Julia Sacher and research funds from the Max Planck Society. We thank Dr. Pierre-Louis Bazin and Dr. Daniel Margulies for critical discussion.
Author information
Authors and Affiliations
Contributions
C.B., K.A., K.M., A.V. and J.S. conception and design of the study; J.S. and A.V. provided funding; A.P., I.B., J.S. and J.K. conducted the experiments; A.P. and J.K. technical support; C.B., C.J.S., I.B. and J.S. data-analysis; C.B., C.J.S., K.M., V.P.R., A.V. and J.S. data interpretation; C.B. and J.S. drafted manuscript; C.B., C.J.S., K.M., V.P.R., A.V. and J.S. critical revision; all authors approved the final draft of the manuscript for submission.
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Electronic supplementary material
Rights and permissions
This work is licensed under a Creative Commons Attribution 4.0 International License. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in the credit line; if the material is not included under the Creative Commons license, users will need to obtain permission from the license holder to reproduce the material. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/
About this article

Cite this article
Barth, C., Steele, C., Mueller, K. et al. In-vivo Dynamics of the Human Hippocampus across the Menstrual Cycle. Sci Rep 6, 32833 (2016). https://doi.org/10.1038/srep32833
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/srep32833
This article is cited by
-
Brain flexibility increases during the peri-ovulatory phase as compared to early follicular phase of the menstrual cycle
Scientific Reports (2024)
-
Sex and menstrual cycle influence human spatial navigation strategies and performance
Scientific Reports (2023)
-
Learning exceptions to category rules varies across the menstrual cycle
Scientific Reports (2023)
-
Ultra-high-field 7T MRI reveals changes in human medial temporal lobe volume in female adults during menstrual cycle
Nature Mental Health (2023)
-
Profiling GABA(A) Receptor Subunit Expression in the Hippocampus of PMDD Rat Models Based on TCM Theories
Molecular Neurobiology (2023)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
