
Abstract
Caenorhabditis elegans is an important non-mammalian alternative assay model for toxicological study. Previous study has indicated that exposure to multi-walled carbon nanotubes (MWCNTs) dysregulated the transcriptional expression of mir-259. In this study, we examined the molecular basis for mir-259 in regulating MWCNTs toxicity in nematodes. Mutation of mir-259 induced a susceptible property to MWCNTs toxicity and MWCNTs exposure induced a significant increase in mir-259::GFP in pharyngeal/intestinal valve and reproductive tract, implying that mir-259 might mediate a protection mechanisms for nematodes against MWCNTs toxicity. RSKS-1, a putative ribosomal protein S6 kinase, acted as the target for mir-259 in regulating MWCNTs toxicity and mutation of rsks-1 suppressed the susceptible property of mir-259 mutant to MWCNTs toxicity. Moreover, mir-259 functioned in pharynx-intestinal valve and RSKS-1 functioned in pharynx to regulate MWCNTs toxicity. Furthermore, RSKS-1 regulated MWCNTs toxicity by suppressing the function of AAK-2-DAF-16 signaling cascade. Our results will strengthen our understanding the microRNAs mediated protection mechanisms for animals against the toxicity from certain nanomaterials.
Similar content being viewed by others
Introduction
Multi-walled carbon nanotubes (MWCNTs), one member of the carbon nanotubes (CNTs), have numerous unique physicochemical properties. So far, MWCNTs have been produced in bulk for diverse purposes. With the increase in MWCNTs manufacture, it is likely that the increasing exposure of human and environmental organisms to MWCNTs will occur1,2. The potential toxic effects of ENMs including the MWCNTs on organisms have received the great attentions3,4,5. It has been shown that exposure to CNTs can lead to several aspects of toxicity on organisms through the induction of oxidative stress and/or inflammation and crossing the biological barriers6,7,8,9. To determine the underlying molecular mechanisms of MWCNTs toxicity, some dysregulated genes or microRNAs (miRNAs) have been identified in mice or NIH/2Ts cells10,11. miRNAs, a class of short noncoding RNAs, usually exhibit their biological functions by post-transcriptionally inhibiting the expression of targeted genes12.
Due to the properties of short lifespan, ease of manipulation and especially the well-described genetic background13,14,15, the important non-mammalian alternative toxicity assay model of Caenorhabditis elegans has been recently used in the toxicological study of MWCNTs. In nematodes, MWCNTs exposure could cause the damage on the functions of both primary targeted organs such as intestine and secondary targeted organs such as neurons and reproductive organs16,17,18,19. MWCNTs could be further translocated into the secondary targeted organs such as the reproductive organs in nematodes16,17,20. Moreover, based on the SOLiD sequencing, some dysregulated miRNAs have been identified in MWCNTs exposed nematodes21. Biological functions of some dysregulated miRNAs in regulating MWCNTs toxicity have been confirmed with the aid of the available mutants21. Nevertheless, the molecular signaling pathways mediated by these candidate miRNAs in regulating MWCNTs toxicity are still largely unclear in nematodes.
Previous study has demonstrated that MWCNTs exposure increased the expression of mir-259 in nematodes21. In nematodes, mutation of mir-259 induced a susceptible property to MWCNTs toxicity21. The MWCNTs (1 mg/L) exposed mir-259(n4106) mutant exhibited a more significant reduction in brood size, decrease in locomotion behavior and induction of intestinal autofluorescence or reactive oxygen species (ROS) production than MWCMTs exposed wild-type nematodes21. However, the underlying epigenetic mechanism for mir-259 in regulating MWCNTs toxicity is still unclear. In this study, we investigated the molecular signaling mediated by mir-259 in regulating MWCNTs toxicity in the in vivo assay system of C. elegans. In nematodes, rsks-1 gene encodes a putative ribosomal protein S6 kinase (S6K) that is required for the longevity control22, aak-2 gene encodes a catalytic alpha subunit of AMP-activated protein kinases (AMPKs) and daf-16 gene encodes a FOXO transcriptional factor in the insulin signaling pathway. We raised a RSKS-1-AAK-2-DAF-16 signaling cascade mediated by mir-259 in the control of MWCNTs toxicity in nematodes. Our study reveals the key function of mir-259 mediated signaling cascade in encoding a protection mechanism for nematodes against the MWCNTs toxicity.
Results
Effects of MWCNTs exposure on lifespan and locomotion behavior during aging in mir-259 mutant nematodes
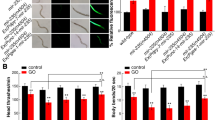
Lifespan can reflect the long-term effect of certain toxicants on animals. mir-259(n4106) has the similar lifespan to that in wild-type nematodes (Fig. 1a). After prolonged exposure, MWCNTs (1 mg/L) exposed mir-259(n4106) mutant exhibited the significantly reduced lifespan than MWCNTs (1 mg/L) exposed wild-type nematodes (Fig. 1a).

Effects of MWCNTs exposure on lifespan and locomotion behavior during aging in nematodes.
(a) Effects of MWCNTs exposure on lifespan in nematodes. (b) Effects of MWCNTs exposure on locomotion behavior during aging in nematodes. Exposure concentration of MWCNTs was 1 mg/L. Prolonged exposure was performed from L1-larvae to young adults. Bars represent means ± SD. **P < 0.01 vs control (if not specially indicated).
Moreover, we selected the endpoint of locomotion behavior to assess the change of aging-related property during aging in MWCNTs exposed nematodes23. mir-259(n4106) has the similar head thrash and body bend to those in wild-type nematodes (Fig. 1b). After prolonged exposure, MWCNTs (1 mg/L) exposed mir-259(n4106) mutant showed the significantly decreased head thrash or body bend at adult day-8 than MWCNTs (1 mg/L) exposed wild-type nematodes (Fig. 1b). Therefore, MWCNTs exposure may cause the adverse effects on lifespan and aging related properties such as locomotion behavior during aging in nematodes.
Effect of MWCNTs exposure on spatial expression of mir-259 in nematodes
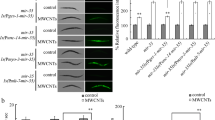
With the aid of transgenic strain of maIs268[mir-259::GFP], we investigated the effect of MWCNTs exposure on spatial expression of mir-259::GFP. In nematodes, mir-259 is expressed in pharyngeal/intestinal valve and reproductive tract24. After prolonged exposure, MWCNTs (1 mg/L) significantly increased the fluorescence intensity of mir-259::GFP in pharyngeal/intestinal valve and reproductive tract compared in nematodes (Fig. 2).

Effects of MWCNTs exposure on mir-259::GFP expression in nematodes.
Arrowheads indicate pharyngeal/intestinal valve in the head and reproductive tract in the mid-region, respectively. Exposure concentration of MWCNTs was 1 mg/L. Prolonged exposure was performed from L1-larvae to young adults. Bars represent means ± SD. **P < 0.01 vs control.
Tissue-specific activity of mir-259 in regulating MWCNTs toxicity in nematodes
With the aid of tissue-specific promoters, we next investigated the tissue-specific activity of mir-259 in regulating MWCNTs toxicity in nematodes. Rescue assay by expression of mir-259 in intestine, pharynx, or reproductive tract did not significantly influence the susceptible property of mir-259(n4106) mutant nematodes to MWCNTs toxicity on lifespan (Fig. 3). In contrast, expression of mir-259 in pharynx/intestinal valve could significantly suppress the susceptible property of mir-259(n4106) mutant nematodes to MWCNTs toxicity on lifespan (Fig. 3). These results imply that mir-259 may act in the pharynx/intestinal valve to regulate MWCNTs toxicity in nematodes.

Tissue-specific activity of mir-259 in regulating MWCNTs toxicity on lifespan in nematodes.
Exposure concentration of MWCNTs was 1 mg/L. Prolonged exposure was performed from L1-larvae to young adults. Bars represent means ± SD. **P < 0.01 vs N2 (if not specially indicated).
rsks-1 might act as the potential targeted gene for mir-259 in nematodes
Using the TargetScan tool, we found that mir-259 may function as an upstream regulator for rsks-1 gene by binding its 3′-UTR. In the loss-of-function mir-259(n4106) mutant, the expression of rsks-1 gene was significantly increased compared with that in wild-type nematodes (Fig. 4a), implying that mir-259 may inhibit the expression of rsks-1 gene.

Effects of rsks-1 mutation on MWCNTs toxicity in nematodes.
(a) Effect of mir-259 mutation on expression of rsks-1 gene. Bars represent means ± SD. **P < 0.01 vs N2. (b) Effects of rsks-1 mutation on MWCNTs toxicity in reducing lifespan in nematodes. Bars represent means ± SD. **P < 0.01 vs control (if not specially indicated). (c) Effects of rsks-1 mutation on MWCNTs toxicity in decreasing locomotion behavior in nematodes. Bars represent means ± SD. **P < 0.01 vs control (if not specially indicated). Exposure concentration of MWCNTs was 1 mg/L. Prolonged exposure was performed from L1-larvae to young adults.
RSKS-1 was involved in the control of MWCNTs toxicity in nematodes
Using the loss-of-function rsks-1(ok1255) mutant, we investigated the potential function of RSKS-1 in the control of MWCNTs toxicity in nematodes. The rsks-1(ok1255) mutant has the similar lifespan and locomotion behavior to those in wild-type nematodes (Fig. 4b,c). After prolonged exposure, the MWCNTs (1 mg/L) exposed rsks-1(ok1255) mutant exhibited the similar lifespan and locomotion behavior at adult day-8 to those in rsks-1(ok1255) mutant or wild-type nematodes without MWCNTs exposure (Fig. 4b,c). That is, the rsks-1(ok1255) mutant had the resistant property to MWCNTs toxicity in nematodes.
Genetic interaction between mir-259 and rsks-1 in regulating MWCNTs toxicity in nematodes
To confirm the interaction between mir-259 and rsks-1 in regulating MWCNTs toxicity, we compared the MWCNTs toxicity in double mutant of rsks-1(ok1255);mir-259(n4106) with that in single mutant of mir-259(n4106) or rsks-1(ok1255). After MWCNTs (1 mg/L) exposure, the lifespan and locomotion behavior at adult day-8 in double mutant of rsks-1(ok1255);mir-259(n4106) were similar to those in single mutant of rsks-1(ok1255) (Fig. 5), implying that the susceptible property of mir-259(n4106) mutant to MWCNTs toxicity on lifespan and aging-related properties could be suppressed by rsks-1 mutation in nematodes. Therefore, mir-259 may inhibit the function of RSKS-1 in positively regulating the MWCNTs toxicity in nematodes.

Genetic interaction between mir-259 and rsks-1 in regulating MWCNTs toxicity in nematodes.
(a) Genetic interaction between mir-259 and rsks-1 in regulating MWCNTs toxicity in reducing lifespan in nematodes. (b) Genetic interaction between mir-259 and rsks-1 in regulating MWCNTs toxicity in decreasing locomotion behavior in nematodes. Exposure concentration of MWCNTs was 1 mg/L. Prolonged exposure was performed from L1-larvae to young adults. Bars represent means ± SD. **P < 0.01 vs N2 (if not specially indicated).
Tissue-specific activity of rsks-1 in regulating MWCNTs toxicity in nematodes
In C. elegans, rsks-1 gene is expressed in pharynx and hypodermis16. Using the tissue-specific promoters, we investigated the tissue-specific activity of rsks-1 in regulating MWCNTs toxicity in nematodes. Rescue assay by expression of rsks-1 in hypodermis did not significantly influence the lifespan in MWCNTs (1 mg/L) exposed rsks-1(ok1255) mutant nematodes (Fig. 6). In contrast, expression of rsks-1 in pharynx could significantly decrease the lifespan in MWCNTs (1 mg/L) exposed rsks-1(ok1255) mutant nematodes (Fig. 6). These results suggest that RSKS-1 may act in the pharynx to regulate MWCNTs toxicity in nematodes.

Tissue-specific activity of rsks-1 in regulating MWCNTs toxicity on lifespan in nematodes.
Exposure concentration of MWCNTs was 1 mg/L. Prolonged exposure was performed from L1-larvae to young adults. Bars represent means ± SD. **P < 0.01 vs N2 (if not specially indicated).
Genetic interaction between rsks-1 and aak-2 in regulating MWCNTs toxicity in nematodes
Previous study has implied that AAK-2 may act as an important molecular target for RSKS-1 in regulating longevity22. To determine whether RSKS-1 can act though the AAK-2 mediated signaling to regulate MWCNTs toxicity, we compared the MWCNTs toxicity in double mutant of rsks-1(ok1255);aak-2(ok524) with that in single mutant of rsks-1(ok1255) or aak-2(ok524). After MWCNTs (1 mg/L) exposure, the lifespan and locomotion behavior at adult day-8 in double mutant of rsks-1(ok1255);aak-2(ok524) were similar to those in aak-2(ok524) mutant nematodes (Fig. 7), suggesting that the resistant property of rsks-1(ok1255) mutant to MWCNTs toxicity on lifespan and aging-related properties could be inhibited by aak-2 mutation in nematodes. These results imply that RSKS-1 may genetically act upstream of AAK-2 to regulate MWCNTs toxicity in nematodes.

Genetic interaction between rsks-1 and aak-2 in regulating MWCNTs toxicity in nematodes.
(a) Genetic interaction between rsks-1 and aak-2 in regulating MWCNTs toxicity in reducing lifespan in nematodes. (b) Genetic interaction between rsks-1 and aak-2 in regulating MWCNTs toxicity in decreasing locomotion behavior in nematodes. Exposure concentration of MWCNTs was 1 mg/L. Prolonged exposure was performed from L1-larvae to young adults. Bars represent means ± SD. **P < 0.01 vs N2 (if not specially indicated).
Genetic interaction between aak-2 and daf-16 in regulating MWCNTs toxicity in nematodes
Previous study has further suggested AAK-2 may function upstream of DAF-16 in insulin signaling pathway to regulate biological processes such as longevity in nematodes22. To determine the genetic interaction between aak-2 and daf-16 in regulating MWCNTs toxicity, we compared the MWCNTs toxicity in double mutant of daf-16(mu86);aak-2(om524) with that in single mutant of daf-16(mu86) or aak-2(om524). After MWCNTs (1 mg/L) exposure, we found that the lifespan and locomotion behavior at adult day-8 in double mutant of daf-16(mu86);aak-2(om524) were similar to those in single mutant of aak-2(om524) or daf-16(mu86) nematodes (Fig. 8), implying that AAK-2 can act together with DAF-16 in the same genetic pathway to regulate the MWCNTs toxicity in nematodes.

Genetic interaction between aak-2 and daf-16 in regulating MWCNTs toxicity in nematodes.
(a) Genetic interaction between aak-2 and daf-16 in regulating MWCNTs toxicity in reducing lifespan in nematodes. (b) Genetic interaction between aak-2 and daf-16 in regulating MWCNTs toxicity in decreasing locomotion behavior in nematodes. Exposure concentration of MWCNTs was 1 mg/L. Prolonged exposure was performed from L1-larvae to young adults. Bars represent means ± SD. **P < 0.01 vs N2.
Distribution and translocation of MWCNTs in mir-259, rsks-1, aak-2 and daf-16 mutant nematodes
Biodistribution and translocation are crucial factors for the toxicity formation of ENMs in nematodes15. With the aid of molecular probe of Rhodamine B (Rho B), we prepared the MWCNTs/Rho B. After MWCNTs/Rho B exposure, we observed a more pronounced MWCNTs/Rho B distribution in the body of mir-259(n4106), aak-2(ok524) and daf-16(mu86) mutants compared with wild-type N2 (Fig. 9). In contrast, mutation of rsks-1 gene significantly suppressed the distribution of MWCNTs/Rho B in the body of nematodes compared with wild-type N2 (Fig. 9). Exposure to Rho B caused the relatively equal distribution of fluorescence in the tissues of wild-type N2, mir-259(n4106), rsks-1(ok1255), aak-2(ok524), or daf-16(mu86) mutant nematodes (Fig. S1). Therefore, mutation of the mir-259, aak-2, or daf-16 enhanced the biodistribution and translocation of MWCNTs in the body, whereas mutation of rsks-1 suppressed the accumulation of MWCNTs in the body of nematodes.

Distribution and translocation of MWCNTs/Rho B in nematodes.
Arrowheads indicate the intestine. Exposure concentration of MWCNTs/Rho B was 1 mg/L. Prolonged exposure was performed from L1-larvae to young adults. Bars represent means ± SD. **P < 0.01 vs N2.
Discussion
Our previous study has demonstrated that, after MWCNTs exposure, loss-of-function mutation of mir-259 could cause the more significant reduction in brood size, decrease in locomotion behavior and induction of intestinal autofluorescence or ROS production compared with wild-type nematodes21. In this study, after MWCNTs exposure, we further observed that mutation of mir-259 could result in the more reduced lifespan and more severely decreased locomotion behavior during the aging (Fig. 1). Therefore, our results suggest that, besides the functions of primary and secondary targeted organs, MWCNTs exposure may further potentially adversely affect the longevity and aging related phenotypes in organisms.
Using the transgenic strain of maIs268, we observed that MWCNTs could significantly increase the expression of mir-259::GFP in both the pharyngeal/intestinal valve and the reproductive tract (Fig. 2). Meanwhile, we found that the loss-of-function mir-259 mutant was susceptible to MWCNTs toxicity in nematodes21 (Fig. 1). These data imply that mir-259 may mediate a protection mechanism for nematodes against the MWCNTs toxicity. That is, MWCNTs may induce a protection mechanism encoded by the activated mir-259 in nematodes. Similarly, previous study has demonstrated that the activated mir-360 may encode a protection mechanism for nematodes against the toxic effects of graphene oxide (GO) in inducing germline apoptosis25.
Previous studies have suggested that some important signaling pathways can act in certain tissues to regulate the toxicity of ENMs in nematodes. For example, acs-22 gene encoding a protein homologous to mammalian fatty acid transport protein 4 could act in the intestine to regulate MWCNTs toxicity20 and insulin signaling pathways could act in the intestine to regulate the GO toxicity26. unc-30 gene encoding a homeodomain transcription factor could act in the RMEs or D-type GABAergic motor neurons to regulate the neurotoxicity of quantum dots (QDs)27,28. In this study, using the tissue-specific promoters, we found that the mir-259 acted in the pharynx/intestinal valve to regulate MWCNTs toxicity in nematodes (Fig. 3), implying the potential important function of mir-259 in affecting the function of pharynx/intestinal valve. We observed that mutation of mir-259 enhanced the distribution of MWCNTs/Rho B in the body of nematodes (Fig. 9). Moreover, our results further imply the possible important role of pharynx/intestinal valve in the control of toxicity and translocation of certain ENMs in nematodes.
In this study, we provide several lines of evidence to indicate the potential role of RSKS-1 as the target for mir-259 in regulating the MWCMTs toxicity in nematodes. We observed that the phenotypes in MWCNTs exposed rsks-1 mutant were opposite to those in MWCNTs exposed mir-259 mutant. The rsks-1 mutant was resistant to the MWCNTs toxicity on longevity and aging related phenotypes (Fig. 4). Moreover, we found that mutation of the rsks-1 gene could suppress the susceptible property of mir-259(n4106) mutant to MWCNTs toxicity on lifespan and aging-related properties (Fig. 5). Therefore, although we did not exclude the possible involvement of other targets in the control of MWCNTs toxicity, our results indicated that mir-259 can negatively regulate the MWCNTs toxicity by inhibiting the function of RSKS-1, one of its important targets.
For the tissue-specific activity, our results suggest that rsks-1 gene acted in the pharynx to regulate the MWCNTs toxicity on longevity and aging related phenotypes in nematodes (Fig. 6). Based on the distribution pattern of MWCNTs/Rho B, we also found that mutation of rsks-1 significantly suppressed the deposition of MWCNTs in the body of nematodes (Fig. 9). Considering the fact that RSKS-1 acted in the pharynx to regulate the MWCNTs toxicity, the MWCNTs uptake by feeding might be altered in the rsks-1 mutant nematodes. In contrast, the altered MWCNTs translocation into the secondary targeted organs through the intestinal barrier might be not directly due to the rsks-1 mutation in nematodes. That is, our data suggest the important role of rsks-1 gene in the pharynx in regulating the toxicity and accumulation of MWCNTs. Meanwhile, our data also imply the possible crucial role of pharynx in the control of toxicity and accumulation of certain ENMs in nematodes.
In this study, we identified the downstream signaling cascade for RSKS-1 in regulating the MWCNTs toxicity. In C. elegans, aak-2 can function downstream of stressors and energy level signals to positively regulate the adult lifespan29. Here we further showed that AAK-2 functioned downstream of RSKS-1 to regulate the MWCNTs toxicity, since mutation of the aak-2 gene suppressed the resistant property of rsks-1(ok1255) mutant to MWCNTs toxicity on the longevity and the aging related phenotypes (Fig. 7). Moreover, our results suggest that AAK-2 may further regulate the MWCNTs toxicity on longevity and aging related phenotypes by acting with the transcriptional factor DAF-16 in the same genetic pathway (Fig. 8). Therefore, RSKS-1 may act upstream of the AAK-2-DAF-16 signaling cascade to regulate the MWCNTs toxicity in nematodes. The AAK-2-DAF-16 signaling cascade was also shown to be involved in the longevity regulation in nematodes22, implying that the AAK-2-DAF-16 signaling cascade may be a conserved signaling cascade involved in the control of biological processes in nematodes. Nevertheless, RSKS-1, AAK-2 and DAF-16 may not regulate the MWCNTs toxicity by forming a complex, because RSKS-1 acted in the pharynx to regulate the MWCNTs toxicity and DAF-16 normally acted in the intestine to regulate the nanotoxicity and innate immunity26,30. In addition, previous studies have only shown that AAK-2 can physically interact with ICD-1, ICD-2, F49E8.7, or UNC-42 to regulate biological processes in nematodes (http://www.wormbase.org/species/c_elegans/gene/WBGene00020142#01-9g8-10).
In C. elegans, the insulin receptor DAF-2 can reduce the longevity or enhance the toxicity of certain toxicants such as MWCNTs by suppressing the expression and function of DAF-1631,32,33. Our recent study has demonstrated that MWCNTs exposure would inhibit the expression of mir-355, which further induced the toxic effects of MWCNTs on nematodes by suppressing the function of DAF-233. Therefore, on the one hand, MWCNTs may suppress the function of DAF-16 in inducing the toxicity of MWCNTs on nematodes by inhibiting the expression of mir-355; on the other hand, MWCNTs may be also able to potentially enhance the function of DAF-16 in decreasing the toxicity of MWCNTs on nematodes by increasing the expression of mir-259 (Fig. 10).

A diagram showing the mir-259-mediated molecular signaling in the control of MWCNTs toxicity in nematodes.
In conclusion, in this study, we investigated the molecular basis for mir-259 in regulating MWCNTs toxicity. In nematodes, activation of the mir-259 mediated a protection mechanism for animals against the MWCNTs toxicity on longevity and aging related phenotypes. During the control of MWCNTs toxicity, RSKS-1 acted as an important target for mir-259. mir-259 acted in the pharynx-intestinal valve and rsks-1 acted in the pharynx to regulate MWCNTs toxicity. Moreover, RSKS-1 functioned upstream of the AAK-2-DAF-16 signaling cascade and suppressed the function of this signaling cascade to regulate MWCNTs toxicity. The C. elegans mir-259 is the homologue of human miR-21634. Our results highlight the important role of miRNAs mediated protection mechanisms for animals against the adverse effects from environmental toxicants.
Methods
Characterization of MWCNTs
MWCNTs (diameter: 10–20 nm, length: 6–15 μm) were from Shenzhen Nanotech. Port Co. Ltd (Shenzhen, China). MWCNTs morphology in K-medium (50 mM NaCl, 30 mM KCl, 10 mM NaOAc, pH 6.0) was examined by transmission electron microscopy (TEM, JEM-200CX, JEOL, Japan) (Fig. S2a). Length distribution of MWCNTs was shown in Fig. S2b. Fourier transform infrared spectroscopy (FTIR) of MWCNTs was determined (Avatar 370, Thermo Nicolet, USA). The peak at 3367 cm−1 in the spectra of MWCNTs was attributed to -OH groups, the peak at 2850 cm−1 in the spectra of MWCNTs was attributed to C-H (sp3), the peak at 1629 cm−1 in the spectra of MWCNTs was attributed to C-H stretching, the peak at 1053 cm−1 in the spectra of MWCNTs was attributed to C-O stretching and the peak at 2916 cm−1 in the spectra of MWCNTs was attributed to the stretching vibrations of alkyl groups (Fig. S2c). The FTIR spectra of MWCNTs implies the normal chemical structure on the surface of MWCNTs after sonication. Zeta potential of MWCNTs was analyzed by Nano Zetasizer (Nano ZS90, Malvern Instrument, UK). Zeta potential of MWCNTs in K-medium was −33.4 ± 2.5 mV.
C. elegans strains and exposure
Nematode strains used in this study contain wild-type N2, mutants of mir-259(n4106), rsks-1(ok1255), aak-2(ok524), daf-16(mu86), aak-2(ok524);rsks-1(ok1255), daf-16(mu86);aak-2(om524) and rsks-1(ok1255);mir-259(n4106) and transgenic strains of maIs268[mir-259::GFP], mir-259(n4106)Ex(Pmyo-2-mir-259), mir-259(n4106)Ex(Pges-1-mir-259), mir-259(n4106)Ex(Ppie-1-mir-259), mir-259(n4106)Ex(Pref-1-mir-259), rsks-1(ok1255)Ex(Pmyo-2-rsks-1) and rsks-1(ok1255)Ex(Pdpy-7-rsks-1). mir-259(n4106) is a loss-of-function mutation with the deletion. The deletion breakpoints are: GATTATAATGCAAACAACCTGGGGGATC/CAGTATCTTCA…AAGAGCGAAAGT/ACAGTCTCCTCCTTCTTTGCTCACTTCT. rsks-1(ok1255) is a loss-of-function mutation with the deletion of 1700 bp. aak-2(ok524) is a loss-of-function mutation with the deletion of 409 bp. daf-16(mu86) is a loss-of-function mutation with an 11 kb daf-16 genomic deletion that removes nearly all of the winged-helix domain and ~1 kb upstream of the 5′ UTR. Some strains were purchased from Caenorhabditis Genetics Center (funded by NIH Office of Research Infrastructure Programs (P40 OD010440)). Gravid nematodes were maintained on nematode growth medium (NGM) plates seeded with Escherichia coli OP50 and were lysed with a bleaching mixture (0.45 M NaOH, 2% HOCl) after washing animals off the plates into centrifuge tubes13. Synchronous L1-larvae nematodes were prepared as described35.
Exposure and toxicity assessment
MWCNTs were dispersed in K medium to prepare a stock solution (1 mg/mL). The stock MWCNTs solution was sonicated for 30 min (40 kHz, 100 W) and diluted to the used concentration (1 mg/L) with K medium just prior to exposure. Prolonged exposure to MWCNTs was performed from L1-larvae to young adults in 12-well sterile tissue culture plates at 20 °C in the presence of food (OP50). After MWCNTs exposure, the nematodes were used for the toxicity assessment using endpoints of lifespan and locomotion behavior.
Lifespan was assayed at 20 °C basically as described36,37. During the lifespan assay, the hermaphrodite nematodes were transferred daily for the first 7 days of adulthood. Nematodes would be checked every two-day and were scored as dead if they did not move even after repeated taps with a pick. Sixty nematodes were examined per treatment and three replicates were performed.
Endpoints of head thrash and body bend were used to reflect the locomotion behavior of nematodes as described38,39. Head thrash and body bend were assessed under the dissecting microscope by eyes. A head thrash is defined as a change in the direction of bending at the mid body. A body bend is defined as a change in the direction of the part of the nematodes corresponding to the posterior bulb of the pharynx along the y axis, assuming that nematode was traveling along the x axis. Adult day-1 was used as a control for assay of locomotion behavior during aging. Twenty nematodes were examined per treatment and six replicates were performed.
Bioinformatics analysis for targeted gene prediction of mir-259
The corresponding targeted genes for mir-259 were predicted using TargetScan version 6.2 (http://www.targetscan.org/worm_52/). TargetScan is a tool for searching for the presence of conserved sites that match seed region of a miRNA so as to predict the biological targets of certain miRNA.
Reverse-transcription and quantitative real-time polymerase chain reaction (qRT-PCR)
Total RNAs extracted using RNeasy Mini kit (Qiagen) were reverse transcribed using PrimeScript™ RT reagent kit (Takara, Otsu, Shiga, Japan). Purity and concentration of RNAs were evaluated by OD260/280 in a spectrophotometer. cDNA synthesis was performed in a 12.5 μL reaction volume containing 625 ng total RNA, 0.5 mM reverse-transcript primers, 50 mM Tris-HCl, 75 mM KCl, 3 mM MgCl2, 10 mM dithiothreitol, 20 units ribonuclease inhibitor and 100 U reverse transcriptase (Takara, China). After cDNA synthesis, real-time PCR was performed using SYBR Premix Ex Taq™ (Takara) for the amplification of the examined gene. Real-time PCR was performed using primers for target gene of rsks-1 (forward primer, 5′-CCGTTTGTGGGATTCACC-3′; reverse primer, 5′-TGGCTTTCTCGGGCTCTT-3′) and reference gene of tba-1 (forward primer, 5′-TCAACACTGCCATCGCCGCC-3′; reverse primer, 5′-TCCAAGCGAGACCAGGCTTCAG-3′). Relative quantification of targeted gene of rsks-1 in comparison to reference tba-1 gene was determined and the final results were expressed as relative expression ratio between targeted gene and reference gene. All reactions were performed in triplicate.
DNA constructs and germline transformation
To generate entry vector carrying promoter sequence, promoter region for ges-1 gene specially expressed in the intestine, myo-2 gene specially expressed in the pharynx, pie-1 gene expressed in the reproductive tract, ref-1 gene specially expressed in the valve between pharynx and intestine, or dpy-7 gene specially expressed in the hypodermis was amplified by PCR from wild-type C. elegans genomic DNA. These promoter fragments were inserted into pPD95_77 vector in the sense orientation. rsks-1/Y47D3A.16 cDNA or mir-259 was amplified by PCR and inserted into corresponding entry vector carrying the ges-1, myo-2, pie-1, ref-1, or dpy-7 promoter sequence. Germline transformation was performed by coinjecting testing DNA at the concentration of 10–40 μg/mL and marker DNA of Plin-44::gfp at the concentration of 60 μg/mL into the gonad of nematodes as described40.
Distribution and translocation of MWCNTs
To investigate the distribution and translocation of MWCNTs, Rho B was loaded on MWCNTs by mixing Rho B (1 mg/mL, 0.3 mL) with MWCNTs (0.1 mg/mL, 5 mL) as described previously20. The unbound Rho B was removed by dialysis against the distilled water over 72-h. The prepared MWCNTs/Rho B was stored at 4 °C before use. Nematodes were incubated with MWCNTs/Rho B at the concentration of 1 mg/L for 3-h. After incubation, nematodes were washed with M9 buffer for three times. The exposed nematodes were analyzed under a fluorescence microscopy. Rho B treatment was used as the control. Twenty nematodes were examined per treatment.
Statistical analysis
Data were expressed as means ± standard deviation (SD). Graphs were prepared with Microsoft Excel software (Microsoft Corp., Redmond, WA). Statistical analysis was performed using SPSS 12.0 software (SPSS Inc., Chicago, USA) and the differences between groups were determined using analysis of variance (ANOVA). The probability levels of 0.05 and 0.01 were considered to be statistically significant. The lifespan data were analyzed using a 2-tailed 2 sample t-test (Minitab Ltd, Coventry, UK).
Additional Information
How to cite this article: Zhuang, Z. et al. Function of RSKS-1-AAK-2-DAF-16 signaling cascade in enhancing toxicity of multi-walled carbon nanotubes can be suppressed by mir-259 activation in Caenorhabditis elegans. Sci. Rep. 6, 32409; doi: 10.1038/srep32409 (2016).
References
Helland, A., Wick, P., Koehler, A., Schmid, K. & Som, C. Reviewing the environmental and human health knowledge base of carbon nanotubes. Environ. Health Perspect. 115, 1125–1131 (2007).
Simate, G. S., Iyuke, S. E., Ndlovu, S., Heydenrych, M. & Walubita, L. F. Human health effects of residual carbon nanotubes and traditional water treatment chemicals in drinking water. Environ. Int. 39, 38–49 (2012).
Krug, H. F. & Wick P. Nanotoxicology: an interdisciplinary challenge. Angew. Chem. Int. Ed. 50, 1260–1278 (2011).
Krug H. F. Nanosafety research – are we on the right track? Angew. Chem. Int. Ed. 53, 12304–12319 (2014).
Robinson, R. L. M. et al. How should the completeness and quality of curated nanomaterial data be evaluated? Nanoscale 8, 9919–9943 (2016).
Donaldson, K. et al. Carbon nanotubes: a review of their properties in relation to pulmonary toxicology and workplace safety. Toxicol. Sci. 92, 5–22 (2006).
Liu, Y., Zhao, Y., Sun, B. & Chen, C. Understanding the toxicity of carbon nanotubes. Accounts Chem. Res. 46, 702–713 (2013).
Mercer, R. R. et al. Distribution and persistence of pleural penetrations by multi-walled carbon nanotubes. Part. Fibre Toxicol. 7, 28 (2010)
Rothen-Rutishauser, B. et al. Relating the physicochemical characteristics and dispersion of multiwalled carbon nanotubes in different suspension media to their oxidative reactivity in vitro and inflammation in vivo. Nanotoxicology 4, 331–342 (2010).
Pacurari, M. et al. Multi-walled carbon nanotuibe-induced gene expression in the mouse lung: association with lung pathology. Toxicol. Appl. Pharmacol. 255, 18–31 (2011).
Li, S. et al. Assessment of nanomaterial cytotoxicity with SOLiD sequencing-based microRNA expression profiling. Biomaterials 32, 9021–9030 (2011).
Bartel, D. P. MicroRNAs: genomics, biogenesis, mechanism and function. Cell 116, 281–297 (2004).
Brenner, S. The genetics of Caenorhabditis elegans. Genetics 77, 71–94 (1974).
Leung, M. C. et al. Caenorhabditis elegans: an emerging model in biomedical and environmental toxicology. Toxicol. Sci. 106, 5–28 (2008).
Zhao, Y.-L., Wu, Q.-L., Li, Y.-P. & Wang, D.-Y. Translocation, transfer and in vivo safety evaluation of engineered nanomaterials in the non-mammalian alternative toxicity assay model of nematode Caenorhabditis elegans. RSC Adv. 3, 5741–5757 (2013).
Wu, Q.-L. et al. Crucial role of biological barrier at the primary targeted organs in controlling translocation and toxicity of multi-walled carbon nanotubes in nematode Caenorhabditis elegans. Nanoscale 5, 11166–11178 (2013).
Nouara, A. et al. Carboxylic acid functionalization prevents the translocation of multi-walled carbon nanotubes at predicted environmental relevant concentrations into targeted organs of nematode Caenorhabditis elegans. Nanoscale 5, 6088–6096 (2013).
Shu, C.-J. et al. Pretreatment with paeonol prevents the adverse effects and alters the translocation of multi-walled carbon nanotubes in nematode Caenorhabditis elegans. RSC Adv. 5, 8942–8951 (2015).
Eom, H. et al. A systems toxicology approach on the mechanism of uptake and toxicity of MWCNT in Caenorhabditis elegans. Chem.-Biol. Interact. 239, 153–164 (2015).
Zhi, L.-T., Fu, W., Wang, X. & Wang, D.-Y. ACS-22, a protein homologous to mammalian fatty acid transport protein 4, is essential for the control of toxicity and translocation of multi-walled carbon nanotubes in Caenorhabditis elegans. RSC Adv. 6, 4151–4159 (2016).
Zhao, Y.-L. et al. In vivo translocation and toxicity of multi-walled carbon nanotubes are regulated by microRNAs. Nanoscale 6, 4275–4284 (2014).
Chen, D. et al. Germline signaling mediates the synergistically prolonged longevity produced by double mutations in daf-2 and rsks-1 In C. elegans. Cell Rep. 5, 1600–1610 (2013).
Wu, Q.-L., Zhao, Y.-L., Fang, J.-P. & Wang, D.-Y. Immune response is required for the control of in vivo translocation and chronic toxicity of graphene oxide. Nanoscale 6, 5894–5906 (2014).
Martinez, N. J. et al. Genome-scale spatiotemporal analysis of Caenorhabditis elegans microRNA promoter activity. Genome Res. 18, 2005–2015 (2008).
Zhao, Y.-L., Wu, Q.-L. & Wang, D.-Y. An epigenetic signal encoded protection mechanism is activated by graphene oxide to inhibit its induced reproductive toxicity in Caenorhabditis elegans. Biomaterials 79, 15–24 (2016).
Zhao, Y.-L., Yang, R.-L., Rui, Q. & Wang, D.-Y. Intestinal insulin signaling encodes two different molecular mechanisms for the shortened longevity induced by graphene oxide in Caenorhabditis elegans. Sci. Rep. 6, 24024 (2016).
Zhao, Y.-L., Wang, X., Wu, Q.-L., Li, Y.-P. & Wang, D.-Y. Translocation and neurotoxicity of CdTe quantum dots in RMEs motor neurons in nematode Caenorhabditis elegans. J. Hazard. Mater. 283, 480–489 (2015).
Zhao, Y.-L. et al. Quantum dots exposure alters both development and function of D-type GABAergic motor neurons in nematode Caenorhabditis elegans. Toxicol. Res. 4, 399–408 (2015).
Apfeld, J., O’Connor, G., McDonagh, T., DiStefano, P. S. & Curtis, R. The AMP-activated protein kinase AAK-2 links energy levels and insulin-like signals to lifespan in C. elegans. Genes Dev. 18, 3004–3009 (2004).
Partridge, F. A., Gravato-Nobre, M. J. & Hodgkin, J. Signal transduction pathways that function in both development and innate immunity. Dev. Dyn. 239, 1330–1336 (2010).
Shen, L.-L., Du, M., Lin, X.-F., Cai, T. & Wang, D.-Y. Genes required for the functions of olfactory AWA neuron regulate the longevity of Caenorhabditis elegans in an insulin/IGF signaling-dependent fashion. Neurosci. Bull. 26, 91–103 (2010).
Yang, R.-L. et al. Insulin signaling regulates toxicity of traffic-related PM2.5 on intestinal development and function in nematode Caenorhabditis elegans. Toxicol. Res. 4, 333–343 (2015).
Zhao, Y.-L., Yang, J.-N. & Wang, D.-Y. A microRNA-mediated insulin signaling pathway regulates the toxicity of multi-walled carbon nanotubes in nematode Caenorhabditis elegans. Sci. Rep. 6, 23234 (2016).
Ibañez-Ventoso, C., Vora, M. & Driscoll, M. Sequence relationships among C. elegans, D. melanogaster and human microRNAs highlight the extensive conservation of microRNAs in biology. PLoS ONE 3, e2818 (2008).
Donkin, S. & Williams, P. L. Influence of developmental stage, salts and food presence on various end points using Caenorhabditis elegans for aquatic toxicity testing. Environ. Toxicol. Chem. 14, 2139–2147 (1995).
Shakoor, S., Sun, L.-M. & Wang, D.-Y. Multi-walled carbon nanotubes enhanced fungal colonization and suppressed innate immune response to fungal infection in nematodes. Toxicol. Res. 5, 492–499 (2016).
Sun, L.-M. et al. Contribution of heavy metals to toxicity of coal combustion related fine particulate matter (PM2.5) in Caenorhabditis elegans with wild-type or susceptible genetic background. Chemosphere 144, 2392–2400 (2016).
Wu, Q.-L. et al. Inhibition of ROS elevation and damage on mitochondrial function prevents lead-induced neurotoxic effects on structures and functions of AFD neurons in Caenorhabditis elegans. J. Environ. Sci. 24, 733–742 (2012).
Li, Y.-P. et al. High concentration of vitamin E decreases thermosensation and thermotaxis learning and the underlying mechanisms in nematode Caenorhabditis elegans. PLoS ONE 8, e71180 (2013).
Mello, C. & Fire, A. DNA transformation. Methods Cell. Biol. 48, 451–482 (1995).
Acknowledgements
This work was supported by the grant from Fundamental Research Funds for the Central Universities in China (no. 2242016K41059).
Author information
Authors and Affiliations
Contributions
D.W. designed the project. Z.Z., M.L., H.L., L.L., W.G. and Q.W. carried out the experiments. D.W. wrote the manuscript. All authors discussed the results and reviewed the manuscript.
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Electronic supplementary material
Rights and permissions
This work is licensed under a Creative Commons Attribution 4.0 International License. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in the credit line; if the material is not included under the Creative Commons license, users will need to obtain permission from the license holder to reproduce the material. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/
About this article

Cite this article
Zhuang, Z., Li, M., Liu, H. et al. Function of RSKS-1-AAK-2-DAF-16 signaling cascade in enhancing toxicity of multi-walled carbon nanotubes can be suppressed by mir-259 activation in Caenorhabditis elegans. Sci Rep 6, 32409 (2016). https://doi.org/10.1038/srep32409
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/srep32409
This article is cited by
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
