
Abstract
The anthracnose caused by Colletotrichum species is an important disease that primarily causes fruit rot in pepper. Eighty-eight strains representing seven species of Colletotrichum were obtained from rotten pepper fruits in Sichuan Province, China, and characterized according to morphology and the glyceraldehyde-3-phosphate dehydrogenase (GAPDH) sequence. Fifty-two strains were chosen for identification by phylogenetic analyses of multi-locus sequences, including the nuclear ribosomal internal transcribed spacer (ITS) region and the β-tubulin (TUB2), actin (ACT), calmodulin (CAL) and GAPDH genes. Based on the combined datasets, the 88 strains were identified as Colletotrichum gloeosporioides, C. siamense, C. fructicola, C. truncatum, C. scovillei, and C. brevisporum, and one new species was detected, described as Colletotrichum sichuanensis. Notably, C. siamense and C. scovillei were recorded for the first time as the causes of anthracnose in peppers in China. In addition, with the exception of C. truncatum, this is the first report of all of the other Colletotrichum species studied in pepper from Sichuan. The fungal species were all non-host-specific, as the isolates were able to infect not only Capsicum spp. but also Pyrus pyrifolia in pathogenicity tests. These findings suggest that the fungal species associated with anthracnose in pepper may inoculate other hosts as initial inoculum.
Similar content being viewed by others


Introduction
Pepper (Capsicum annuum), an important fruit that is also used as a spice, is rich in vitamins, capsaicin and capsochrome. One of the primary pepper-growing provinces in China is Sichuan Province, where the crop is cultivated over an area of approximately 70 thousand hm2, with approximately 1,000 thousand tons of annual output.
Colletotrichum is an important pathogenic genus worldwide. These fungi cause disease symptoms that are generally known as anthracnose in a wide range of vegetables, fruits and other crops1. In pepper, anthracnose is a destructive disease caused by a complex of Colletotrichum species that causes extensive yield losses at both the pre- and post-harvest stages during warm and rainy seasons2.
Anthracnose in pepper is associated with at least eleven Colletotrichum species, including C. truncatum3,4,5,6, C. gloeosporioides6,7,8,9, C. acutatum6,10,11, C. coccodes12,13,14,15, C. fructicola7,16,17, C. siamense17,18, C. dematium14, C. boninense19,20, C. brevisporum, C. cliviae7, and C. scovillei21. Eight of these species have been reported in China, whereas C. siamense, C. dematium and C. scovillei have not. To date, only three species (C. acutatum22, C. truncatum5 and C. boninense19) have been reported in Sichuan Province, although previous studies have not fully investigated the Colletotrichum species associated with pepper anthracnose in this province.
Colletotrichum gloeosporioides is a species complex that was formerly regarded as a cosmopolitan species that infects various hosts, including pepper; however, it might have been misidentified as the causative agent. For example, Phoulivong et al.16 failed to isolate C. gloeosporioides sensu stricto from tropical fruits, although C. gloeosporioides was previously thought to be the cause of tropical fruit rot. Similarly, Lima et al.23 found that none of the strains isolated from mango (a tropical fruit) belonged to C. gloeosporioides sensu stricto; instead, phylogenetic analysis revealed that most of the strains belonged to the ‘gloeosporioides’ complex. Although the nuclear ribosomal internal transcribed spacer (ITS) region is the most commonly used region for differentiating fungi24, it has also been widely acknowledged that this region cannot fully differentiate among Colletotrichum species18,25,26,27. Multi-locus phylogeny is broadly applied for identifying Colletotrichum spp. Weir et al.18 have suggested that the C. gloeosporioides complex consists of 23 taxa, according to multi-locus phylogeny. Several new species have also been described on the basis of multi-locus phylogeny, e.g., C. anthrisci, C. liriopes, C. rusci and C. verruculosum3; C. bletillum, C. caudasporum, C. duyunensis, C. endophytum, C. excelsum-altitudum, C. guizhouensis and C. ochracea24; C. asianum, C. fructicola and C. siamense26; C. cliviae, C. hippeastri and C. hymenocallidis27; C. corchorum-capsularis28; and C. endophytica29.
Despite several reports of Colletotrichum species in pepper from limited collection areas5,19,22, little is known about the association of these species with pepper in Sichuan Province, China. Further, it remains unclear whether all of the species isolated from pepper are equally pathogenic and host specific.
The objective of this study was to characterize the Colletotrichum species associated with anthracnose in pepper from different geographic areas of Sichuan Province, China, according to morphological, multi-locus phylogenetic and pathogenic characteristics.
Results
Symptom types of pepper anthracnose caused by Colletotrichum species
A total of 173 symptomatic samples were collected from primary pepper-producing regions, covering 31 districts in Sichuan Province, China. Based on the morphological characteristics coupled with the microscopic observations, the following three typical symptom types in the infected pepper fruits in the fields were noted (Fig. 1): Type I: the typical symptoms were variation in colour from dark brown to black, sunken lesions and many black acervuli on the surface, which usually produced dirty white conidial masses under humid conditions. In some cultivars with less pulp, these typical conidial masses were infrequently observed in black acervuli (Fig. 1a–c). The conidia responding for this symptom type had the typical falcate spores; Type II: the symptoms included sunken necrotic tissues, ranging in colour from brown to black, with concentric rings of acervuli (Fig. 1d–f). The main Type II symptoms were similar to the main Type I symptoms, except that the acervuli produced viscous, flesh-pink conidial masses under wet conditions and cylindrical to long cylindrical conidia; and Type III: the typical symptoms included light brown to dark brown tissues, and sunken, orange conidial masses that were powdery in a dry environment; in addition, the conidia were fusiform (Fig. 1g–i). Notably, more than one type of disease symptom was often observed in a single pepper fruit in the field.

Typical symptoms of pepper anthracnose in the field.
(a–c) Type I symptoms were characterized by dark brown to black, sunken lesions with a slightly raised rim and many black acervuli on the surface, which produced dirty white conidial masses under humid conditions. (d–f) Type II symptoms included dark brown to black, sunken lesions with many black acervuli on the surface, which produced flesh pink, viscous conidial masses under humid conditions. (g–i) Type III symptoms included brown to light black to dark brown, sunken, lesions with orange conidial masses.
Colletotrichum species collection
A total of 352 single-spore cultures were isolated from 173 symptomatic samples. Eighty-eight isolates were subsequently selected for further determination on the basis of their origins, colony characteristics and conidial morphologies (Table 1).
Morphological and cultural characteristics
Eighty-eight isolates were classified into six morphological groups according to morphological and cultural characteristics. Group 1 included 23 isolates fitting the description of the C. gloeosporioides complex, and Group 2 included 16 isolates fitting the description of C. fructicola. In addition, Group 3 consisted of 32 isolates matching the description of C. truncatum, Group 4 had seven isolates fitting the description of the C. acutatum complex, and Group 5 consisted of six isolates matching the description of C. brevisporum. Group 6 contained four isolates that did not fit the description of any currently known Colletotrichum species. Group 3 (C. truncatum) was the predominant group, accounting for 36.4% of the total isolates. A summary of the morphological data for the Colletotrichum species in Groups 1–6 is presented in Table 2.
Colony characteristics (Fig. 2): Distinct morphology on potato dextrose agar (PDA) was observed in each group after 7 days. The isolates from Group 1 produced pale yellowish colonies, with sparse white aerial mycelia. The reverse side of the colonies was white, and many bright orange conidial masses were observed near the inoculum point. The colonies produced by Group 2 isolates varied from white to black-green on PDA, with dense grey aerial mycelia and a few bright orange conidial masses near the inoculum point. The colonies produced by Group 3 isolates varied from pale grey to dark grey, with dense pale grey aerial mycelia and small black granules over the entire surface. The reverse side of the colonies was dark brown, and a few pale yellow conidial masses were observed near the inoculum point. The colonies produced by Group 4 isolates varied from white to pale orange, with dense white aerial mycelia, and the reverse side of the colonies was pale orange. Isolates belonging to Group 5 produced dark grey colonies with sparse grey aerial mycelia. The reverse side of the colonies was grey, and a few bright orange conidial masses were observed near the inoculum point, as well as some spots scattered over the colony surface. Lastly, the isolates from Group 6 produced pale grey colonies, with sparse white aerial mycelia. The colonies from Group 3, 4 and 5 were stable and unique, and the colonies from Group 2 were significantly different compared with those from the other groups under stable culture conditions.

Morphology and cultural characteristics of Colletotrichum spp. from pepper anthracnose.
(A) Upper view of a colony on PDA; (B) reverse view of colony on PDA; (C) micrographs of conidia of Colletotrichum spp.; (D) micrographs of conidial appressoria of Colletotrichum spp.; (E) micrographs of mycelial appressoria of Colletotrichum spp.; (F) conidiophores. Scale bars = 10 μm for (C–E); 20 μm for (F).
Growth rate (Table 2): Group 4 exhibited a significantly different growth rate compared with the other five groups (P = 0.05). The isolates from Group 6 (6.1 ± 0.4 mm/day) grew the fastest, followed by those from Group 1 (5.6 ± 1.2 mm/day), Group 2 (5.9 ± 0.4 mm/day), Group 5 (5.3 ± 0.6 mm/day), Group 3 (4.5 ± 0.5 mm/day) and Group 4 (3.8 ± 0.4 mm/day).
Conidial morphology (Table 2 and Fig. 2): The following four types of conidia were observed: cylindrical (observed in Groups 1, 2 and 6), falcate (Group 3), fusiform (Group 4) and long cylindrical (Group 5). The conidial widths of Group 6 were significantly different from those of Groups 1 and 2; however, all of these groups had cylindrical conidia with obtuse to slightly rounded ends. The conidia produced by the Group 3 isolates were falcate, with gradual tapering towards each end. Group 4 produced fusiform conidia, whereas Group 5 produced long and cylindrical conidia, with obtuse to slightly rounded ends. The differences in the conidial shapes of Groups 3, 4 and 5 were very significant, allowing these groups to be easily distinguished from one another. Almost all of the conidia were aseptate, but they often developed a septum after germinating and forming appressoria.
Conidial appressorium morphology (Table 2 and Fig. 2): There was little distinction among the groups in terms of the sizes and shapes of conidial appressoria, except for Groups 4 and 6, which exhibited significant differences compared with the other groups. The conidial appressoria of Groups 1, 2, 3 and 5 varied from ovoid to slightly irregular in shape and from brown to dark black in colour. Group 4 produced grey, globular and smaller conidial appressoria. Most of the conidial appressoria produced by Group 6 were irregular and pale brown to dark brown, with a crenate edge.
Mycelial appressorium morphology (Table 2 and Fig. 2): The mycelial appressoria produced by the isolates of Groups 1 and 2 varied from ovoid, clavate and slightly irregular to irregular, smooth or slightly lobed, and they were light brown to brown in colour. The appressoria of Group 3 ranged from ovate, ellipsoidal or slightly irregular to irregular in shape, and they were smooth or lobate and brown to dark brown. The appressoria produced by Group 4 were globose or ovate to slightly irregular, and they were light brown to brown and smaller in size than those of the other groups. In addition, the appressoria produced by Group 5 varied from ovoid, clavate or slightly irregular to irregular in shape. They were smooth or slightly lobed and brown to dark brown and were sometimes black in the middle. Further, the appressoria of Group 6 were ellipsoidal or irregular, smooth or slightly lobed to strongly lobed, solitary or in chains, and light brown to brown in colour.
Conidiophores (Fig. 2): The conidiophores of all groups were hyaline to pale brown, simple or septate, rarely branched, and smooth walled. Four types of conidiophores were observed: (i) nearly cylindrical, but narrower towards the end (as observed in Groups 1 and 2); (ii) cylindrical, with a truncate top (Groups 3 and 5); (iii) shortly clavate, nearly hyaline, with a cylindrical base, and obviously inflated, with gradually tapering towards the top (Group 4); and (iv) frequently produced by mycelia, cylindrical, with swollen ends (oblong) and slight narrowing in some areas (Group 6).
Setae: All isolates from Groups 3 and 5 and some isolates from Group 1 produced setae; in contrast, the isolates from all the other groups rarely produced setae. The setae were commonly smooth, septate, and light brown to dark brown in colour, base cylindrical to conical, and sometimes slightly inflated, and the tips were acute to roundish. No obvious differences in setal characteristics (shape and dimensions) were found among the different groups when grown on PDA.
Sclerotia and Ascomata: Most Group 5 isolates steadily produced a large amount of black solids that appeared similar to sclerotia and were round to irregular and semi-immersed. Conidial masses and setae sometimes formed on the black solids. On PDA, Group 6 isolates always produced ascomata in clusters, which were brown and globose to near globose and possessed a neck. The isolates from the other groups rarely produced ascomata, even in host tissues.
Phylogenetic analysis
A phylogram generated based on the GAPDH gene region revealed 5 primary clades (i.e., C. truncatum, C. brevisporum, C. gloeosporioides sensu lato, C. acutatum sensu lato and one unknown species (Colletotrichum sp.)) (Fig. 3).

A neighbour-joining tree based on partial GAPDH gene sequences from 88 Colletotrichum isolates.
Parsimony bootstrap values of more than 50% are shown at the nodes. Isolates selected for subsequent phylogenetic analyses are highlighted in red.
Fifty-two representative isolates were chosen from the morphological groups for molecular analysis, including 25 isolates from the C. gloeosporioides complex, 10 C. truncatum isolates, seven C. acutatum sensu lato isolates, six C. brevisporum isolates and four Colletotrichum sp. isolates.
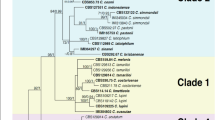
Multi-locus phylogenetic analysis was conducted among 87 strains, with Monilochaetes infuscans (CBS 869.96) used as an outgroup (Table 3). The dataset for five genes (ITS, TUB2, ACT, GAPDH and CAL) contained 2,155 characters, including alignment gaps, of which 997 characters were parsimony-informative, 321 were parsimony-uninformative, and 837 were constant. This parsimony analysis resulted in the most parsimonious tree (TL = 2800, CI = 0.7257, RI = 0.9541, RC = 0.6924, and HI = 0.2743). The phylogram showed that the 52 pepper anthracnose isolates belonged to seven distinct clades. The isolates from Group 2 clustered with C. fructicola, those from Group 3 clustered with C. truncatum, those from Group 4 clustered with C. scovillei, and those from Group 5 clustered with C. brevisporum. The Group 1 isolates grouped with two clades; 4 isolates clustered with C. gloeosporioides, and the remaining isolates clustered with C. siamense (Fig. 4). After combining two phylograms (Figs 3 and 4), 8 and 16 strains were found to belong to C. gloeosporioides sensu stricto and C. siamense, respectively. The isolates from Group 6 were from an unknown species (Colletotrichum sp.). The submission number of the sequence alignment in TreeBASE is 18832.

Phylogram generated from maximum parsimony analysis based on alignment of ITS, TUB2, ACT, GADPH and CAL gene sequences, showing the phylogenetic relationships of Colletotrichum species causing anthracnose disease in Capsicum annuum from Sichuan Province, China.
Parsimony bootstrap values of more than 50% are shown at the nodes. Isolates from this study are shown in bold. The tree is rooted with Monilochaetes infuscans.
Taxonomy
Colletotrichum sichuanensis G.S. Gong & F.L. Liu, sp. nov. (Fig. 5).

Colletotrichum sichuanensis (from holotype).
(a,b) Colonies on PDA at 7 days, upper (a) and reverse (b); (c,d) conidia; (e) conidiogenous cells; (f,g) conidial appressoria; (h–j) mycelial appressoria; (k) ascomata on PDA; (l) peridium; (m–o) asci; (p,q) ascospores. Scale bars: c, d, f–j, p, q = 10 μm; e, l, m, o = 20 μm; n = 40 μm
MycoBank: MB 815288.
Etymology: sichuanensis, in reference to the province where the type was found.
Description: Colonies on PDA at first white, later becoming pale grey and reverse pale grey, with a maximum diameter of 68.7 mm over 5 days at 28 °C and a growth rate of 6.1–6.4 mm/day ( = 6.3 ± 0.1, n = 5). Aerial mycelium white and sparse, with the frequent absence of conidial masses. Ascomata nearly always present in clusters on PDA. Conidiophores generated from mycelia are nearly hyaline, branched, and cylindrical, with slightly swollen ends, simple or occasionally branched. Conidia common on mycelia, one-celled, smooth-walled, hyaline, and cylindrical, with obtuse to slightly rounded ends, 15.0–18.9 × 5.4–6.5 μm (
= 6.3 ± 0.1, n = 5). Aerial mycelium white and sparse, with the frequent absence of conidial masses. Ascomata nearly always present in clusters on PDA. Conidiophores generated from mycelia are nearly hyaline, branched, and cylindrical, with slightly swollen ends, simple or occasionally branched. Conidia common on mycelia, one-celled, smooth-walled, hyaline, and cylindrical, with obtuse to slightly rounded ends, 15.0–18.9 × 5.4–6.5 μm ( = 16.9 ± 1.0 × 6.2 ± 0.3, n = 30). Conidial appressoria light brown to dark brown, slightly irregular to irregular, crenate or lobed, 8.1–12.4 × 5.4–8.8 μm (
= 16.9 ± 1.0 × 6.2 ± 0.3, n = 30). Conidial appressoria light brown to dark brown, slightly irregular to irregular, crenate or lobed, 8.1–12.4 × 5.4–8.8 μm ( = 10.2 ± 1.0 × 6.8 ± 0.8, n = 30). Appressoria in slide culture light brown to brown, ellipsoidal or irregular, smooth or slightly lobed to strongly lobed, solitary or in chains, 6.4–20.2 × 4.8–9.8 μm (
= 10.2 ± 1.0 × 6.8 ± 0.8, n = 30). Appressoria in slide culture light brown to brown, ellipsoidal or irregular, smooth or slightly lobed to strongly lobed, solitary or in chains, 6.4–20.2 × 4.8–9.8 μm ( = 11.5 ± 3.0 × 7.0 ± 1.1, n = 30). Setae absent.
= 11.5 ± 3.0 × 7.0 ± 1.1, n = 30). Setae absent.
Teleomorph: Glomerella sp.
Ascomata, light brown to brown, globose to subglobose, with a neck, arranged in clusters. Peridium of textura angularis, thick-walled. Asci 30.8–61.6 × 7.4–13.8 μm ( = 47.5 ± 8.0 × 9.5 ± 1.5, n = 30), unitunicate, thin-walled, and clavate. Ascospores 10.2–23.3 × 3.9–6.8 μm (
= 47.5 ± 8.0 × 9.5 ± 1.5, n = 30), unitunicate, thin-walled, and clavate. Ascospores 10.2–23.3 × 3.9–6.8 μm ( = 17.5 ± 2.6 × 5.4 ± 0.8, n = 30), one-celled, hyaline, and slightly curved to curved, with obtuse to slightly rounded ends.
= 17.5 ± 2.6 × 5.4 ± 0.8, n = 30), one-celled, hyaline, and slightly curved to curved, with obtuse to slightly rounded ends.
Holotype: Baoxing, Yaan City, Sichuan Province, China, on fruit of Capsicum annuum, 5 September 2013, coll. G. S. Gong (holotype living culture LJTJ30). A living culture (strain LJTJ30) was deposited at the Department of Plant Pathology of Sichuan Agricultural University. Known distribution: Sichuan Province, China.
Additional examined specimens: Jiangyou, Mianyang City, Sichuan Province, China, on Capsicum annuum fruit, 26 July 2013, coll. G. S. Gong (holotype living culture LJTJ3); Yuechi, Guangan City, Sichuan Province, China, on Capsicum annuum fruit, 27 August 2013, coll. F. L. Liu (holotype living culture LJTJ16); and Wenjiang, Chengdu City, Sichuan Province, China, on Capsicum annuum fruit, 3 July 2013, coll. F. L. Liu (holotype living culture LJTJ22). A living culture (strain LJTJ3, LJTJ16 and LJTJ22) was deposited at the Department of Plant Pathology at Sichuan Agricultural University.
Pathogenicity tests
Fifty-two representative isolates selected from among the species were used for pathogenicity testing. All of these isolates were pathogenic to both pepper fruits and pears, although the pathogenicity of each species differed across experimental varieties, with different infection incidences. All species were able to infect Capsicum annuum L. var. conoides (Mill.) Irish and Pyrus pyrifolia at a high incidence. However, C. brevisporum and C. sichuanensis appeared to be only slightly virulent towards Ca. annuum var. dactylus M, with a rather low infection incidence (Table 4 and Fig. 6). These results indicated that some pepper varieties might be resistant to some Colletotrichum species.

Symptoms in pepper and pear after inoculation with Colletotrichum spp. I, Symptoms in pepper (Capsicum annuum var. dactylus M); II, symptoms in pepper (Capsicum annuum L. var. conoides (Mill.) Irish); III, symptoms in pear (Pyrus pyrifolia); (A), symptoms in pepper and pear inoculated with a mycelial disc of C. gloeosporioides; (B), symptoms in pepper and pear inoculated with a mycelial disc of C. siamense; (C), symptoms in pepper and pear inoculated with a mycelial disc of C. fructicola; (D), symptoms in pepper and pear inoculated with a mycelial disc of C. truncatum; (E), symptoms in pepper and pear inoculated with a mycelial disc of C. scovillei; (F), symptoms in pepper and pear inoculated with a mycelial disc of C. brevisporum; (G), symptoms in pepper and pear inoculated with a mycelial disc of C. sichuanensis; (H), the control, inoculated with an agar disc.
Based on the description of the symptoms in pepper after inoculation, C. truncatum was determined to be the pathogen causing Type I symptom, characterized by copious black acervuli with seta and dirty white conidial masses produced on decaying tissues under humid conditions (Fig. 1a–c). C. scovillei induced Type III symptoms (Fig. 1g–i), and the other species caused Type II symptoms (Fig. 1d–f). Our results indicate that with the exception of C. truncatum and C. scovillei, it is difficult to differentiate among Colletotrichum species based solely on the symptom types in the field.
Discussion
The primary objective of this study was to identify the Colletotrichum species that are currently causing anthracnose disease in pepper grown in Sichuan Province, China. Based on the morphological characteristics and phylogenetic analysis, 88 isolates were identified as C. gloeosporioides sensu stricto (eight strains, 9.1%), C. siamense (16 strains, 18.2%), C. fructicola (15 strains, 17.0%), C. truncatum (32 strains, 36.4%), C. scovillei (seven strains, 8.0%), C. brevisporum (six strains, 6.8%) and C. sichuanensis (a new species, four strains, 4.5%). Additionally, C. gloeosporioides and C. siamense could only be distinguished by phylogenetic analyses and not by morphological analyses. The morphological groupings based on colony characteristics, growth rate, conidial morphology, conidial appressorium morphology and mycelial appressorium morphology were almost completely consistent with the results of phylogenetic analysis derived from the molecular data.
In vitro culture-related characteristics were important for differentiating among Colletotrichum species26. C. truncatum, C. scovillei, C. brevisporum, C. sichuanensis isolates and some C. fructicola isolates with unique and relatively stable colonies could be easily distinguished. However, the colonies of C. gloeosporioides, C. siamense and some C. fructicola isolates overlapped in terms of their morphological characteristics, and phenotypic variations were identified among the species under different environmental conditions. The colony growth rate of C. scovillei was significantly slower than those of the species in the other groups. Previous studies have shown that C. acutatum can be differentiated from C. gloeosporioides based on its slower growth rate30. Than et al.2 have also suggested that colony growth rates are important for distinguishing among C. gloeosporioides, C. truncatum and C. acutatum. In the present study, the slow growth of C. scovillei conformed to the characteristics of the C. acutatum complex. The observed differences in conidial size were significant, with the exception of the lengths and widths of Groups 1 and 2. Denoyes and Baudry31 used conidial shape to differentiate among Colletotrichum species that are pathogenic to strawberries, although Cai et al.25 and Crouch et al.32 have suggested that conidial appressoria are taxonomically uninformative and of little use for species identification. In contrast, the conidial appressoria of C. scovillei could be easily distinguished from those of the other species examined in our study, in agreement with the results of Du et al.33. Similarly, Crouch et al.32 have found that the shapes and sizes of mycelial appressoria in combination with the host range are useful for identifying grass-associated Colletotrichum species. We found that the mycelial appressoria produced by C. scovillei and C. brevisporum were typically smoother than those produced by the other species and that all C. truncatum and C. brevisporum isolates steadily produced setae. In addition, C. gloeosporioides has been reported to produce setae occasionally or under certain conditions34, and many other Colletotrichum species are known to produce setae3. In the present study, the cultural characteristics, colony growth rate, conidial shapes and sizes, and conidial and mycelial appressoria were the primary features used for classification.
Morphological examination was conducted to classify the 88 isolates into six groups, although our multi-locus phylogenetic analysis actually identified seven Colletotrichum species. Groups 2–6 contained different Colletotrichum species, and Group 1 consisted of two species: C. gloeosporioides and C. siamense. Thus, morphological criteria alone are not always sufficient for species identification14. Indeed, multi-locus phylogeny showed that the isolates with similar morphological characteristics belonged to the C. gloeosporioides, C. siamense and C. fructicola clades. Moreover, the C. gloeosporioides and C. siamense isolates could not be distinguished according to their morphological and cultural characteristics, indicating that multi-locus phylogenetic analysis is useful for differentiating among species in the Colletotrichum genus. Many investigators have suggested the use of multi-locus phylogenetic analysis to overcome the inadequacies of morphological criteria3,17,24,26,27,35,36,37,38,39.
Colletotrichum gloeosporioides was first described in citrus from Italy40. The name C. gloeosporioides represents both C. gloeosporioides sensu lato, which encompasses the entire species complex, and C. gloeosporioides sensu stricto18. C. gloeosporioides sensu lato consists of at least 22 species, including C. gloeosporioides, C. siamense, and C. fructicola1,18,25,26,41. C. siamense and C. fructicola were originally known as opportunistic pathogens of Coffea arabica berries in Thailand26, and both of these species are non-host-specific. C. fructicola has also been reported to be a pathogen causing pepper anthracnose in Thailand16, India17 and China7. Although Than et al.2 first isolated C. siamense from chilli pepper in Thailand, the isolates belonging to C. siamense were identified as C. gloeosporioides in that study, and Weir et al.18 later revised the classification. C. siamense has also been isolated from pepper in India. However, this species has not been reported to be a causative agent of pepper anthracnose in China. Therefore, this work is the first report of pepper anthracnose caused by C. siamense.
Colletotrichum truncatum, originally described on Phaseolus lunatus, was typified by Damm et al.3, and this species has been associated with anthracnose on legume crops and pepper, as well as on many other hosts3,9,34. The C. capsici isolate typified by Shenoy et al.42 causes anthracnose in a wide range of hosts, including pepper and legume species1,43,44, and Damm et al.3 synonymized the C. capsici taxon with C. truncatum on the basis of its multi-locus phylogeny and morphology. Regardless, not all researchers are in agreement with this viewpoint1.
Colletotrichum acutatum is widely known as a fruit rot pathogen in strawberry2, apple45, pepper2,11 and grape46, and this fungus was first recorded in Australia on Carica papaya, Capsicum frutescens and Delphinium ajacis by Simmonds30. C. acutatum is also a species complex containing at least 14 species, including C. scovillei47. The ex-type strain of C. scovillei was initially identified as C. acutatum48, and Than et al.2 also identified C. scovillei as C. acutatum on chilli pepper from Thailand. Although C. scovillei was identified as C. acutatum in these two papers, it was later revised by Damm et al.47. Kanto et al.21 also isolated C. scovillei from sweet pepper in Japan. In our study, we only isolated C. scovillei belonging to C. acutatum sensu lato from the pepper fruits. Thus, the main species from the C. acutatum complex that is pathogenic to pepper in Sichuan Province might be C. scovillei rather than C. acutatum sensu stricto. To our knowledge, this work is also the first report of C. scovillei as a causative agent of pepper anthracnose in China.
Colletotrichum brevisporum has been recorded on Neoregelia sp. from Thailand, as well as on papaya fruits and Pandanus pygmaeus Thouars35,49. Yang7 have also reported C. brevisporum on pepper from China. The conidial lengths of C. brevisporum in the present study were longer than those reported by Noireung et al.35, but they were consistent with those reported by Yang7.
The results of our phylogenetic analysis strongly support the Colletotrichum sichuanensis clade, which is closely related to C. cliviae. These two species have similar conidial shapes but different conidial sizes; C. sichuanensis has shorter conidia than C. cliviae (21.8 μm), with a mean length of 16.7 μm. C. sichuanensis also differs from C. cliviae with regard to colony colour. In addition, C. sichuanensis steadily produced ascomata on PDA, whereas the other species rarely produced ascomata. Further, C. sichuanensis grew more slowly in culture than C. cliviae (11.3–12.9 mm/day for C. sichuanensis compared with 15.2–16 mm/day for C. cliviae).
Given that they could infect not only Capsicum spp. but also Pyrus pyrifolia, all of the species isolated from pepper in our study were non-host-specific. In addition, C. scovillei was the most virulent species towards Capsicum spp. Tang6 found that C. acutatum and C. truncatum were more virulent than C. gloeosporioides and that the C. acutatum incubation period was the shortest. Further, Than et al.2,14 reported that C. acutatum was a very virulent species that could infect wound-resistant C. chinense PBC 932, whereas C. gloeosporioides and C. capsici (syn. C. truncatum) could not.
Colletotrichum acutatum10, C. truncatum5 and C. boninense19 have been previously reported in Sichuan; however, C. boninense was not isolated in our study; it is possible that this species was missed during sampling or isolation. In summary, C. siamense and C. scovillei are recorded for the first time as causing anthracnose in pepper from China. Additionally, we have identified one new species, which has been introduced as C. sichuanensis.
Methods
Collection and isolation
In 2012 and 2013, pepper fruits with anthracnose symptoms were collected from primary production areas in Sichuan Province, China. Tissues of approximately 5 mm in diameter were collected from the edges of lesions, surface-sterilized with 75% ethanol for 30 s and 1% NaClO for approximately 1 min, washed three times with sterile distilled water, and then dried on sterile filter paper. The treated tissues were plated on PDA supplemented with 50 mg l−1 streptomycin. The plates were incubated at 27 ± 1 °C for 5 days. Single-spore cultures were obtained for each Colletotrichum isolate according to the procedure described by Gong et al.50. The resulting strains were maintained on PDA slants at 4 °C for short-term storage and in 25% glycerol at −70 °C for long-term storage.
Morphological and cultural characterization
Mycelial discs (5 mm diameter) were collected from actively growing areas near the growing edges of 5-day-old cultures, transferred to PDA and incubated at 27 °C in the dark for 10 days. Five replicates were employed. The colony diameter was recorded each day from two perpendicular cross-sections, and the colony characteristics were also recorded.
The sizes and shapes of conidia, asci and ascospores from each culture were recorded. The lengths and widths of 30 conidia, asci and ascospores were measured for each isolate.
Conidial appressoria were induced according to the method of Yang et al.27.
Mycelial appressoria were produced using an improved slide culture technique, as described by Sutton51 and Cai et al.25. One hundred microlitres of hot water agar (WA) was placed on a sterile slide. Mycelial plugs of approximately 2 mm in diameter were inoculated onto one-third of the WA and then incubated in a Petri dish with wet filter paper at 27 °C. After 5–7 days, agar pieces containing the inoculated plugs were gently removed with a scalpel, and the shapes and sizes of the appressoria that formed along the WA were then recorded.
Samples for microscopy were prepared using clear water or lactic acid and observed with a Carl Zeiss Axio Imager Z2 microscope(Germany) or a Nikon Eclipse 80i microscope(Japan) using differential interference contrast (DIC) illumination.
DNA extraction
Fifty-two representative isolates were chosen according to the morphological and cultural characteristics and incubated on PDA at 27 °C for 7–10 days. Mycelia were scraped from the colony surface using a sterile medicine spoon. Total genomic DNA was extracted from the isolates using a modified protocol, as outlined by Guo et al.52.
PCR amplification and DNA sequencing
As an initial analysis of genetic diversity, the glyceraldehydes-3-phosphate dehydrogenase (GAPDH) gene was amplified from the isolates in this study with the primers GDF/GDR53. Fifty-two isolates representing wide ranges of genetic diversity and geographic origins were selected for further investigation.
The nuclear rDNA ITS region and the β-tubulin (TUB2), partial actin (ACT) and calmodulin (CAL) genes were amplified from 52 representative isolates using the primers ITS1/ITS454,55, Bt2a/Bt2b56, ACT512F/ACT783R57 and CL1/CL2A58, respectively. PCR was performed under the conditions described by Prihastuti et al.26.
The amplifications were performed in a 40 μl mixture containing 17 μl ddH2O, 20 μl 2 × PCR MasterMix (TIANGEN Co., China), 1 μl DNA template (30–50 ng/μl), and 1 μl of each primer (10 μM). DNA sequencing was performed by Sangon Biotech Co., Ltd. (Shanghai, China).
Phylogenetic analysis
Alignment of the GAPDH genes of all of the isolates was performed using Clustal X59. MEGA v. 5 was used to build a distance tree with the neighbour-joining (NJ) algorithm. The sequences were compared with those in the NCBI sequence database using the BLAST algorithm for approximate identification.
The sequences of the 52 isolates and the reference sequences obtained from GenBank (Table 3) were aligned using Clustal X. Then, a phylogenetic tree was constructed with the combined ITS, TUB2, ACT, GAPDH and CAL dataset.
Parsimony trees were inferred by PAUP v4.0b10 using a heuristic search option with 1,000 random sequence additions60. All gaps were treated as missing data. Max trees were unlimited, zero-length branches were collapsed, and all multiple parsimonious trees were saved. Clade stability was assessed by bootstrap (BT) analysis with 1,000 replicates. In addition, descriptive tree statistics, such as parsimony (Tree Length [TL], Consistency Index [CI], Retention Index [RI], Related Consistency Index [RC] and Homoplasy Index [HI]), were calculated.
Pathogenicity tests
Pears were included in the pathogenicity tests for two main reasons: i) because peppers often are planted in pear orchards; and ii) to assess whether Colletotrichum species from pepper are host specific. Fruits of Capsicum annuum (Ca. annuum var. dactylus M and Ca. annuum L. var. conoides (Mill.) Irish) and Pyrus pyrifolia were surface-sterilized in 75% ethanol for 3 min and then rinsed three times in sterile distilled water. The fruits were stabbed lightly with a sterile needle, and a mycelial disc with a diameter of 5 mm from a 4-day-old colony obtained from an isolate grown on PDA at 27 °C was attached to each artificially wounded fruit. The PDA discs were covered with moistened cotton for 3 days. The cotton was then removed, and the fruits were incubated for 14 days in a growth chamber at 27 °C with a 12 h light/12 h dark cycle. Six replicates and an equal number of control fruits inoculated only with agar discs were included.
Additional Information
How to cite this article: Liu, F. et al. Molecular and phenotypic characterization of Colletotrichum species associated with anthracnose disease in peppers from Sichuan Province, China. Sci. Rep. 6, 32761; doi: 10.1038/srep32761 (2016).
References
Hyde, K. D. et al. Colletotrichum-names in current use. Fungal Divers. 39, 147–182 (2009).
Than, P. P. et al. Characterization and pathogenicity of Colletotrichum species associated with anthracnose on chilli (capsicum spp.) in Thailand. Plant Pathol. 57, 562–572 (2008).
Damm, U., Woudenberg, J. H. C., Cannon, P. F. & Crous, P. W. Colletotrichum species with curved conidia from herbaceous hosts. Fungal Divers. 39, 45–87 (2009).
Montri, P., Taylor, P. W. J. & Mongkolporn, O. Pathotypes of Colletotrichum capsici, the causal agent of chili anthracnose, in Thailand. Plant Dis. 93, 17–20 (2009).
Li, N. Study on the species of anthracnose pathogens and the groups genetic diversity, Master’s thesis (Sichuan Agricultural University, 2012).
Tang, J. M. A study on pathogens identification of pepper fruit anthracnose and their biological characteristics in Guangxi, Master’s thesis (Guangxi University, 2012).
Yang, Y. L. Multi-locus phylogeny of Colletotrichum species in Guizhou, Yunnan and Guangxi, China. PhD thesis (Huazhong Agricultural University, 2010).
Anderson, J. M., Aitken, E. A. B., Dann, E. K. & Coates, L. M. Morphological and molecular diversity of Colletotrichum spp. Causing pepper spot and anthracnose of lychee (litchi chinensis) in Australia. Plant Pathol. 62, 279–288 (2013).
Ramdial, H. & Rampersad, S. N. Characterization of Colletotrichum spp. Causing anthracnose of bell pepper (Capsicum annuum L.) in Trinidad. Phytoparasitica 43, 37–49 (2014).
Xia, H., Wang, X. L., Zhu, H. J. & Gao, B. D. First report of anthracnose caused by Glomerella acutata on chili pepper in China. Plant Dis. 95, 219 (2011).
Harp, T., Kuhn, P., Roberts, P. D. & Pernezny, K. L. Management and cross-infectivity potential of Colletotrichum acutatum causing anthracnose on bell pepper in Florida. Phytoparasitica 42, 31–39 (2014).
Shin, H. J., Xu, T., Zhang, C. L. & Cheng, Z. J. The comparative study of capsicum anthracnose pathogens from Korea with that of China. Journal of Zhejiang University 26, 629–634 (2000).
Harp, T. L. et al. The etiology of recent pepper anthracnose outbreaks in Florida. Crop Protect. 27, 1380–1384 (2008).
Than, P. P. et al. Chilli anthracnose disease caused by Colletotrichum species. J. Zhejiang U. Sci. 9, B 9, 764–778 (2008).
Sharma, P. N. et al. First report on association of Colletotrichum coccodes with chili anthracnose in India. Plant Dis. 95, 1584–1584 (2011).
Phoulivong, S. et al. Colletotrichum gloeosporioides is not a common pathogen on tropical fruits. Fungal Divers. 44, 33–43 (2010).
Sharma, G. & Shenoy, B. D. Colletotrichum fructicola and C. Siamense are involved in chilli anthracnose in India. Arch. Phytopathol. Plant Protect. 47, 1179–1194 (2014).
Weir, B. S., Johnston, P. R. & Damm, U. The Colletotrichum gloeosporioides species complex. Stud. Mycol. 73, 115–180 (2012).
Diao, Y. Z., Fan, J. R., Wang, Z. W. & Liu, X. L. First report of Colletotrichum boninense causing anthracnose on pepper in China. Plant Dis. 97, 138–138 (2013).
Tozze, H. J. Jr. et al. First report of Colletotrichum boninense causing anthracnose on pepper in Brazil. Plant Dis. 93, 106–106 (2009).
Kanto, T. et al. Anthracnose of sweet pepper caused by Colletotrichum scovillei in Japan. J. Gen. Plant Pathol. 80, 73–78 (2014).
Zhang, G. Z. et al. Identification of pepper anthracnose and resistant screen of breeding materials in Sichuan. Southwest China Journal of Agricultural Sciences 26, 1026–1029 (2013).
Lima, N. B. et al. Five Colletotrichum species are responsible for mango anthracnose in northeastern Brazil. Fungal Divers. 61, 75–88 (2013).
Tao, G. et al. Endophytic Colletotrichum species from Bletilla ochracea (Orchidaceae), with descriptions of seven new speices. Fungal Divers. 61, 139–164 (2013).
Cai, L. et al. A polyphasic approach for studying Colletotrichum. Fungal Divers. 39, 183–204 (2009).
Prihastuti, H. et al. Characterization of Colletotrichum species associated with coffee berries in northern Thailand. Fungal Divers. 39, 89 (2009).
Yang, Y. L. et al. Colletotrichum anthracnose of Amaryllidaceae. Fungal Divers. 39, 123–146 (2009).
Niu X. et al. Colletotrichum species associated with jute (Corchorus capsularis L.) anthracnose in southeastern China. Sci. Rep. 6, 25179, 10.1038/srep25179 (2016).
Manamgoda, D. S. et al. Endophytic Colletotrichum from tropical grasses with a new species C. endophytica. Fungal Divers. 61, 107–115 (2013).
Simmonds, J. H. A study of the species of Colletotrichum causing ripe fruit rots in Queensland. Queensland Journal of Agricultural and Animal Science 22, 437–459 (1965).
Denoyes, B. & Baudry, A. Species identification and pathogenicity study of French Colletotrichum strains isolated from strawberry using morphological and cultural characteristics. Phytopathology 85, 53–57 (1995).
Crouch, J. A., Clarke, B. B., White, J. F. & Hillman, B. I. Systematic analysis of the falcate-spored graminicolous Colletotrichum and a description of six new species from warm-season grasses. Mycologia 101, 717–732 (2009).
Du, M., Schardl, C. L., Nuckles, E. M. & Vaillancourt, L. J. Using mating-type gene sequences for improved phylogenetic resolution of Collectotrichum species complexes. Mycologia 97, 641–658 (2005).
Sawant, I. S. et al. Emergence of Colletotrichum gloeosporioides sensu lato as the dominant pathogen of anthracnose disease of grapes in India as evidenced by cultural, morphological and molecular data. Australasian Plant Pathol. 41, 493–504 (2012).
Noireung, P. et al. Novel species of Colletotrichum revealed by morphology and molecular analysis. Cryptogam, Mycol. 33, 347–362 (2012).
Huang, F. et al. Colletotrichum species associated with cultivated citrus in China. Fungal Divers. 61, 61–74 (2013).
Liu, F., Damm, U., Cai, L. & Crous, P. W. Species of the Colletotrichum gloeosporioides complex associated with anthracnose diseases of Proteaceae. Fungal Divers. 61, 89–105 (2013).
Udayanga, D. et al. What are the common anthracnose pathogens of tropical fruits? Fungal Divers. 61, 165–179 (2013).
Vieira, W. A. S. et al. Endophytic species of Colletotrichum associated with mango in northeastern Brazil. Fungal Divers. 67, 181–202 (2014).
Penzig, D. O. Funghi agrumicoli. Contribuzione allo studio dei funghi parassiti degli agrumi, Vol. 2, Michelia (1882).
Rueda-Hernández, K. R. et al. Differential organ distribution, pathogenicity and benomyl sensitivity of Colletotrichum spp. From blackberry plants in Northern Colombia. J. Phytopathol. 161, 246–253 (2013).
Shenoy, B. D., Jeewon, R. & Lam, W. H. Morpho-molecular characterisation and epitypification of Colletotrichum capsici (Glomerellaceae, Sordariomycetes), the causative agent of anthracnose in chilli. Fungal Divers. 27, 197–211 (2007).
Pring, R. J., Nash, C., Zakaria, M. & Bailey, J. A. Infection process and host range of Colletotrichum capsici. Physiol. Mol. Plant Pathol. 46, 137–152 (1995).
Chai, A. et al. Identification of Colletotrichum capsici (Syd.) butler causing anthracnose on pumpkin in China. Can. J. Plant Pathol. 36, 121–124 (2014).
Víchová, J., Stanková, B. & Pokorný, R. First report of Colletotrichum acutatum on tomato and apple fruits in the Czech Republic. Plant Dis. 96, 769–769 (2012).
Samuelian, S. K., Greer, L. A., Savocchia, S. & Steel, C. C. Application of Cabrio (a.i. pyraclostrobin) at flowering and veraison reduces the severity of bitter rot (Greeneria uvicola) and ripe rot (Colletotrichum acutatum) of grapes. Aust. J. Grape Wine Res. 20, 292–298 (2014).
Damm, U., Cannon, P. F., Woudenberg, J. H. & Crous, P. W. The Colletotrichum acutatum species complex. Stud. Mycol. 73, 37–113 (2012).
Nirenberg, H. I., Feiler, U. & Hagedorn, G. Description of Colletotrichum lupini comb. Nov. inmodern terms. Mycologia 94, 307–320 (2002).
Vieira, W. A. S. et al. First report of papaya fruit anthracnose caused by Colletotrichum brevisporum in Brazil. Plant Dis. 97, 1659 (2013).
Gong, G. S. et al. A simple method for single fungal spore isolation. Journal of Maize Sciences 18, 126–127, 134 (2010).
Sutton, B. C. The Coelomycetes. Fungi imperfecti with pycnidia, acervuli and stromata (Commonwealth Mycological Institute, 1980).
Guo, L. D., Hyde, K. D. & Liew, E. C. Y. Identification of endophytic fungi from Livistona chinensis based on morphology and rDNA sequences. New Phytol. 147, 617–630 (2000).
Templeton, M. D., Rikkerink, E. H., Solon, S. L. & Crowhurst, R. N. Cloning and molecular characterization of the glyceraldehyde-3-phosphate dehydrogenase-encoding gene and cDNA from the plant pathogenic fungus Glomerella cingulata. Gene 122, 225–230 (1992).
White, T. J., Bruns, T., Lee, S. & Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. PCR Protocols: A Guide to Methods and Applications 18, 315–322 (1990).
Gardes, M. & Bruns, T. D. ITS primers with enhanced specificity for basidiomycetes–application to the identification of mycorrhizae and rusts. Mol. Ecol. 2, 113–118 (1993).
Glass, N. L. & Donaldson, G. C. Development of primer sets designed for use with the PCR to amplify conserved genes from filamentous ascomycetes. Appl. Environ. Microbiol. 61, 1323–1330 (1995).
Carbone, I. & Kohn, L. M. A method for designing primer sets for speciation studies in filamentous ascomycetes. Mycologia 91, 553–556 (1999).
O’Donnell, K., Nirenberg, H. I., Aoki, T. & Cigelnik, E. A multigene phylogeny of the Gibberella fujikuroi species complex: detection of additional phylogenetically distinct species. Mycoscience 41, 61–78 (2000).
Thompson, J. D. et al. The CLUSTAL_X windows interface: flexible strategies for multiple sequence alignment aided by quality analysis tools. Nucleic Acids Res. 25, 4876–4882 (1997).
Swofford. PAUP* Beta10. phylogenetic analysis using parsimony (*and other methods). Version 4b10 (Sinauer Associates, 2002).
Acknowledgements
This study was funded by the Two-Way Support Project of Sichuan Agricultural University. We are grateful to our team at the Crop Disease Laboratory for helping to collect the samples.
Author information
Authors and Affiliations
Contributions
F.L.L. and G.S.G. conceived the experiments and were the main authors. F.L.L., G.S.G. and G.T.T. conducted and performed the experiments. F.L.L., G.S.G., G.T.T., X.J.Z., J.X., X.L.C., Y.L., X.F.S., X.B.Q. and Y.Z. analysed the results. G.S.G., H.B.C., S.R.Z., X.F.S. and X.B.Q. collected the samples. All authors reviewed the manuscript.
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Rights and permissions
This work is licensed under a Creative Commons Attribution 4.0 International License. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in the credit line; if the material is not included under the Creative Commons license, users will need to obtain permission from the license holder to reproduce the material. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/
About this article

Cite this article
Liu, F., Tang, G., Zheng, X. et al. Molecular and phenotypic characterization of Colletotrichum species associated with anthracnose disease in peppers from Sichuan Province, China. Sci Rep 6, 32761 (2016). https://doi.org/10.1038/srep32761
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/srep32761
This article is cited by
-
What’s in my Pot? Six Colletotrichum Species Causing Anthracnose in Brazilian Pecan Orchards
Current Microbiology (2024)
-
Polyphasic identification of two novel Colletotrichum species causing leaf spots on mangroves in Thailand
European Journal of Plant Pathology (2024)
-
Identification, prevalence and pathogenicity of Colletotrichum species associated with chilli anthracnose in India
Journal of Plant Pathology (2023)
-
A new approach to quantify anthracnose symptoms in inoculated Capsicum spp. fruits
Tropical Plant Pathology (2022)
-
First report of postharvest anthracnose of mango fruit caused by Colletotrichum plurivorum in China
Journal of Plant Pathology (2022)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
