
Abstract
Recently, 5-hydroxymethylcytosine patterning across the tumor genome was considered as a hallmark of cancer development and progression. However, locus-specific difference of hydroxymethylation between colorectal cancer and normal tissue is unknown. In this study, we performed a newly developed method, HMST-seq, to profile 726 aberrant methylated loci and 689 aberrant hydroxymethylated loci synchronously in genome wide of colorectal cancers, majority of which presented higher methylation or lower hydroxymethylationin than in normal group. Besides, abnormal hydroxymethylated modification was more frequently occur at proximal regions close to TSSs and TSSs regions than abnormal methylation. Subsequently, we screened four genes (ALOX15, GHRHR, TFPI2 and TKTL1) with aberrant methylation and aberrant hydroxymethylation at some genome position by functional enrichment analysis as candidate genes associated with colorectal cancer. Our results may allow us to select differentially epigenetically modified target genes implicated in colorectal cancer tumorigenesis.
Similar content being viewed by others

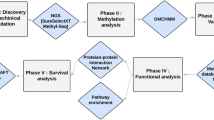

Introduction
DNA methylation is an important epigenetic modification for gene transcription and an amount of evidence has been shown that genes with high levels of 5-methylcytosine (5mC) in their promoter region are transcriptionally silent, subsequently, DNA methylation gradually accumulates resulting in long-term gene silencing. Aberrant DNA methylation patterns – hyper-methylation and hypo-methylation differ from normal tissue – have been discovered in many kinds of human cancers1,2. Hyper-methylation typically occurs at CpG islands in the promoter region and is associated with gene inactivation. Global hypo-methylation has also been implicated in the progression of cancer through different mechanisms. Typically, there is hyper-methylation of tumor suppressor genes (TSGs) and hypomethylation of oncogenes.
5-mC can be oxidized to 5-hydroxymethylcytosine (5hmC), then, can be further oxidized to 5-formylcytosine (5fC) and 5- carboxylcytosine (5caC) by the ten-eleven translocation (TET) enzyme family proteins, which may play a role in DNA demethylation3,4. 5hmC has been found significantly reduced in malignant cells, including colorectal cancer (CRC)5. Since conventional technologies, such as bisulfite sequencing and restriction enzyme cutting technology, could not distinguish between 5-mC and 5-hmC6,7 and the existence of 5-hmC in samples could influence the accuracy of DNA methylation detection8. Thus, there is an urgent requirement of a new technology to detect 5-mC and 5-hmC simultaneously in malignant tumor genomic profile research for higher accuracy of detection of aberrant DNA methylation patterns compare to normal tissue.
In this research, we used a newly developed single-base high-throughput sequencing approach (hydroxymethylation and methylation sensitive tag sequencing (HMST-seq)) to globally detect distribution of both 5mC and 5hmC at CCGG sites synchronously, in CRC samples and their adjacent non-cancerous normal tissues. Subsequently, we performed functional analysis of genes containing differentially hydroxymethylated or methylated regions (DhMR or DMR) by WEB-based GEne SeT AnaLysis Toolkit (WebGestalt) to screen genes associated with colorectal cancer. As a result, four genes, including ALOX15, GHRHR, TFPI2 and TKTL1, were revealed as key CRC-associated candidate genes with aberrant methylation and aberrant hydroxymethylation at same promoter position. Significant hypermethylation as well as hypo-hydroxymethylation of promoters were revealed for these genes in CRC samples. In conclusion, based on novel HMST-seq technology, our study identified new candidate genes that were epigenetically modulated in CRC, which can be applied as clinical biomarkers or candidates of tumor suppressor genes in future studies.
Results
Distribution of 5mC and 5hmC in CRC samples and matching normal tissues
To investigate the distribution of 5mC and 5hmC in CRC specimens and matching normal samples, a new improved technology ‘hydroxymethylation and methylation sensitive tag sequencing (HMST-seq)’ was performed on the 16 pairs of CRCs and accordingly normal samples. 5mC and 5hmC can be investigated using HMST-seq at ‘CCGG’ sites with a single-base resolution and three libraries were established for every sample, including ‘C + mC + hmC’ library represented to contain unmodified C, mC and hmC tags through MspI-mediated digestion of genomic DNA, ‘C + mC’ library represented to contain C and mC tags through MspI-mediated digestion of glucosylated genomic DNA and ‘C’ library represented to contain C tags only through HpaII-mediated digestion of normal genomic DNA9. GRSN method was performed to normalize the data for the three libraries.
On average, approximately 342 million reads for each library were generated and about 110.7 million (32.34%) reads were aligned to a virtual library of the human genome (hg19). An average of 57.23% uniquely aligned reads (0 mismatch + 1 mismatch) and 25.59% multiple aligned reads (0 mismatch + 1 mismatch) were then obtained for each library, resulting in an average of 1.346 million total cytosine with greater than 30 X sequencing depth for each library (see Supplementary Table S1). Based on our method, the hydroxymethylation level can be defined as the ratio between ‘C + mC + hmC’ and ‘C + mC’ tags, while methylation abundance can be represented by ratio between tag counts of “C + mC” and “C” libraries. We then examined the genome-wide distribution of methylation and hydroxymethylation levels of each detectable CCGG site. The majority of the CCGG sites displayed high level of methylation. In contrast, majority of the CCGG sites displayed low level of hydroxymethylation (Fig. 1). Thereby, on average, 1,162,336 (82,59%) and 106,907 (7.58%) total examined CCGG sites were identified as significant for 5mC and 5hmC, respectively, in matching normal specimens, while 5mC (1,105,904; 78.38%) and 5hmC (75,236; 5.33%) modification on informative sites were observed in the CRC specimens (see Supplementary Table S2) (sequencing depth >10 X, FDR < 0.001)9.

Boxplot showing distribution of methylation and hydroxymethylation levels of each detectable CCGG sites in genome wide of CRCs (red) and matched normal sample (bule).
The hydroxymethylation level is defined as the ratio between ‘C + mC + hmC’ and ‘C + mC’ tags, while methylation abundance is defined as the ratio between tag counts of “C + mC” and “C” libraries. T1 is a sign of patient 1’s tumor sample, while N1 is a sign of patient 1’s normal sample.
DMRs and DhMRs were revealed by comparison of CRC and matching normal samples
To identify differentially methylated regions (DMRs) and differentially hydroxymethylated regions (DhMRs) between CRC and matching normal samples according to the data of global methylation and hydroxymethylation levels of each detectable CCGG site in 16 pairs of CRC and non-CRC samples, a sliding window strategy was performed as our previous study9. Take the first 5 CCGG sites that contains at least 4 CCGG sites with the same changing trend between tumor and normal group and Wilcoxon rank sum test P-value < 0.05 as the seed sites of a candidate DMR or DhMR. We found 726 DMRs (see Supplementary Table S3) and 689 DhMRs (see Supplementary Table S4) between CRCs group and normal tissue group, interestingly, we discovered that DhMRs were more likely to occur at proximal regions close to TSSs and TSSs regions (20.17%) rather than DMRs (13.22%), although most part of DMRs (83.88%) and DhMRs (77.51%) distributing in intergenic and gene body (Fig. 2).

The distribution of DMRs and DhMRs.
(a) The distribution ratio of DMRs and DhMRs within genomic regions. The DhMR or DMR located in two different elements are attributed to the element that overlaps with a bigger proportion of the DhMR or DMR and counted only once. (b) The distance of DMRs and DhMRs to transcriptional start sites (TSSs).
635 out of 726 DMRs (87.47%) were hypermethylated, while 551 out of 689 DhMRs (79.97%) were hypo-hydroxymethylated in CRC group (Fig. 3). Considering the facts that 5hmC is an intermediate during in TET-associated DNA demethylation, which is one of proposed mechanisms of DNA demethylation10, lower level of hydroxymethylation suggested for less dynamic preprogramming rate for specific genomic region. All together, the high percentage of hypermethylated DMRs as well as hypo-hydroxymethylated DhMRs both indicated that majority of detected CCGG sites by HMST-seq were stably hypermethylatd in CRC group comparing with normal controls. Nevertheless, only 114 genes were detected containing both DMR and DhMR, among which only 38 genes were with DMR and DhMR at the same location, at least partly supporting the role of 5-hmC as an inter-mediate in DNA demethylation. Among 38 genes with DMR and DhMR at same genome position, 35 genes were detected hyper-methylation and hypo-hydroxymethylation in CRC group and 11 out of these 35 genes containing DMR and DhMR at promoter region.

Range of methylation and hydroxymethylation level in DMRs and DhMRs.
The vertical axis shows the abundance of methylation (left) and hydroxymethylation (right) in DMRs or DhMRs. Hyper-DMR (n = 635), loci with higher methylation in CRC group; Hypo-DMR (n = 91), loci with lower methylation in CRC group; Hyper-DhMR (n = 138), loci with higher hydroxymethylation in CRC group; Hypo-DhMR (n = 551), loci with lower hydroxymethylated modification in CRC group; all of these DMRs and DhMRs are confirmed by statistics analysis comparing with normal tissue.
CRC-associated candidate genes with aberrant DNA modifications
Functional enrichment analysis for genes with DMRs or DhMRs between CRC and matching normal samples was performed using WebGestalt. We found that DMR and DhMR genes were enriched in ‘Pathways in cancer’, ‘MAPK signaling pathway’, ‘Endocytosis’ and ‘Regulation of actin cytoskeleton’ in KEGG pathways, but these genes rarely overlapped. Besides, cancer-related pathways including, ‘Focal adhesion’, ‘Neuroactive ligand-receptor interaction’ and ‘Metabolic pathways’ were enriched for DhMR genes (see Supplementary Table S5 and Fig. 4). According to the disease enrichment analysis, we found that DhMR genes were notably enriched in epithelial carcinomas or gastrointestinal neoplasms such as, ‘Gastrointestinal Neoplasms’, ‘epithelial cancers’, ‘Carcinoma’, ‘Carcinoma in Situ’ and ‘Adenocarcinoma’, while only 11 DMR genes were detected associated with ‘Gastrointestinal Diseases’ (see Supplementary Table S6 and Fig. 4).

KEGG pathway and diseases enrichment analysis of genes related to DMRs or DhMRs.
(a) KEGG pathway enrichment analysis of DMRs, (b) diseases enrichment analysis of DMRs, (c) KEGG pathway enrichment analysis of DhMRs, (d) diseases enrichment analysis of DhMRs. The x-axis shows the P value from hypergeometric test adjusted by the multiple test adjustment.
We then selected 53 DMR genes that significantly enriched in 10 disease categories and 2 KEGG categories (‘Pathways in cancer’ and ‘MAPK signaling pathway’), as well as 56 DhMR genes that significantly enriched in 10 disease categories and 3 KEGG categories (“Focal adhesion”, “Pathways in cancer” and “MAPK signaling pathway”) as candidate genes associated with colorectal cancer.
Among these 103 candidate genes, we found 6 genes were common in both DMR and DhMR data, including ALOX15, CCNE1, FGFR2, GHRHR, TFPI2 and TKTL1. Among these 6 genes, ALOX15, GHRHR, TFPI2 and TKTL1 contained both DMR and DhMR in the same genomic position. We then examined the modification status of these genes. Opposite tendency of methylation and hydroxymethylation changes was revealed for all these genes, that is, all genes were significantly hypermethylated as well as hypo-hydroxymethylated in CRC samples. As these DMRs or DhMRs were all distributed in promoter or TSS regions, these genes might be suppressored for gene expression in CRC.
Discussion
Hyper-methylation is considered as the principle mechanism of dysfunction of tumor suppressor genes in cancer progression, moreover, promoter hyper-methylation status of some tumor suppressor genes in colon adenocarcinoma were confirmed by previous studies11,12, while hydroxymethylation has been reported as an epigenetic marker whose decrease is broadly and tightly associated with cancer development13,14,15. Since 5-hydroxymethylation in cancer research is still in infancy, further investigation is needed. Nevertheless, some conventional technologies cannot distinguish 5hmC from 5mC, such as bisulfite treatment or methylation-sensitive enzyme digestion-based methods6,7. Affinity enrichment assays are bulk assays and are unable to resolve 5mC with nucleotide precision16. Therefore, difference technologies might lead to conflicting results among close research studies and impact inaccurate quantification of DNA methylation levels. Thus, a sensitive and specific technology is required to distinguishing between 5hmC and 5mC in the identification of DNA methylation.
In this study, we applied the newly developed HMST-Seq technology, which was confirmed as a cost-effective and selective detection method of epigenetic modification9,17, to profile DNA methylation and hydroxymethylation levels of CRC samples and matching normal tissue synchronously in genome wide. According to our data, 106,907 (7.58%) and 75,236 (5.33%) of the total CCGG sites were called out as 5hmC sites in control and CRC samples, respectively. If we used bisulfite-treatment based methods, but not HMST-seq, all of these 5hmC sites will be taken as 5mC, since conventional bisulfite-treatment based methods can’t differentiate 5hmC from 5mC6,7. This result clearly indicated for the increased sensitivity of HMST-seq in examining DNA modifications. Regarding to epigenetic changes of CRC genomes, our results showed 726 DMRs and 689 DhMRs between CRCs and normal samples and most of they presented hyper-methylation or lower level of hydroxymethylation in CRC samples, indicating majority of detected CCGG sites by HMST-seq were stably hypermethylatd in CRCs comparing with normal controls. Among 38 genes with DMRs and DhMRs at same locus, 35 genes were detected with hyper-methylation and hypo-hydroxymethylation in CRC group and 11 out of these 35 genes containing DMR and DhMR at promoter region, suggesting that reducing of 5hmC and increasing of 5mC in these 35 genes were observed at same genome position in CRCs comparing with normal tissue, moreover promoter was the most frequent location for observing changes of 5mC and 5hmC level in these loci. 5hmC was reported as a strong inhibitor of the DNA methyltransferase maintenance methylation reaction catalyzed by DNMT118,19, leading to passive DNA demethylation over subsequent replication cycles. Thus, depletion of 5hmC of specific locus in CRC probably resulted in reduction of DNA methyltransferase inhibition, generating hyper-methylation at this locus. Thus, we inferred that low level of hydroxymethylation of promoters might mark or even facilitate DNA hypermethylation at same loci of CRCs by recovering activity of DNA methyltransferase. However, DNA methylation-demethylation is a complex story and global changes in DNA methylation are thus likely to require a concerted action of several DNA demethylation mechanisms, thus, TET-associated DNA demethylation is not the only proposed mechanism of DNA demethylation, other factor could also lead to DNA demethylation (eg. the activity of Dnmt1 DNA methyltransferase)10. Therefore, changes of 5mC in locus could not alone account for changes of TET-catalyzed 5-hydroxymethylcytosine level. In our data, the rest observed loci without identical location with both DMR and DhMR might be explained by other DNA demethylation mechanisms. But the detail needed to be further studied.
In present research, we discovered that DhMRs were more likely to occur at proximal regions close to TSSs and TSSs regions (20.17%) rather than DMRs (13.22%), illustrating that changes in 5hmC of CRCs probably play an important role in gene transcription and expression. Recently, 5hmC was considered as an important factor in transcript process20,21, most importantly, gene-specific 5-hmC changes in human colon cancers are directly correlated with changes in gene expression22, thus, finding gene-specific 5hmC changes in CRC seems very important to screen different expression of genes in CRC and normal tissue.
Based on KEGG data, we discovered 53 DMR genes and 56 DhMR genes that enriched multiple highly interrelated cancer signaling pathways, including ‘Focal adhesion’, ‘Pathways in cancer’ and ‘MAPK signaling pathway’ as well as some diseases, such as ‘Gastrointestinal Diseases’ and ‘Gastrointestinal Neoplasms’, these 103 were genes selected as candidate genes associated with colorectal cancer. Among these 103 candidate genes, the identical locus of DMR and DhMR was discovered in ALOX15, GHRHR, TFPI2 and TKTL1 genes.
15-Lipoxygenase-1 (15-LOX-1, ALOX15) is an inducible and highly regulated enzyme in normal human cells23 that plays an important role in the production of lipid signaling mediators24. Some researches have proved down-regulation of ALOX15 in various human cancers (e.g., cancers of the colon25,26,27, breast28, lung29 and pancreas30, supporting the theory that ALOX15 is a tumor suppressor gene in these cancers. Zou et al. found that the ALOX15 promoter was methylated in colorectal cancer cells in vitro and in 36% (18/50) of colorectal cancer patients but virtually absent in the normal colons without history of colorectal cancer or polyps31. Consistently, in this study, ALOX15 was observed with hyper-methylation and hypo-hydroxymethylation at promoter in CRC samples comparing with matching normal tissue. However, Zou elucidated that ALOX15 promoter DNA methylation level had no significant correlation with its expression and promoter demethylation failed to reestablish ALOX15 expression31. These data indicated that promoter methylation was not the only reason for suppression of ALOX15, otherwise involve multiple layers of repression mechanisms. Since presence of 5hmC in promoter would impact the level of transcription, our characterization of lower hydroxymethylated level in promoter of CRCs, provides a possible reason of down-regulation of ALOX15 in colon cancer. The validate work need to be done in the future study.
Growth-hormone-releasing hormone receptor (GHRHR) is dominantly expressed in the pituitary but it is also found in various cancer cell lines and tumors32. Combination of GHRH and GHRHR activates the mitogen-activated protein kinases pathway (MAPKs), which is involved in cell proliferation and differentiation33. Hohla et al. discovered strong expression of GHRHR in tubulovillous adenomas and colorectal cancers and absence in normal colonic mucosa34. In our present research, GHRHR promoter was detected hyper-methylated and hypo-hydroxymethylated in CRC samples. Nevertheless, we cannot exclude other mechanism of boost expression of GHRHR. Further studies on histone modification or transcriptional activator are needed to explain why hyper-methylation of GHRHR promoter presents strong protein expression.
Tissue factor pathway inhibitor 2 (TFPI2), a Kunitz-type serine proteinase inhibitor, The protein can inhibit a variety of serine proteases including factor VIIa/tissue factor, factor Xa, plasmin, trypsin, chymotryspin and plasma kallikrein. Hibi et al. detected TFPI2 methylation in the serum of colorectal cancer patients35, while after curative surgery methylation of TFPI2 no longer detected in the serum DNA, indicating that methylated TFPI2 in serum DNA was derived from colorectal cancer36. Furthermore, he found methylation of TFPI2 is frequently observed in colorectal cancer, suggesting TFPI2 may act as a tumor suppressor in colorectal carcinomas37. However, no research was involved in analysis of hydroxymethylated level of TFPI2 and location of hydroxymethylation modification. Our data showed higher methylation and lower hydroxymethylation levels of TFPI2 at TSS in CRC samples, providing the specific locus with aberrant hydroxymethylated level, which could be another reason of impacting transcription of TFPI2.
Transketolase-like 1 (TKTL1) is a transketolase that acts as a homodimer and catalyzes the conversion of sedoheptulose 7-phosphate and D-glyceraldehyde 3-phosphate to D-ribose 5-phosphate and D-xylulose 5-phosphate. This reaction links the pentose phosphate pathway with the glycolytic pathway. Previous studies have proven that TKTL1 was required for rapid cell growth and proliferation of human colon cancer cells38,39. Interestingly, Diaz-Moralli et al. discovered that a significant association of TKTL1 expression with lymph-node involvement while a significant decrease of TKTL1 expression associated with metastasis40, similar appearance has been observed in mutation of ras in CRC: the incidence of mutation increases from 7% to 60% starting early till late carcinomas, but decreases to 45% in invasive phrase, indicating an possibility that TKTL1 contributed in cell aggressiveness but not in metastasized cells like ras mutation41,42. Besides, Smith et al. proposed that TKTL1 could be up-regulated in response to hypo-methylation in head and neck cancer and lung cancer43. In this research, we detected hypo-methylation and hyper-hydroxymethylation at TSS of TKTL1 in CRC, provided another suggestion that increasing expression of TKTL1 in aggressive CRC probably due to not only hypo-methylation but also hyper-hydroxymethylation at TSS.
In summary, our study highlighted genes with aberrant methylation and hydroxymethylation in CRCs by HMST-seq, subsequently, enrichment analysis of these genes was performed using WebGestalt and 103 genes were identified as differentially epigenetically modified target genes implicated in colorectal cancer tumorigenesis. Among them, ALOX15, GHRHR, TFPI2 and TKTL1 were observed the identical locus of DMR and DhMR. However, expression and functions of select differentially epigenetically modified target genes implicated in colorectal cancer tumorigenesis were needed to validate in further research base on our DMR and DhMR data.
Materials and Methods
Patient samples
This study was conducted under the principles of the World Medical Association Helsinki agreement. Ethical approval was obtained from the Chongqing Medical University Ethics Committee. Informed consent was obtained for experimentation with human subjects. All the 16 pairs of specimens (tumors and relevant adjacent tissues) were obtained from patients who signed informed consent before surgery (see Supplementary Table 7S) and immediately frozen at −80 °C in liquid nitrogen container until DNA extraction. The CRC diagnosis was confirmed through pathological examination.
DNA extraction
Genomic DNA was extracted from samples according to the instruction of DNeasy Blood and Tissue kit (Qiagen, Valencia, CA, USA), provided by the manufacturer.
Data analysis by hydroxymethylation and methylation sensitive tag sequencing (HMST-Seq)
Genomic DNA, extracted by DNeasy Blood and Tissue kit, was detected by HMST-Seq and the libraries were constructed on the basis of our previous study9. Three independent libraries with initially different enzyme digestion were constructed for each sample, which were termed “C” library (c), “C + mC” library (a) and “C + mC + hmC” (b), respectively. For “C + mC” library construction, an aliquot of genomic DNA from each sample was first glucosylated by incubating DNA substrates with T4 β-glucosyltransferase (β-GT) (NEB). After glucosylation, the aliquot of glucosylated DNA was then digested with MspI (NEB). For the “C” or “C + mC + hmC” libraries, the aliquots of DNA without glucosylation were directly digested with HpaII or MspI (NEB), respectively. After digestion, the digested aliquots of DNA from all three libraries were ligated with biotinylated linker, fragmented by NlaIII, captured by streptavidin-conjugated beads, digested with MmeI to generate short sequence tags (16–17 bp) and ligated with sequencing linkers and amplified by PCR. Then, the purified tags were sequenced using Illumina HiSeq analyzer according to the manufacturer’s instructions.
Sequencing data will be mapped to reference genome. Only the uniquely mapped reads can be used for standard analysis and personal bioinformatics analysis.
Data Filtering and Alignment
We extracted the 16–17 bp tags from sequencing reads after base calling, adapter removal and low-quality reads filtering. Then the virtual library was constructed as follows: The human genome sequence (hg19) was in silico digested by MspI or NlaIII, we defined the DNA sequences between the nearest NlaIII sites around each MspI site in both directions as the virtual library reference for mapping. Using the open-source programming language Perl, all the tags were mapped to the virtual library with no more than one mismatch and the unambiguous mapped tags were used for further analysis.
The data normalization among libraries was performed according to Global Rank-invariant Set Normalization (GRSN) method44 and the hydroxymethylation or methylation abundance of CCGG site was determined as the ratio between tag counts of two different libraries. Thus, the hydroxymethylation level can be defined as the ratio between ‘C + mC + hmC’ and ‘C + mC’ tags, while methylation abundance can be represented by ratio between tag counts of “C + mC” and “C” libraries. Moreover, CCGG sites containing significantly different tag counts (sequencing depth >10X, FDR < 0.001) based on Poisson distribution, simultaneously, a ratio of tags between two libraries larger than 1 were defined to be significantly modified sites.
Differentially methylated or hydroxymethylated regions
Differentially hydroxymethylated or methylated regions were defined in the following steps: a) take the first 5 CCGG sites that contains at least 4 CCGG sites with the same changing trend between tumor and normal group and Wilcoxon rank sum test P-value < 0.05 as the seed sites of a candidate DMR; b) then a 3′ downstream adjacent CCGG with the same changing trend was incorporated with this candidate DMR. Up to 2000 bp inter-distance was allowed between the two adjacent CCGGs, Wilcoxon rank sum test was performed in the incorporated region; c) repeat these steps until Wilcoxon rank sum test P-value >= 0.05; d) the incorporated region were defined as a DhMR or DMR.
Enrichment analysis of DMRs and DhMRs
Functional enrichment analysis for genes with DMRs or DhMRs was performed using WebGestalt45,46, which is freely accessible at http://www.webgestalt.org.
Additional Information
How to cite this article: Li, M. et al. Filtrating colorectal cancer associated genes by integrated analyses of global DNA methylation and hydroxymethylation in cancer and normal tissue. Sci. Rep. 6, 31826; doi: 10.1038/srep31826 (2016).
References
Baylin, S. B. & Jones, P. A. A decade of exploring the cancer epigenome - biological and translational implications. Nat Rev Cancer 11, 726–734 (2011).
Timp, W. & Feinberg, A. P. Cancer as a dysregulated epigenome allowing cellular growth advantage at the expense of the host. Nat Rev Cancer 13, 497–510 (2013).
Tahiliani, M. et al. Conversion of 5-methylcytosine to 5-hydroxymethylcytosine in mammalian DNA by MLL partner TET1. Science 324, 930–935 (2009).
Ito, S. et al. Tet proteins can convert 5-methylcytosine to 5-formylcytosine and 5-carboxylcytosine. Science 333, 1300–1303 (2011).
Kudo, Y. et al. Loss of 5-hydroxymethylcytosine is accompanied with malignant cellular transformation. Cancer Sci. 103, 670–676 (2012).
Huang, Y. et al. The behaviour of 5-hydroxymethylcytosine in bisulfite sequencing. PLoS One 5, e8888 (2010).
Jin, S. G., Kadam, S. & Pfeifer, G. P. Examination of the specificity of DNA methylation profiling techniques towards 5-methylcytosine and 5-hydroxymethylcytosine. Nucleic Acids Res 38, e125 (2010).
Gao, F. & Xia, Y. Hydroxymethylation- and methylation-sensitive tag sequencing: how will this technology change clinical applications of DNA methylation profiling? Epigenomics 5, 355–357 (2013).
Gao, F. et al. Integrated detection of both 5-mC and 5-hmC by high-throughput tag sequencing technology highlights methylation reprogramming of bivalent genes during cellular differentiation. Epigenetics 8, 421–430 (2013).
Hill, P. W., Amouroux, R. & Hajkova, P. DNA demethylation, Tet proteins and 5-hydroxymethylcytosine in epigenetic reprogramming: an emerging complex story. Genomics 104, 324–33 (2014).
Michailidi, C. et al. Expression and promoter methylation status of hMLH1, MGMT, APC and CDH1 genes in patients with colon adenocarcinoma. Exp Biol Med (Maywood) 240, 12 (2015).
Mitchell, S. M. et al. A panel of genes methylated with high frequency in colorectal cancer. BMC Cancer 14, 54 (2014).
Yang, H. et al. Tumor development is associated with decrease of TET gene expression and 5-methylcytosine hydroxylation. Oncogene 32, 663–669 (2013).
Kroeze, L. I., van der Reijden, B. A. & Jansen, J. H. 5-Hydroxymethylcytosine: An epigenetic mark frequently deregulated in cancer. Biochim Biophys Acta 1855, 144–154 (2015).
Rodger, E. J., Chatterjee, A. & Morison, I. M. 5-hydroxymethylcytosine: a potential therapeutic target in cancer. Epigenomics 6, 503–514 (2014).
Pastor, W. A. et al. Genome-wide mapping of 5-hydroxymethylcytosine in embryonic stem cells. Nature 473, 394–397 (2011).
Gao, F. et al. Integrated analyses of DNA methylation and hydroxymethylation reveal tumor suppressive roles of ECM1, ATF5 and EOMES in human hepatocellular carcinoma. Genome Biol. 15, 533 (2014).
Hashimoto, H. et al. Recognition and potential mechanisms for replication and erasure of cytosine hydroxymethylation. Nucleic Acids Res. 40, 4841–4849 (2012).
Valinluck, V. & Sowers, L. C. Endogenous cytosine damage products alter the site selectivity of human DNA maintenance methyltransferase DNMT1. Cancer Res., 67, 946–950 (2007).
Ye, C. & Li, L. 5-hydroxymethylcytosine: a new insight into epigenetics in cancer. Cancer Biol Ther. 15, 10–5 (2014).
Ficz, G. et al. Dynamic regulation of 5-hydroxymethylcytosine in mouse ES cells and during differentiation. Nature 473, 398–402 (2011).
Chapman, C. G. et al. Corrigendum: TET-catalyzed 5-hydroxymethylcytosine regulates gene expression in differentiating colonocytes and colon cancer. Sci Rep. 28, 24963 (2016).
Kuhn, H., Walther, M. & Kuban, R. J. Mammalian arachidonate 15-lipoxygenases structure, function and biological implications. Prostaglandins Other Lipid Mediat. 68–69, 263–290 (2002).
Brash, A. R., Boeglin, W. E. & Chang, M. S. Discovery of a second 15S-lipoxygenase in humans. Proc. Natl Acad. Sci. USA 94, 6148–6152 (1997).
Nixon, J. B., Kim, K. S., Lamb, P. W., Bottone, F. G. & Eling, T. E. 15-Lipoxygenase-1 has anti-tumorigenic effects in colorectal cancer. Prostaglandins Leukot Essent Fatty Acids 70, 7–15 (2004).
Shureiqi, I. et al. Decreased 13-S-hydroxyoctadecadienoic acid levels and 15-lipoxygenase-1 expression in human colon cancers. Carcinogenesis 20, 1985–1995 (1999).
Heslin, M. J. et al. Tumor-associated down-regulation of 15-lipoxygenase-1 is reversed by celecoxib in colorectal cancer. Ann. Surg. 241, 941–946, discussion 6–7 (2005).
Jiang, W. G., Watkins, G., Douglas-Jones, A. & Mansel, R. E. Reduction of isoforms of 15-lipoxygenase (15-LOX)-1 and 15-LOX-2 in human breast cancer. Prostaglandins Leukot Essent Fatty Acids 74, 235–245 (2006).
Moussalli, M. J. et al. Mechanistic contribution of ubiquitous 15-lipoxygenase-1 expression loss in cancer cells to terminal cell differentiation evasion. Cancer Prev. Res. (Phila) 4, 1961–1972 (2011).
Hennig, R. et al. 15-lipoxygenase-1 production is lost in pancreatic cancer and overexpression of the gene inhibits tumor cell growth. Neoplasia 9, 917–926 (2007).
Zuo, X. et al. 15-Lipoxygenase-1 transcriptional silencing by DNA methyltransferase-1 independently of DNA methylation. FASEB J. 22, 1981–1992 (2008).
Schally, A. V., Varga, J. L. & Engel, J. B. Antagonists of growth-hormone-releasing hormone: an emerging new therapy for cancer. Nat Clin Pract Endocrinol Metab. 4, 33–43 (2008).
Mebratu, Y. & Tesfaigzi, Y. How ERK1/2 activation controls cell proliferation and cell death: Is subcellular localization the answer? Cell Cycle 8, 1168–1175 (2009).
Hohla, F. et al. Differential expression of GHRH receptor and its splice variant 1 in human normal and malignant mucosa of the oesophagus and colon. Int J Oncol. 33, 137–43 (2008).
Hibi, K. et al. Detection of TFPI2 methylation in the serum of colorectal cancer patients. Cancer Lett. 311, 96–100 (2011).
Hibi, K. et al. Methylation of TFPI2 no longer detected in the serum DNA of colorectal cancer patients after curative surgery. Anticancer Res. 32, 787–790 (2012).
Hibi, K. et al. Methylation of TFPI2 gene is frequently detected in advanced well-differentiated colorectal cancer. Anticancer Res. 30, 1205–1207 (2010).
Xu, X., Zur Hausen, A., Coy, J. F. & Löchelt, M. Transketolase-like protein 1 (TKTL1) is required for rapid cell growth and full viability of human tumor cells. Int J Cancer 124, 1330–1337 (2009).
Hu, L. H. et al. The TKTL1 gene influences total transketolase activity and cell proliferation in human colon cancer LoVo cells. Anticancer Drugs 18, 427–433 (2007).
Diaz-Moralli, S., Tarrado-Castellarnau, M., Alenda, C., Castells, A. & Cascante, M. Transketolase-like 1 expression is modulated during colorectal cancer progression and metastasis formation. PLoS One 6, e25323 (2011).
Gillies, R. J., Robey, I. & Gatenby, R. A. Causes and consequences of increased glucose metabolism of cancers. J Nucl Med. 49, 24S–42S (2008).
Fearon, E. R. & Vogelstein, B. A genetic model for colorectal tumorigenesis. Cell 61, 759–767 (1990).
Smith, I. M. et al. Coordinated activation of candidate proto-oncogenes and cancer testes antigens via promoter demethylation in head and neck cancer and lung cancer. PLoS One 4, e4961 (2009).
Pelz, C. R., Kulesz-Martin, M., Bagby, G. & Sears, R. C. Global rank-invariant set normalization (GRSN) to reduce systematic distortions in microarray data. BMC Bioinformatics 9, 520 (2008).
Zhang, B., Kirov, S. & Snoddy, J. WebGestalt: an integrated system for exploring gene sets in various biological contexts. Nucleic Acids Res 33, W741–W748 (2005).
Wang, J., Duncan, D., Shi, Z. & Zhang, B. WEB-based GEne SeT AnaLysis Toolkit (WebGestalt): update 2013. Nucleic Acids Res 41, W77–W83 (2013).
Acknowledgements
This work was supported by ‘The National High Technology Research and Development Program of China’ [grant number 2012AA02A201] and ‘Scientific Research Foundation of Chongqing Medical University’ [grant number 201413].
Author information
Authors and Affiliations
Contributions
M.L., F.G. and Y.X. performed the experiments, analyzed the data and wrote the manuscript., H.L. and J.W. performed experiments. Y.T., W.Z., C.J. and Q.L. provided patient samples. Y.W. conceived the study and edited the manuscript.
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Electronic supplementary material
Rights and permissions
This work is licensed under a Creative Commons Attribution 4.0 International License. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in the credit line; if the material is not included under the Creative Commons license, users will need to obtain permission from the license holder to reproduce the material. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/
About this article

Cite this article
Li, M., Gao, F., Xia, Y. et al. Filtrating colorectal cancer associated genes by integrated analyses of global DNA methylation and hydroxymethylation in cancer and normal tissue. Sci Rep 6, 31826 (2016). https://doi.org/10.1038/srep31826
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/srep31826
This article is cited by
-
Assessment of Diagnostic Accuracy and Clinical Utility of DNA Methylation (5-mC) in Detecting Severity of Occupational Lead Exposure
Indian Journal of Clinical Biochemistry (2023)
-
DNA methylation profiles in cancer diagnosis and therapeutics
Clinical and Experimental Medicine (2018)
-
Single-molecule quantification of 5-hydroxymethylcytosine for diagnosis of blood and colon cancers
Clinical Epigenetics (2017)
-
Hypo-hydroxymethylation of rRNA genes in the precocious Eriocheir sinensis testes revealed using hMeDIP-seq
Scientific Reports (2017)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
