
Abstract
Dens are crucial in the early development of many mammals, making den site selection an important component of parental care in such species. Resource availability and shelter from predators primarily govern den selection. Species inhabiting human-dominated landscapes typically den away from human disturbance, often shifting dens to avoid humans during the early life of their young. Domesticated dogs have evolved in human proximity over centuries, being bred and reared in human homes for generations. While pets rely on their owners for shelter and care, free-ranging dogs roam uncared and typically whelp in dens. We conducted a study on 148 free-ranging dog dens in India to understand their denning habits. Distance from resources influenced den choice, but anthropogenic disturbance did not. Dens were found in areas of high human activity and begging from humans was preferred over scavenging. A study on 15 pregnant females revealed that females actively searched for denning sites, rejecting several intermediate ones before selecting the final den. We propose that the obvious preference of dogs for denning close to humans is a behavioural adaptation that helps them to survive in the urban landscape, in spite of the high human induced mortality during the early life of pups.
Similar content being viewed by others


Introduction
Parental care is defined as “any form of parental behaviour that appears likely to increase the fitness of offspring”1. This broad definition includes a host of behaviours, from allocation of resources for eggs prior to mating and offspring provisioning after birth to nest guarding and suckling1,2. The most basic and perhaps common manifestation of parental care is the building or acquisition of nesting or denning sites that can provide protection to the offspring during the early stages of their development3,4. In mammals, parental care involves suckling which occurs over an extended period of time and demands high energy investment from the mothers5. Carnivorous mammals typically give birth to relatively underdeveloped young that need complete care during their early life for survival4,6,7. In such species, the den plays an important role in the development of the young, providing shelter from the environment, optimal ambient conditions for successful development and protection from predators8,9,10,11. Not surprisingly, predator avoidance and proximity to resources have been shown to be two important factors that drive den site selection in many species8,10,12,13,14,15,16,17,18,19,20,21,22,23,24,25,26.
Rapid urbanization has resulted in fragmentation of habitats available to wildlife worldwide, forcing animals to live in close proximity to humans27,28. Some animals like skunks (Spilogale putorius) in USA and swfit foxes (Vulpes velox) in Canada have adapted to this habitat change by using manmade structures as dens12,29,30, placing their dens in close proximity to roads8,12,30 and/or human settlements12,17,30. Some species, however, avoid humans by selecting den sites away from human activity14,18,31. Thus the urban environment provides ample ground for animal-human conflict as well as mutualism and understanding the denning habits of urban species may contribute to their conservation21,32,33.
Canids are mostly mesocarnivores, often acting as top predators in food chains, have a wide distribution, spanning all continents but Antarctica and occupy diverse habitats, from undisturbed forests to metropolitan areas17,34,35,36,37,38,39,40,41,42,43. Some canids like dholes (Cuon alpinus) and Arctic foxes (Vulpes lagopus) live far from human settlements and rarely interact with humans, but others like gray wolves (Canis lupus lupus), coyotes (Canis latrans) and dingoes (Canis dingo) have adapted to living in and around human habitation20,44,45. The domesticated dog (Canis familiaris) has evolved from gray wolves through a process of domestication. While pet dogs are capable of leading a lifestyle completely dependent on humans46,47,48,49, free-ranging dogs depend on human-generated waste for their sustenance, but are not under direct human supervision50. Nearly 80% of the world’s dog population comprises of free-ranging dogs and these dogs are the best model systems for understanding the unsolved puzzle of dog domestication51,52,53. Pet dogs breed and whelp under human supervision, but show basic denning behaviours like selecting a preferred spot, digging into the ground and tearing bedding material prior to whelping54. They have also been reported to abandon the den suddenly in order to give birth in the proximity of a preferred human54. In contrast, wolves tend to den away from human activities and are intolerant of humans approaching their dens15,31,55,56,57,58,59. Coyotes inhabit human dominated environments, but move their pups to new den sites whenever disturbed by humans60. Foxes also tend to have birthing dens in man-made structures and human dominated landscapes, but in areas where human disturbance is low and they often shift dens when disturbed61,62,63. Thus pet dogs seem to have retained the ancestral habit of denning, but have shifted their preference to den close to preferred humans as a result of domestication. Free-ranging dogs could provide us an insight to this transition in denning behaviour and would contribute to our understanding the various factors that led to the success of dogs in adapting to urban and semi-urban habitats across the globe.
Free-ranging dogs in India are found in all possible human habitations, from forest fringes to metropolitan cities and they are typically scavengers, surviving on scraps and wastes from human kitchens41,53,64. They breed in and around human habitations, and, unlike their wilder relatives, do not avoid human interactions during pup rearing5. Mothers are highly protective and can get quite aggressive while guarding pups65, but shifting of dens as a response to anthropogenic disturbance has not yet been reported in these dogs. On the other hand, there is very high level of human induced mortality in the early life of pups66. Thus humans are a potential threat to the pups and thus the free-ranging dogs in India are a good model system for understanding the choice of denning sites in the face of conflict with humans. In this study we aim to characterize den sites of free-ranging dogs in urban and semi-urban localities and understand the denning preferences of pregnant females. We hypothesize that dogs would prefer denning sites that provide greater protection from predators, less anthropogenic disturbance and easy access to resources.
Results
Den characteristics
Dens were seasonal, most having been located between October and March. Dens could be highly protected and sheltered (inside buildings or other structures), or quite open (located in fields and by the sides of roads). There was no apparent preference of the mothers for sheltered dens (chi-square test: χ2 = 0.014, df = 1, p < 0.907). Pregnant females preferred den sites that gave them access to human provided food (begging), rather than direct scavenging at dustbins and garbage dumping sites (chi-square test: χ2 = 44.938, df = 1, p < 0.0001). Den scores ranged from 9 to 20 (normalized scores 6.25 to 75), with the highest percentage of dens (14.19%) having scores 17 and 18 (normalized scores 56.25 and 62.5) (Please see Supplementary Information 1 for more details). Average litter size was 4.07 ± 1.99. The den score was not correlated with the size of the litter born in the respective dens (linear regression analysis: R2 = 0.003, F = 0.472, p = 0.493), suggesting that the mothers might not be choosing dens based on the expected litter size. However, a 3d plot of frequencies against den scores and litter sizes revealed a pattern of higher incidence of medium to large den scores over small ones, irrespective of litter size (Fig. 1), suggesting that there indeed might be a preference for certain parameters that we used to characterize the dens.

A 3d scatterplot showing the distribution of den scores for various observed litter sizes (N = 148 dens).
The GLMM analysis revealed that the distance of the den from resources like food and water plays a very important role (p = 0.00952) in den selection by the mother (Table 1, Supplementary Information 2). Females that produce small litters tend to select dens that are at an intermediate distance from the resources. On the other hand, females that produce large litters seem to be more flexible in their choice with respect to the distance from resources (Fig. 2a). The structure of the den plays a marginally important role (p = 0.04696) in den selection (Fig. 2b), but the combination of structure and resources is more important (p = 0.01794). Hence it appears that mothers with small litters prefer to be not too close to resources and generally have smaller dens than mothers with large litters. The survivorship analysis revealed that none of the den characteristics affected pup survival (Supplementary Information 3).

(a) Box-whisker plot showing the distance from the den to resources for large (>4 pups) and small (1–4 pups) litters. (b) Box-whisker plot showing the den structure for large (>4 pups) and small (1–4 pups) litters.
Den selection
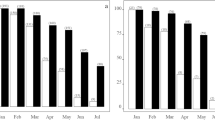
All the 20 pregnant females that were tracked prior to whelping used at least one den site prior to the final den in which the pups were born. 15 of these females could be tracked for a month prior to whelping and they occupied an average of 3.6 ± 0.98 dens during this period. The fifteen pregnant females that were tracked for a month had 2–6 intermediate dens, which they occupied for varied times, ranging from 4 to 21 days during the one month that they were tracked. There was significant variation between the first den score (D1S), average intermediate den score (IDS) and final den scores (DFS) for the 15 females (Kruskal-Wallis test, F = 55.925, df = 2, 41, p < 0.0001); wth D1S < IDS < DFS (Wilcoxon paired-sample test with Bonferroni correction, D1-DF: T = 0.00, N = 15, p < 0.0001; D1-ID: T = 0.00, N = 15, p < 0.0001; ID-DF: T = 1.00, N = 15, p < 0.0001; Fig. 3). The number of intermediate dens used by the mother did not depend on the time available to her before whelping (Linear regression: R2 = 0.196, F = 3.161, p = 0.099), suggesting that the mothers actively selected dens, settling in one that they preferred, irrespective of the number of den sites they sampled.

A bar graph showing the mean ± standard deviation of den scores for 15 pregnant females that were tracked for a month prior to whelping.
The light gray bar represents the first dens (D1), dark gray bar represents the intermediate dens (ID) and the black bar represents the final dens (DF).
The pregnant females covered large distances in search of dens every day (mean ± s.d. = 312.476 ± 182.034 m). The den score difference between the final and first dens (D1DF) increased with the linear distance between the dens (Linear regression: R2 = 0.456, F = 10.908, p = 0.006). D1DF, however, did not increase with the total distance covered over the one month period of observations, considering the actual paths taken by the females (Linear regression: R2 = 0.130, F = 1.938, p = 0.187), but increased with the total number of days spent in intermediate dens (Linear regression: R2 = 0.314, F = 5.944, p = 0.030). The linear distance between the first and final dens increased with an increase in the distance covered in searching (Linear regression: R2 = 0.316, F = 5.998, p = 0.029). Thus, with increased searching effort the pregnant females were more likely to find more suitable dens and they indeed searched actively, settling for the most comfortable den available. Majority of final dens had a score of 17 or 18, for both the pregnant females’ data set and the population level data (Supplementary Information 4, Table S2), suggesting 17–18 to be the optimal den score (Supplementary Information 4, Fig. S1).
Discussion
Free-ranging dogs showed considerable variation in the selection of their denning sites, from open fields to highly protected shelters, small and dark holes to large and well-lit spaces. However, in spite of the large distribution of den scores observed, there seemed to be an optimal den score (scores 17 and 18) which was evident not only from the survey of birthing dens from the population, but also from observations of pregnant females. Unlike wolves and coyotes, which prefer to den away from humans56,60,67,68, even while exploiting resources in human dominated landscapes, free-ranging dogs routinely chose sites of high human activity like markets, railway platforms and residential buildings for dens. Canids typically prefer to den close to water sources67,69,70 and this was also true of the free-ranging dogs. They selected dens close to resources that included water sources as diverse as dripping taps to natural water bodies.
Smaller litters tended to be born at dens that were located at intermediate distances from resources and of relatively smaller size, as compared to the dens of larger litters. Dens with larger litters could be found at variable distances from resources. This could mean that the pregnant females are able to judge whether they would be giving birth to small or large litters. However, it is more likely that the mothers simply select dens in which they are comfortable; being heavier and bigger in size while carrying more pups, they would thus prefer larger dens in spite of their distance from resources, leading to this pattern of den selection, without necessarily involving cognitive abilities of the pregnant females. The pregnant females actively searched for denning sites prior to whelping and routinely rejected a previously selected den for a new one, with higher final den scores. This trend was so strong that for only one of the 20 females the final den had a lower score than the one preceding it. On average, the mothers settled in dens having higher scores than the first dens as well as the average score of the intermediate dens. The pregnant females actively covered large distances in search of dens and the chances of finding better dens increased as they ventured further from the first dens. Thus it is possible for pregnant females to move away from their core territories in search of better denning sites and we have indeed observed several cases of pregnant females moving into unoccupied territories in the past (Bhadra et al., pers. obs.). Since neither the searching effort, nor the final den scores were correlated with litter size, it is most likely that the mothers searched actively for preferred dens, settling for the best available site, considering comfort in terms of available space and resources, rather than the expected size of her litter.
Canids like wolves and coyotes that live close to humans, tend to den in secluded places, away from human disturbance and respond to human disturbance at den sites by shifting their pups60,71,72,73. Interestingly, the presence of human disturbance did not affect the choice of den sites in the free-ranging dogs. On the contrary, they preferred humans as food sources over foraging sites like dustbins, i.e., they depended more on begging than scavenging41,43. Moreover, while the mothers shifted dens prior to whelping, they did not appear to shift their pups at all. The analysis of the mortality levels in the first month of pup age also shows that the mother’s choice of den site does not influence the survival of pups. Ironically, the mortality of pups increases with age, reaching 81% by the seventh month, of which 63% is human induced66.
The tendency of the pregnant females to den in the proximity of people seems to be a double edged sword. On the one hand, this provides easy access to resources (through begging), without facing competition from conspecifics, while on the other, this exposes the pups to interfering humans, which can be potentially harmful. The socio-cultural milieu of India promotes tolerance and ancient texts even recommend regular feeding of scavengers like dogs and crows74. Moreover, the laws of the land and animal rights activists ensure that the dogs lead a relatively unthreatened life on streets. Hence, the risks of denning close to humans are by far outweighed by the advantages of this behaviour. Our results show that while dogs have retained the behaviour of den selection prior to whelping, much like their ancestors, they have adapted to the human dominated environment by accepting, rather than avoiding the proximity to humans while denning. Indeed, they seem to be exploiting the tendency of humans to respond to them75 for easy access to resources. Free-ranging dogs are capable of hunting in packs like their ancestors39,76,77, the wolves, but in the urban habitat, they thrive as scavengers, living in small social groups and often choosing to forage solitarily42,52. Denning close to humans enables them to avoid competition with other mesocarnivores and predation. The tendency to den in and around human habitations and beg from humans seems to be unique to dogs and quite in contrast to other canids. In spite of the high mortality of pups induced by humans, the ability to exploit humans as sources of food and shelter appears to be a behavioural adaptation that helps dogs to survive in the urban landscape.
Methods
Den characteristics
Free-ranging dog den sites were identified through surveys42 between 2010 and 2015 by tracking dog groups and following pregnant females in various parts of West Bengal, India. In this study, we used 148 den sites in rural (5), urban (90) and semi urban (53) localities. The study was conducted in and around Kolkata (22.5667°N, 88.3667°E), Saltlake (22.5800°N, 88.4200°E), Kalyani (22.9750°N, 88.4344°E), IISER-K campus in Mohanpur (22.9638°N, 88.5246°E), Barasat (22.7200°N, 88.4800°E) and Barrackpore (22.7600°N, 88.3700°E), West Bengal, India. The localities were selected arbitrarily based on convenience and safety of sampling, in residential or business areas or both.
Observers visited the areas at random times during the day and walked on all roads and by-lanes42. Whenever a den was sighted, it was measured for length, breadth and height (if applicable) and a record was made of its physical parameters, distance from resources, nature of available resources and the size of the litter in the den (Supplementary Information 1). Each den was photographed and the date of birth of the pups was recorded with inputs from local people whenever possible. The nature of the resources available were recorded as dustbins (when dogs were seen to scavenge from dustbins and other garbage dumping sites) and people (when dogs were seen to obtain food by begging, or were actively fed by humans). We expected the dogs to prefer dens that provided greater protection from predators, had less anthropogenic disturbance and easy access to resources. Based on this, the den characteristics were given scores and thus each den obtained a total score based on its characteristics (Supplementary Information 1). For a subset of the dens (41 dens out of total 148 dens), we also obtained data on mortality of the pups up to one month of age, because beyond this stage pups do not stay inside the dens any longer.
Den selection
In order to understand the den selection procedure of expecting mothers, pregnant females were identified and followed until parturition. Each female was observed twice a day, every alternate day, from 0700–1000 h and 1500–1800 h. The study was conducted over three breeding seasons, between February 2013 and March 2015, in Kolkata, Saltlake and Barrackpore. We obtained data on 20 pregnant females, of which 15 females could be tracked for as long as a month before parturition, while the rest whelped earlier. For each pregnant female, we recorded all the above mentioned criteria (other than litter details) for the first den (D1), the intermediate dens (ID) which she was seen to occupy and the final den (DF) in which she whelped. We also recorded the date of birth, size of the litter and sex of the pups for the final dens. While following pregnant females, we recorded daily distance travelled and the corresponding route on a map, which was used to estimate the length of the path covered over the entire tracking period and calculate the linear distance between her first and final dens. We also maintained records of the time spent resting in each intermediate den. The distance between the first and final dens, the total path covered in searching and the time spent in intermediate dens were used as estimates of the searching effort of the females for locating preferred den sites. We calculated the intermediate den score (IDS) by averaging over all the den scores from the first to the last but one den occupied by the pregnant females. The difference in scores between the first and final dens was also calculated and designated as D1DF.
Statistical analysis
All statistical analyses were carried out using StatistiXL 1.10, Statistica version 12 and R statistics (R Studio)78. The dens were given scores for each of the characteristics and the total den score was computed. We used a generalized linear mixed model (GLMM) to investigate the importance of den characteristics for den site. For the GLMM analysis, litter size was considered as the response variable, with den characteristics as the fixed effects while the identity of the groups and the year of observation were taken as the random effects. Den characteristics were grouped into four categories, structure (area, height and level), resources (distance from food and water), presence of human disturbances and den quality (nature of shade and light availability) (Supplementary Information 2). Human disturbances were quantified based on the qualitative data of human (especially child) interactions with the new born pups. Many cases were observed where human child took away the pups as pet66 or interrupted the suckling bouts or sometimes beaten the pups or the mother as a part of their play. Since there were very few dens with large litters (having 7 or more pups) and mean litter size was 4, we grouped the litter sizes into two categories, small (1–4) and large (>4) and ran the GLMM analysis for a binomial distribution of litter size. In order to check if the dens selected by the mothers impact the survival of the pups (from birth to one month of pup age), we ran a Cox mixed-effects model for survivorship, considering den characteristics (structure, resources, human disturbances and quality) as fixed effects. Age of pups and survival up to one month of age (survived or not survived) were the response variables. Group identity and year of observation were added as random effects (Supplementary Information 3).
We used a Kruskal-Wallis test to compare the first den score, average intermediate den score and final den score. The three sets of den score were then compared using a Wilcoxon paired-sample test, in order to check if the females chose “better” dens while rejecting “poorer” ones. Linear regression analysis was performed to check if the number of intermediate dens used by the pregnant females depended on the time available to them before whelping.
In order to understand if the mother’s searching effort led to her finding better dens, we carried out linear regression analyses of the difference in scores between the final and first dens (D1DF) with the following: (i) the linear distance between the first and final dens; (ii) the total distance covered over the one month period of observations, considering the actual paths taken by the females and (iii) the total number of days spent in intermediate dens. Another linear regression analysis was used to check if the distance between the first and final dens increased with the total path covered in den search.
Ethical statement
No dogs were harmed during this study and all work that has been reported here was purely observation based. The methods reported in this paper were approved by the animal ethics committee of IISER Kolkata (approval number: 1385/ac/10/CPCSEA) and in accordance with approved guidelines of animal rights regulations of the Government of India.
Additional Information
How to cite this article: Majumder, S. S. et al. Denning habits of free-ranging dogs reveal preference for human proximity. Sci. Rep. 6, 32014; doi: 10.1038/srep32014 (2016).
References
Clutton-Brock, T. In The evolution of parental care Illustrated edn (Princeton University Press, New Jersey, 1991).
Gubernick, D. J. In Parental care in mammals Illustrated edn (eds. Gubernick, D. J. & Klopfer, P. H. ) 243–305 (Springer: US,, 1981).
Moehlman, P. D. Intraspecific variation in canid social systems in Carnivore behavior, ecology and evolution Illustrated edn (ed. Gittleman, J. L. ) 143–163 (Springer: US,, 1989).
Smits, C., Smith, C. & Slough, B. Physical characteristics of arctic fox (Alopex lagopus) dens in northern Yukon Territory, Canada. Arctic 41, 12–16 (1988).
Paul, M., Sen Majumder, S. & Bhadra, A. Selfish mothers? An empirical test of parent-offspring conflict over extended parental care. Behav. Processes 103, 17–22 (2014).
Ewer, R. In The carnivores illustrated reprint edn (Cornell University Press, 1973).
Poole, T. In Social behaviour in mammals (Springer: US,, 1985).
Pruss, S. Selection of natal dens by the swift fox (Vulpes velox) on the Canadian prairies. Can. J. Zool. 77, 646–652 (1999).
Arjo, W. Characteristics of current and historical kit fox (Vulpes macrotis) dens in the Great Basin Desert. Can. J. Zool. 81, 96–102 (2003).
Eide, N., Nellemann, C. & Prestrud, P. Terrain structure and selection of denning areas by arctic foxes on Svalbard. Polar Biol . 24, 132–138 (2001).
Ruggiero, L., Pearson, E. & Henry, S. Characteristics of American marten den sites in Wyoming. J. Wildl. Manage. 62, 663–673 (1998).
Hwang, Y. Local-and landscape-level den selection of striped skunks on the Canadian prairies. Can. J. Zool. 85, 33–39 (2007).
Lesmeister, D. Summer resting and den site selection by eastern spotted skunks (Spilogale putorius) in Arkansas. J. mamm . 89, 1512–1520 (2008).
Ross, S. & Kamnitzer, R. Den-site selection is critical for Pallas’s cats (Otocolobus manul). Can. J. Zool. 88, 905–913 (2010).
Norris, D. Forest composition around wolf (Canis lupus) dens in eastern Algonquin Provincial Park, Ontario. Can. J. Zool. 80, 866–872 (2002).
Henner, C. & Chamberlain, M. A multi-resolution assessment of raccoon den selection. J. Wildl. Manage. 68, 179–187 (2004).
Meriggi, A., Rosa, P., Brangi, A. & Matteucci, C. Habitat use and diet of the wolf in northern Italy. Acta Theriol. 36, 141–151 (1991).
Ye, X., Yong, Y., Yu, C. & Zhang, Z. Den selection by the giant panda in Foping Nature Reserve, China. J. Nat. Hist . 41, 2529–2536 (2007).
Szor, G., Berteaux, D. & Gauthier, G. Finding the right home: distribution of food resources and terrain characteristics influence selection of denning sites and reproductive dens in arctic foxes. Polar Biol . 31, 351–362 (2008).
Jackson, C., Power, R. & Groom, R. Heading for the Hills: Risk Avoidance Drives Den Site Selection in African Wild Dogs. PLoS One 9, e99686 (2014).
Elbroch, L., Lendrum, P., Alexander, P. & Quigley, H. Cougar den site selection in the Southern Yellowstone Ecosystem. Mammal Res . 60, 89–96 (2015).
Chapman, R. In The effects of human disturbance on wolves (Canis lupus L.) 1–154 (University of Alaska, Biology and Resource Management Project, Alaska Cooperative Park Studies Unit, 1976).
Clark, K. In Food habits and behavior of the tundra wolf on central Baffin Island 1–223 (PhD thesis, University of Toronto, 1971).
Haber, G. In Socio-ecological dynamics of wolves and prey in a subarctic ecosystem (Joint Federal-State Land Use Planning Commission for Alaska, 1977).
Jordan, P., Shelton, P. & Allen, D. Numbers, turnover and social structure of the Isle Royale wolf population. Am. Zool . 7, 233–252 (1967).
Lawhead, B. In Wolf den site characteristics in the Nelchina Basin, Alaska 1–65 (University of Alaska, 1983).
Woodroffe, R. Predators and people: using human densities to interpret declines of large carnivores. Anim. Conserv. 3, 165–173 (2000).
Inskip, C. & Zimmermann, A. Human-felid conflict: a review of patterns and priorities worldwide. Oryx 43, 18–34 (2009).
Herr, J., Schley, L., Engel, E. & Roper, T. Den preferences and denning behaviour in urban stone martens (Martes foina). Mamm. Biol. für 75, 138–145 (2010).
Gould, N. & Andelt, W. Reproduction and denning by urban and rural San Clemente Island foxes (Urocyon littoralis clementae). Can. J. Zool. 89, 976–984 (2011).
Theuerkauf, J. & Jȩdrzejewski, W. Accuracy of radiotelemetry to estimate wolf activity and locations. J. Wildl. Manage. 66, 859–864 (2002).
Palomares, F. & Caro, T. Interspecific killing among mammalian carnivores. Am. Nat. 153, 492–508 (1999).
Benson, J., Patterson, B. & Mahoney, P. A protected area influences genotype-specific survival and the structure of a Canis hybrid zone. Ecology 95, 254–264 (2014).
Kruuk, H. & Snell, H. Prey selection by feral dogs from a population of marine iguanas (Amblyrhynchus cristatus). J. Appl. Ecol. 18, 197 (1981).
Daniels, T. & Bekoff, M. Population and social biology of free-ranging dogs, Canis familiaris. J. Mammal. 70, 754–762 (1989).
De Balogh, K. K., Wandeler, A. I. & Meslin, F. X. A dog ecology study in an urban and a semi-rural area of Zambia. Onderstepoort J. Vet. Res . 60, 437–443 (1993).
Pal, S. K. Population ecology of free-ranging urban dogs in West Bengal, India. Acta Theriol. 46, 69–78 (2001).
Kato, M., Inukai, Y., Yamamoto, H. & Kira, S. Survey of the stray dog population and the health education program on the prevention of dog bites and dog acquired infections: a comparative study in Nepal and Okayama prefecture, Japan. Acta Med. Okayama 57, 1–6 (2003).
Butler, J. Free-ranging domestic dogs (Canis familiaris) as predators and prey in rural Zimbabwe: threats of competition and disease to large wild carnivores. Biol. Conserv. 115, 369–378 (2004).
Ortega-Pacheco, A. et al. A survey of dog populations in Urban and rural areas of Yucatan, Mexico. Anthrozoos 20, 261–274 (2007).
Vanak, A. T. & Gompper, M. E. Dogs canis familiaris as carnivores: Their role and function in intraguild competition. Mamm. Rev . 39, 265–283 (2009).
Sen Majumder, S. et al. To be or not to be social: foraging associations of free-ranging dogs in an urban ecosystem. Acta Ethol . 17, 1–8 (2014).
Bhadra, A. & Bhadra, A. Preference for meat is not innate in dogs. J. Ethol. 32, 15–22 (2013).
Mech, L. Wolf population survival in an area of high road density. Am. Midl. Nat . 121, 387–389 (1989).
Thiel, R., Merrill, S. & Mech, L. Tolerance by denning wolves, Canis lupus, to human disturbance. Can. F. Nat . 112, 340–342 (1998).
Vanak, A. T. & Gompper, M. E. Interference competition at the landscape level: The effect of free-ranging dogs on a native mesocarnivore. J. Appl. Ecol . 47, 1225–1232 (2010).
Wandeler, A. I., Matter, H. C., Kappeler, A. & Budde, A. The ecology of dogs and canine rabies: a selective review. Rev. Sci. Tech. 12, 51–71 (1993).
Leonard, J. a. et al. Ancient DNA evidence for Old World origin of New World dogs. Science 298, 1613–1616 (2002).
Scott, J. & Fuller, J. In Genetics and the social behavior of the dog Illustrated reprint edn 1–468 (University of Chicago Press, 1965).
Serpell, J. In The domestic dog: its evolution, behaviour and interactions with people Illustrated reprint edn . 1–268 (Cambridge University Press, Cambridge, 1995).
Ghosh, B., Choudhuri, D. & Pal, B. Some aspects of the sexual behaviour of stray dogs, Canis familiaris. Appl. Anim. Behav. Sci. (1984).
Pal, S., Ghosh, B. & Roy, S. Dispersal behaviour of free-ranging dogs (Canis familiaris) in relation to age, sex, season and dispersal distance. Appl. Anim. Behav. Sci. 61, 123–132 (1998).
Vanak, A. T., Thaker, M. & Gompper, M. E. Experimental examination of behavioural interactions between free-ranging wild and domestic canids. Behav. Ecol. Sociobiol. 64, 279–287 (2009).
Beaver, B. V. In Canine behavior: insights and answers 2nd edn 1–315 (Saunders, Elsevier, 2009).
Chapman, R. Human disturbance at wolf dens—a management problem. 1st Conference on Scientific Research in the National Parks. New Orleans, Louisiana (1976).
Ahmadi, M., Kaboli, M. & Nourani, E. A predictive spatial model for gray wolf (Canis lupus) denning sites in a human-dominated landscape in western Iran. Ecological Research 28, 513–521 (2013).
Mladenoff, D., Sickley, T. & Wydeven, A. Predicting gray wolf landscape recolonization: logistic regression models vs. new field data. Ecol. Appl. 9, 37–44 (1999).
Karlsson, J., Brøseth, H., Sand, H. & Andrén, H. Predicting occurrence of wolf territories in Scandinavia. J. Zool. 272, 276–283 (2007).
Jędrzejewski, W. Habitat suitability model for Polish wolves based on long‐term national census. Animal Conservation 11, 377–390 (2008).
Way, J., Auger, P., Ortega, I. & Strauss, E. Eastern coyote denning behavior in an anthropogenic environment.Northeast Wildl. 56, 18–30 (2001).
Gosselink, T. In Seasonal variations in habitat use and home range of sympatric coyotes and red foxes in agricultural and urban areas of east-central Illinois 1–142 (University of Illinois at Urbana-Champaign, 1999).
Marks, C. & Bloomfield, T. Home-range size and selection of natal den and diurnal shelter sites by urban red foxes (Vulpes vulpes) in Melbourne. Wildl. Res. 33, 339–347 (2006).
Home, C. & Jhala, Y. Estimating breeding pair densities of the Indian fox in Kutch, Gujarat, India. Canid News 13, 1–6 (2010).
Spotte, S. In Societies of wolves and free-ranging dogs Illustrated edn (Cambridge University Press, 2012).
Pal, S. Parental care in free-ranging dogs, Canis familiaris. Appl. Anim. Behav. Sci. 90, 31–47 (2005).
Paul, M., Sen, S., Sau, S., Nandi, A. K. & Bhadra, A. High early life mortality in free- ranging dogs is largely influenced by humans. Sci. Rep. 6, 19641 (2016).
Person, D. & Russell, A. Reproduction and den site selection by wolves in a disturbed landscape. Northwest Sci . 83, 211–224 (2009).
Kaartinen, S., Luoto, M. & Kojola, I. Selection of den sites by wolves in boreal forests in Finland. J. Zool . 281, 99–104 (2010).
Habib, B. & Kumar, S. Den shifting by wolves in semi‐wild landscapes in the Deccan Plateau, Maharashtra, India. J. Zool. 272, 259–265 (2007).
Mech, L. The wolf: ecology and behavior of an endangered species. British journal of hospital medicine (London, England : 2005) 74, (Natural History Press, Garden City, 1970).
Harrison, D. & Gilbert, J. Denning ecology and movements of coyotes in Maine during pup rearing. J. Mammal. 66, 712–719 (1985).
Argue, A. Behavioural response of eastern wolves (Canis lycaon) to disturbance at homesites and its effects on pup survival. Can. J. Zool. 86, 400–406 (2008).
Nonaka, Y. Response of breeding wolves to human disturbance on den sites–an experiment. Candidate thesis, Biology Education Centre, Uppsala University, Sweden and Grimso Wildlife Research station, University of Agriculture, Uppsala, Sweden (2011).
Debroy, B. In Sarama and her children: the dog in Indian myth . (Penguin Books, India, 2008).
Nagasawa, M., Mitsui, S., En, S. & Ohtani, N. Oxytocin-gaze positive loop and the coevolution of human-dog bonds. Science 348, 333–336 (2015).
Vanak, A. T. & Gompper, M. E. Dietary niche separation between sympatric free-ranging domestic dogs and Indian foxes in central India. J. Mammal. 90, 1058–1065 (2009).
Butler, J. R. a. & du Toit, J. T. Diet of free-ranging domestic dogs (Canis familiaris) in rural Zimbabwe: implications for wild scavengers on the periphery of wildlife reserves. Anim. Conserv. 5, 29–37 (2002).
R Core Team R: A language and environment for statistical computing . R Foundation for Statistical Computing, Vienna, Austria, (2013).
Acknowledgements
The authors would like to thank Dr. Anjan K. Nandi of the Department of Physical Sciences, IISER Kolkata for his critical inputs on the GLMM analysis. This work was partially funded by projects from Council for Scientific and Industrial Research, India and SERB, Department of Science and Technology; and supported by IISER Kolkata, India.
Author information
Authors and Affiliations
Contributions
The data for this paper was collected by S.S.M., M.P. and S.S. Statistical analyses were carried out by M.P., S.S.M. and A.B. A.B. supervised the work and co-wrote the paper with M.P.
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Electronic supplementary material
Rights and permissions
This work is licensed under a Creative Commons Attribution 4.0 International License. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in the credit line; if the material is not included under the Creative Commons license, users will need to obtain permission from the license holder to reproduce the material. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/
About this article

Cite this article
Sen Majumder, S., Paul, M., Sau, S. et al. Denning habits of free-ranging dogs reveal preference for human proximity. Sci Rep 6, 32014 (2016). https://doi.org/10.1038/srep32014
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/srep32014
This article is cited by
-
Time-activity budget of urban-adapted free-ranging dogs
acta ethologica (2022)
-
Adjustment in the point-following behaviour of free-ranging dogs – roles of social petting and informative-deceptive nature of cues
Animal Cognition (2022)
-
Response to short-lived human overcrowding by free-ranging dogs
Behavioral Ecology and Sociobiology (2021)
-
Dogs and wolves do not differ in their inhibitory control abilities in a non-social test battery
Animal Cognition (2019)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
