
Abstract
Given that E3 ubiquitin ligases (E3) regulate specific protein degradation in many cancer-related biological processes. E3 copy number variation (CNV) may affect the development and prognosis of colorectal cancer (CRC). Therefore, we detected CNVs of five E3 genes in 518 CRC patients and 518 age, gender and residence matched controls in China and estimated the association between E3 gene CNVs and CRC risk and prognosis. We also estimated their interactions with environmental factors and CRC risk. We find a significant association between the CNVs of MDM2 and CRC risk (amp v.s. wt: odds ratio = 14.37, 95% confidence interval: 1.27, 163.74, P = 0.032), while SKP2 CNVs may significantly decrease CRC risk (del v.s. wt: odds ratio = 0.32, 95% confidence interval: 0.10, 1.00, P = 0.050). However, we find no significant association between the CNVs of other genes and CRC risk. The only significant gene-environment interaction effects are between SKP2 CNVs and consumption of fish and/or fruit (P = 0.014 and P = 0.035) and between FBXW7 CNVs and pork intake (P = 0.040). Finally, we find marginally significant association between β-TRCP CNVs and CRC prognosis (amp v.s. wt, hazard ratio = 0.42, 95% confidence interval: 0.19, 0.97, P = 0.050).
Similar content being viewed by others


Introduction
Colorectal cancer (CRC) is the second most common cancer in women and the third most common in men worldwide1. In 2012, the World Health Organization estimated that about 1,360,000 new CRC cases occurred worldwide. In addition, 694,000 deaths from CRC were estimated worldwide, accounting for 8.5% of all cancer deaths and making CRC the fourth most common cause of death from cancer2. Although the relative 5-year survival rate of European CRC patients increased between 1930 and 20103, that 5-year survival rate was only 30–65% worldwide4.
Genetic susceptibility has a well-established role in the etiology of CRC5,6. Accumulating evidence supports the hypothesis that copy number variation (CNV) is a molecular biomarker for CRC risk and prognosis7. DNA CNVs, as structural variants, can be small: microscopic or submicroscopic; or they can be large: deletions, duplications or insertions, often larger than 1 kb8,9.
CNVs in the E3 ubiquitin ligases (E3) of the ubiquitin-proteasome system (UPS) have been associated with CRC risk and prognosis10,11,12. E3 plays a critical role in the specific protein degradation of UPS, which has an essential regulatory role in cell cycle progression, cell proliferation, differentiation, apoptosis, angiogenesis and cell signaling pathways13,14. The two main subfamilies of E3s are RING and HECT domain containing E3s15. As members of RING E3s, FBXW7, MDM2, SKP2 and β-TRCP have been associated with abnormal expression in some malignancies including blood, breast, colon and prostate16,17,18,19. As a HECT E3 ligase, NEDD4-1 was also proposed to play a vital role in a number of human cancers, including CRC20,21.
FBXW7 serves as a tumor suppressor gene (p53-dependent)22 and loss of FBXW7 has been associated with CRC risk and poor prognosis23. MDM2, functioning as an oncogene, is amplified in approximately one-third of all human carcinomas including CRC24. Increased expression of SKP2 has been significantly associated with poor tumor differentiation and poor prognosis in CRC18. Overexpression of β-TRCP has also been observed in many tumors, such as CRC25, pancreatic cancer26 and breast cancer27. NEDD4-1, as a HECT E3 ligase, is highly expressed in both colorectal and gastric tumor tissues20.
Studies of the CNVs of FBXW7, MDM2, SKP2, β-TRCP and NEDD4-1 genes are mainly limited to intestinal cancer cell lines and clinical pathological tissues10,11,12,19,23,25,28,29,30,31,32. In addition, most studies focus on gene expression; the impact of germline CNVs of these five genes on CRC risk and prognosis are not fully understood. Therefore, we conducted a case-control study to explore associations between the CNVs of FBXW7, MDM2, SKP2, β-TRCP and NEDD4-1 genes and CRC risk. We also followed up with cases to study the association between the CNVs of these five genes and CRC prognosis in China.
Results
Characteristics of study subjects
The basic characteristics of the 518 CRC patients and the 518 gender, age and residence matched controls are summarized in Table 1. However, 32 pairs of our samples were unable to be genotyped in one of the five genes, so gender was not equally distributed in cases and controls (P = 0.002). Education (P < 0.001), occupation (P < 0.001) and family history of other cancers (P < 0.001) were also differently distributed in cases and controls. Of the 518 CRC cases, 262 (57.8%) were colon cancer, 191 (42.2%) were rectal cancer. Gender, occupation, education and family history of cancer were adjusted in the following analysis.
Copy number variation and CRC risk
The FBXW7, MDM2, SKP2, β-TRCP, and NEDD4-1 CNVs were in Hardy-Weinberg equilibrium in all controls. Table 2 shows the CNV frequencies of the five genes and the relationship between the CNVs of the five genes and CRC risk.
We observed significant associations between MDM2 amplification and increased CRC risk (amp v.s. wt: ORadjusted = 14.37, 95% CI: 1.27, 163.74, P = 0.032; amp v.s. del + wt: ORadjusted = 14.40, 95% CI: 1.26, 164.81, P = 0.032). We observed marginally significant association between SKP2 deletions and CRC risk (del v.s. wt: ORadjusted = 0.32, 95% CI: 0.10, 1.00, P = 0.050). While there was no significant association between SKP2 amplification and CRC risk in the amp v.s. del + wt model (amp v.s. del + wt model: OR = 0.33, 95% CI: 0.11, 1.02, P = 0.055). However, we observed no significant associations between FBXW7, β-TRCP or NEDD4-1 CNVs and CRC risk.
Abnormal copy number additive model and CRC risk
In the abnormal copy number additive model, MDM2 CNVs are significantly associated with increased CRC risk (del + amp v.s. wt: ORadjusted = 6.35, 95% CI: 1.67, 24.19, P = 0.007). In the additive models, SKP2 CNVs also significantly decrease CRC risk (del + amp v.s. wt: ORadjusted = 0.32, 95% CI: 0.14, 0.72, P = 0.006).
Gene-environment interactions on CRC risk
We find a significant synergistic interaction effect between SKP2 CNVs and fruit consumption (amp v.s. del + wt: ORi = 13.89, 95% CI: 1.20, 160.57, P = 0.035) (Table 3). In addition, there is a significant interaction effect between the amplification of SKP2 and roughage consumption (≥50 g/week) (amp v.s. del + wt: OReg = 0.18, 95% CI: 0.03, 0.99). We also find significant interaction effects between the amplification of FBXW7 and consumption of roughage (≥50 g/week) or fish (>once/week) (OReg = 0.37, 95% CI: 0.15, 0.91 and OReg = 0.25, 95% CI: 0.07, 0.94, respectively). There were also significant interaction effects between the amplification of NEDD4-1 and consumption of refined grains (>250 g/week) (OReg = 2.83, 95% CI: 1.02, 7.88), Chinese pickled sour cabbage (>twice/month) (OReg = 3.59, 95% CI: 1.23, 10.48) and fatty meats (OReg = 3.60, 95% CI: 1.27, 10.19).
Gene-environment interactions in abnormal copy number additive model
We observed significant synergistic interactions between SKP2 del + amp genotype and fish intake on CRC risk (del + amp v.s. wt: ORi = 13.62, 95% CI: 1.70, 109.36, P = 0.014) (Table 3). In addition, We also observed significant interaction effects between the del + amp genotype of SKP2 and roughage consumption (≥50 g/week), or fruit (≥twice/week) consumption (del + amp v.s. wt: OReg equal to 0.13 (95% CI: 0.04, 0.44) and 0.33 (95% CI: 0.12, 0.96), respectively). We also find significant interaction effects between the del + amp genotype of MDM2 and consumption of refined grains (>250 g/week) (OReg = 5.44, 95% CI: 1.03, 28.86), fatty meats (OReg = 8.55, 95% CI: 1.22, 59.75), eggs (>3/week) (OReg = 7.33, 95% CI: 1.57, 34.30), Chinese pickled sour cabbage (>twice/month) (OReg = 27.61, 95% CI: 2.12, 259.81) and leftovers (>3 times/week) (OReg = 26.67, 95% CI: 2.62, 271.60). Moreover, we observed a significant interaction between FBXW7 CNVs and pork consumption (>250 g/week) (del + amp v.s. wt: ORi = 3.13, 95% CI: 1.06, 9.41, P = 0.040). We find significant interaction effects between the del + amp genotype of FBXW7 and consumption of refined grains (>250 g/day), fatty meats and physical exercise (OReg = 2.81 (95% CI: 1.52, 6.86), OReg = 2.30 (95% CI: 1.03, 5.11) and OReg = 0.06 (95% CI: 0.01, 0.31), respectively). Finally, we also find significant interaction effects between the del + amp genotype of NEDD4-1 and consumption of refined grains (>250 g/day), fatty meats and physical exercise (OReg = 2.75 (95% CI: 1.00, 7.55) OReg = 3.30 (95% CI: 1.21, 9.14) and OReg = 0.06 (95% CI: 0.01, 0.31), respectively).
Copy number variations and CRC prognosis
323 patients completed the follow-up (Table 4). Of the 323 patients, 186 (57.9%) patients didn’t receive chemotherapy, 45 (14.0%) patients received FOXFOX4-based chemotherapy, 22 (6.9%) received XELOX-based chemotherapy, 44 (13.7%) received LCF-based chemotherapy, 6 (1.9%) received 5-Fu-based chemotherapy and 18 (5.6%) received other chemotherapy treatments after surgery. The mean overall survival (OS) of CRC patients was 75.35 ± 2.26 months. The CEA and CA19-9 level before surgery, Dukes stage, pathological type and metastasis were adjusted for in the analysis of FBXW7, MDM2, SKP2, β-TRCP and NEDD4-1 CNVs and CRC prognosis, due to their significant association with CRC prognosis in the univariate Cox proportional hazards regression.
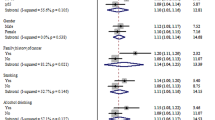
We find a marginally significant association between β-TRCP CNVs and CRC prognosis (amp v.s. wt, HRadjusted = 0.42, 95% CI: 0.19, 0.97, P = 0.050). In the additive model, β-TRCP CNVs (del + amp) is significantly associated with CRC prognosis (del + amp v.s. wt, HRadjusted = 0.39, 95% CI: 0.17, 0.88, P = 0.023) (Table 5, Fig. 1a,b). In the stratified analyses based on tumor location, the significant association between β-TRCP CNVs and CRC prognosis becomes marginally significant in rectal cancer (amp v.s. wt: HRadjusted = 0.22, 95% CI: 0.06, 0.86, P = 0.029; amp v.s. del + wt,: HRadjusted = 0.22, 95% CI: 0.06, 0.86, P = 0.029; del + amp v.s. wt: HRadjusted = 0.21, 95% CI: 0.06, 0.83, P = 0.026), but not significant in colon cancer (Table 5, Fig. 1c–e). There was no statistically significant association between other gene CNVs and colon or rectal cancer in analyses stratified by tumor location (Table 5).

Kaplan–Meier curves of overall survival (OS) according to the five genes CNVs in patients with rectal cancer.
(a) β-TRCP CNVs in CRC; (b) β-TRCP CNVs in combined model in CRC; (c) β-TRCP CNVs in rectal cancer; (d) β-TRCP amplification in rectal cancer; (e) β-TRCP CNVs in combined model in rectal cancer.
Discussion
To our knowledge, this is the first study on the association between germline CNVs of FBXW7, MDM2, SKP2, β-TRCP, NEDD4-1 and CRC risk and prognosis. In this study, MDM2 CNVs significantly increase CRC risk, while SKP2 CNVs significantly decrease CRC risk. We find evidence of three significant gene-environment interactions that increase risk of CRC: SKP2 CNVs interact with consumption of fruit and fish consumption and FBXW7 CNVs interact with pork consumption. We also observe a significant association between β-TRCP CNVs and CRC prognosis.
We observe a significant association between MDM2 amplification CNVs and CRC risk. However, there are few MDM2 amplification among patients and controls (9 and 2 respectively), which limits statistical power. Because both amplification and deletion of MDM2 can increase CRC risk, the del + amp v.s. wt model can be viewed as a conservative estimate of the effect of MDM2 on CRC risk. The amplification of MDM2 may increase CRC risk by up to 14.40-fold and the del + amp genotype of MDM2 may also increase CRC risk by 6.35-fold. MDM2 amplification was observed in 26 of 284 (9%) colorectal cancer tissue samples33, 14 of 80 (18%) CRCs tumor tissue samples34 and almost one-third of sarcomas16. MDM2 could promote tumorigenesis by acting as a positive regulator of p53 or independent of p5335. SNP data also indicate that even small differences in MDM2 levels may affect cancer risk36. Moreover, MDM2 also acts as a tumor suppressor through the Akt pathway, inducing the ubiquitination and degradation of NFAT (an invasion-promoting factor), thereby blocking cancer cell motility and invasion37. This could explain the significant association between MDM2 CNVs and increased CRC risk in the del + amp v.s. wt model in our study. There is no significant association between MDM2 CNVs and CRC prognosis. The role of MDM2 in cancer prognosis is controversial and may be affected by tumor variety and racial differences31,38,39.
We observed that SKP2 CNVs (del + amp) are significantly associated with a 68% decreased risk of CRC. The overexpression of SKP2 was associated with tumor differentiation, malignant transformation and prognosis of malignant tumors11,18. SKP2 gene amplification is commonly observed in metastatic tumors but not in early stage cancers18,40. Thus SKP2 gene amplification is likely to be associated with advanced tumor progression. In our study, 60.9% of CRC patients were in stage I or II (Table 1). This may explain the non-significance of the association between SKP2 CNVs and CRC risk. We did observe significant interactions between SKP2 CNVs and fish or fruit consumption. Fish consumption has been reported to have protective effects in CRC41, which may be attributable to the omega-3 polyunsaturated fatty acids (PUFAs) in fish42. Omega-3s function as an anti-inflammatory and is expected to have a function analogous to aspirin. Aspirin has been shown to reduce the incidence of CRC in both observational studies and randomized trials43,44. Dietary fiber in fruit is hypothesized to reduce the risk of CRC. Potential mechanisms for the protective effect dietary fibers include dilution of fecal carcinogens, reduction of transit time of feces through the bowel and increased production of short chain fatty acids45,46,47.
FBXW7 serves as a substrate adaptor for SCF ubiquitin ligase complex and mediates the recognition and binding of substrate proteins. SCFFBXW7 degrades several proteins with important roles in cell growth, proliferation, differentiation and survival48. Previous studies have reported a tumor-suppressive function of FBXW7 in colorectal tumor cells or tissues23,30 and copy number loss of FBXW7 gene in tumor tissue was reported to be significantly associated with worse CRC prognosis23. The blood level of FBXW7 expression has also been associated with the prognosis of breast cancer patients49. However, we did not observe any significant association between FBXW7 CNVs and CRC risk or prognosis. The study by Chang et al. also found a non-significant association between FBXW7 mRNA expression and CRC risk10, which is consistent with our results. About 6% of tumors harbor FBXW7 loss-of-function variants, with different variants detected in different tumor types. This might reflect tissue-specific roles of FBXW7 substrates48. A significant interaction effect has been observed between FBXW7, pork intake and increased CRC risk. An updated meta-analysis of all prospective studies showed that the risk of CRC increased by 29% for every 100 g/d of red meat consumed50. The hard muscle fibers and high fat content in red meat may be the source of this association.
We found CRC patients with β-TRCP CNVs, have a better prognosis with a 58–61% OS increase. β-TRCP is the component of the ubiquitin ligase complex targeting β-catenin and NF-ΚB for proteasome degradation, which may contribute to the inhibition of apoptosis and to tumor metastasis25. Moreover, enhanced activity of β-TRCP has been widely observed in colorectal tumor cells and primary tumors19,25. The dual function of β-TRCP might explain the significant association between β-TRCP CNVs and improved CRC prognosis. Different mechanisms of oncogenesis in rectal vs. colon cancer may explain why β-TRCP CNVs are only associated with rectal cancer prognosis in our study51. However we do not observe a significant association between β-TRCP CNVs and CRC risk. Mutations in β-TRCP are rarely detected in CRC, which is consistent with our results52,53.
Prior work has indicated that NEDD4-1 may promote tumorigenesis by decreasing PTEN protein level, or through interference with the PI3K/AKT signaling pathway54,55. NEDD4-1 is overexpressed in cancer cell lines12,50, animal models56,57,58 and in human cancer tissues59,60,61. However, we find no significant association between NEDD4-1 CNVs and CRC risk or prognosis. Meanwhile, there have been no studies focused on the effect of NEDD4-1 CNVs in peripheral blood on CRC risk and/or prognosis. One study indicated that SCFβ-TRCP can negatively regulate NEDD4-1 stability and β-TRCP-mediated destruction of the NEDD4-1 oncoprotein may inhibit cell proliferation and migration62. This suggests that epistatic effects between β-TRCP and NEDD4-1 may modify many signaling pathways. Further research is required to shed light on the relationship between these genes and any differences that may exist between their functions in the germline vs. their function in tumor.
As in any case-control or prospective survival study, we must consider the limitations of our study. First, recall bias may be inevitable in the collection of data on environment factors. Second, we collected the frequency of soybean, sausage, fried food and leftovers consumption without collecting information regarding quantity, which limits the statistical power of our analysis of gene-dietary interactions.
We find that MDM2 and SKP2 CNVs are significantly associated with CRC risk. In addition, we observe significant interaction effects between SKP2 CNVs, fish or fruit consumption and between FBXW7 CNVs and pork intake and CRC risk. There is a significant association between β-TRCP CNVs and CRC prognosis. Further research with larger sample sizes and more detailed functional evaluation will be required to confirm our results.
Materials and Methods
Subjects
After obtaining informed consent from study subjects and approval from the Institutional Research Board of Harbin Medical University, we carried out the experiment in accordance with the relevant guidelines, including any relevant details. Informed consent was obtained from all subjects. We identified CRC patients who underwent surgery at the Cancer Hospital of Harbin Medical University, based on pathologic diagnosis without pre-selection. We excluded patients with neuroendocrine carcinoma, malignant melanoma, non-Hodgkin’s lymphoma, gastrointestinal stromal tumors and Lynch syndrome CRC. From November 1st, 2004 to May 1st, 2010, we recruited 518 primary CRC patients. During the same period, we collected cancer free control subjects from the 2nd Affiliated Hospital of Harbin Medical University. We excluded controls with history of gastrointestinal disease according to self-report. 518 controls matched for age, gender and residence were recruited.
Data collection
We interviewed each participant face-to-face using a structured questionnaire with questions on demographic characteristics (age, gender, height and weight education and occupation), history of physical exercise, family history of cancer and dietary status during the 12 months preceding the interview. We collected clinical information from medical records on tumor size, Dukes stage, chemotherapy treatment, histological and pathological types and level of serum carcinoembryonic antigen (CEA) and carbohydrate antigen 19-9 (CA19-9). We followed up with 323 patients from November 2004 to March 2014. Overall survival (OS) was defined at the primary end point in our study. Survival time was calculated from the date of cancer diagnosis to death from colorectal cancer or other causes, or the time of follow-up. The date and cause of death of CRC patients were validated through the medical certification of death and the Harbin death registration system.
DNA extraction and CNV detection
We extracted DNA from all 1036 blood samples (518 CRC and 518 controls) using QIAGEN DNeasy Blood & Tissue Kit. We detected FBXW7, MDM2, SKP2, β-TRCP and NEDD4-1 copy numbers using custom designed TaqMan Copy Number Assays (Supplementary Table S1). The quantitative assays were performed using the 7500 Fast Real-Time polymerase chain reaction machine in 96-well plates with a 10 ul reaction volume containing 20 ng DNA, 5 ul TaqMan Universal PCR Master Mix, 0.5 ul of the CNV assay and 0.5 ul of the reference RNase P assay (Applied Biosystems, Carlsbad, Calif). The reaction was completed using the following cycling conditions: 95 °C for 15 seconds and 60 °C for 1 minute for 40 cycles. We used one sample with 2 copies of each CNV as a quality control in every 96-well assay plate (Supplementary Fig. 1). CNVs for each sample were detected three times. We analyzed data using 7500 software v2.0.6 (Applied Biosystems) to quantify the amplification cycle and then imported the data to Copy Caller version 2.0 (Applied Biosystems) to estimate the gene copy numbers in every sample.
Statistical analyses
We calculated the Hardy-Weinberg equilibrium in controls and compared using Fisher’s exact test. We evaluated homogeneity between cases and controls using Student’s t-test for continuous variables and a Chi-squared test for categorical variables. The unbalanced factors between the two groups were controlled for in a multivariable logistic regression for each gene and in a multivariable logistic regression for gene-environment interactions. We used odds ratios (OR) and corresponding 95% confidence intervals (95% CI) to estimate the associations between FBXW7, MDM2, SKP2, β-TRCP and NEDD4-1 CNVs and CRC risk via conditional logistic regression. We performed crossover analyses to evaluate gene-environment interaction effects on the risk of CRC with four types of OR (ORe, ORg, OReg, ORi). We adjusted the heterogeneous demography characteristics in the conditional logistic regression. We defined 2 copies as the wild type (wt), more than 2 copies as the amplification type (amp) and less than 2 copies as the deletion type(del). Two additive models were applied in the conditional logistic regression analysis: amp v.s. del + wt and del + amp v.s. wt to estimate the association between CNVs CRC risk and prognosis. All statistical tests were two-sided, P value < 0.05 in the overall analysis. Adding a Bonferroni correction, a P value < 0.025 was used in stratified analyses. We used a multiple interpolation method to fill missing values in questionnaire responses (Supplementary Tables S4–S8). All statistical analyses were performed using SAS, version 9.2 (SAS Institute Inc.Cary, NC, USA).
Additional Information
How to cite this article: Bi, H. et al. Copy number variation of E3 ubiquitin ligase genes in peripheral blood leukocyte and colorectal cancer. Sci. Rep. 6, 29869; doi: 10.1038/srep29869 (2016).
References
Jemal, A. et al. Global cancer statistics. CA: a cancer journal for clinicians 61, 69–90, doi: 10.3322/caac.20107 (2011).
Ferlay, J. et al. Cancer incidence and mortality worldwide: sources, methods and major patterns in GLOBOCAN 2012. International journal of cancer. Journal international du cancer, doi: 10.1002/ijc.29210 (2014).
Siegel, R., Desantis, C. & Jemal, A. Colorectal cancer statistics, 2014. CA: a cancer journal for clinicians 64, 104–117, doi: 10.3322/caac.21220 (2014).
Coleman, M. P. et al. Cancer survival in five continents: a worldwide population-based study (CONCORD). The Lancet. Oncology 9, 730–756, doi: 10.1016/S1470-2045(08)70179-7 (2008).
Lichtenstein, P. et al. Environmental and heritable factors in the causation of cancer–analyses of cohorts of twins from Sweden, Denmark and Finland. The New England journal of medicine 343, 78–85, doi: 10.1056/NEJM200007133430201 (2000).
Goel, A. & Boland, C. R. Recent insights into the pathogenesis of colorectal cancer. Current opinion in gastroenterology 26, 47–52, doi: 10.1097/MOG.0b013e328332b850 (2010).
Poulogiannis, G. et al. Prognostic relevance of DNA copy number changes in colorectal cancer. The Journal of pathology 220, 338–347, doi: 10.1002/path.2640 (2010).
Redon, R. et al. Global variation in copy number in the human genome. Nature 444, 444–454, doi: 10.1038/nature05329 (2006).
Stranger, B. E. et al. Relative impact of nucleotide and copy number variation on gene expression phenotypes. Science 315, 848–853, doi: 10.1126/science.1136678 (2007).
Chang, C. C. et al. FBXW7 mutation analysis and its correlation with clinicopathological features and prognosis in colorectal cancer patients. The International journal of biological markers 30, e88–e95, doi: 10.5301/jbm.5000125 (2015).
Shapira, M. et al. The prognostic impact of the ubiquitin ligase subunits Skp2 and Cks1 in colorectal carcinoma. Cancer 103, 1336–1346, doi: 10.1002/cncr.20917 (2005).
Eide, P. W. et al. NEDD4 is overexpressed in colorectal cancer and promotes colonic cell growth independently of the PI3K/PTEN/AKT pathway. Cellular signalling 25, 12–18, doi: 10.1016/j.cellsig.2012.08.012 (2013).
Mani, A. & Gelmann, E. P. The ubiquitin-proteasome pathway and its role in cancer. Journal of clinical oncology: official journal of the American Society of Clinical Oncology 23, 4776–4789, doi: 10.1200/JCO.2005.05.081 (2005).
Chen, D. & Dou, Q. P. The ubiquitin-proteasome system as a prospective molecular target for cancer treatment and prevention. Current protein & peptide science 11, 459–470 (2010).
Tsukamoto, S. & Yokosawa, H. Inhibition of the ubiquitin-proteasome system by natural products for cancer therapy. Planta medica 76, 1064–1074, doi: 10.1055/s-0029-1240901 (2010).
Oliner, J. D., Kinzler, K. W., Meltzer, P. S., George, D. L. & Vogelstein, B. Amplification of a gene encoding a p53-associated protein in human sarcomas. Nature 358, 80–83, doi: 10.1038/358080a0 (1992).
Akhoondi, S. et al. FBXW7/hCDC4 is a general tumor suppressor in human cancer. Cancer research 67, 9006–9012, doi: 10.1158/0008-5472.CAN-07-1320 (2007).
Hershko, D. D. Oncogenic properties and prognostic implications of the ubiquitin ligase Skp2 in cancer. Cancer 112, 1415–1424, doi: 10.1002/cncr.23317 (2008).
Frescas, D. & Pagano, M. Deregulated proteolysis by the F-box proteins SKP2 and beta-TrCP: tipping the scales of cancer. Nature reviews. Cancer 8, 438–449, doi: 10.1038/nrc2396 (2008).
Kim, S. S., Yoo, N. J., Jeong, E. G., Kim, M. S. & Lee, S. H. Expression of NEDD4-1, a PTEN regulator, in gastric and colorectal carcinomas. APMIS: acta pathologica, microbiologica, et immunologica Scandinavica 116, 779–784 (2008).
Li, J. et al. PTEN, a putative protein tyrosine phosphatase gene mutated in human brain, breast and prostate cancer. Science 275, 1943–1947 (1997).
Mao, J. H. et al. Fbxw7/Cdc4 is a p53-dependent, haploinsufficient tumour suppressor gene. Nature 432, 775–779, doi: 10.1038/nature03155 (2004).
Iwatsuki, M. et al. Loss of FBXW7, a cell cycle regulating gene, in colorectal cancer: clinical significance. International journal of cancer. Journal international du cancer 126, 1828–1837, doi: 10.1002/ijc.24879 (2010).
Steinman, H. A. et al. An alternative splice form of Mdm2 induces p53-independent cell growth and tumorigenesis. The Journal of biological chemistry 279, 4877–4886, doi: 10.1074/jbc.M305966200 (2004).
Ougolkov, A. et al. Associations among beta-TrCP, an E3 ubiquitin ligase receptor, beta-catenin and NF-kappaB in colorectal cancer. Journal of the National Cancer Institute 96, 1161–1170, doi: 10.1093/jnci/djh219 (2004).
Muerkoster, S. et al. Increased expression of the E3-ubiquitin ligase receptor subunit betaTRCP1 relates to constitutive nuclear factor-kappaB activation and chemoresistance in pancreatic carcinoma cells. Cancer research 65, 1316–1324, doi: 10.1158/0008-5472.CAN-04-1626 (2005).
Spiegelman, V. S. et al. Induction of homologue of Slimb ubiquitin ligase receptor by mitogen signaling. The Journal of biological chemistry 277, 36624–36630, doi: 10.1074/jbc.M204524200 (2002).
Feuk, L., Carson, A. R. & Scherer, S. W. Structural variation in the human genome. Nature reviews. Genetics 7, 85–97, doi: 10.1038/nrg1767 (2006).
Voutsadakis, I. A. The ubiquitin-proteasome system in colorectal cancer. Biochimica et biophysica acta 1782, 800–808, doi: 10.1016/j.bbadis.2008.06.007 (2008).
Guo, Z., Zhou, Y., Evers, B. M. & Wang, Q. Rictor regulates FBXW7-dependent c-Myc and cyclin E degradation in colorectal cancer cells. Biochemical and biophysical research communications 418, 426–432, doi: 10.1016/j.bbrc.2012.01.054 (2012).
Kondo, I., Iida, S., Takagi, Y. & Sugihara, K. MDM2 mRNA expression in the p53 pathway may predict the potential of invasion and liver metastasis in colorectal cancer. Diseases of the colon and rectum 51, 1395–1402, doi: 10.1007/s10350-008-9382-5 (2008).
Tian, Y. F. et al. SKP2 overexpression is associated with a poor prognosis of rectal cancer treated with chemoradiotherapy and represents a therapeutic target with high potential. Tumour biology: the journal of the International Society for Oncodevelopmental Biology and Medicine 34, 1107–1117, doi: 10.1007/s13277-013-0652-z (2013).
Forslund, A. et al. MDM2 gene amplification is correlated to tumor progression but not to the presence of SNP309 or TP53 mutational status in primary colorectal cancers. Molecular cancer research: MCR 6, 205–211, doi: 10.1158/1541-7786.MCR-07-0239 (2008).
Hav, M. et al. MDM2 gene amplification and protein expressions in colon carcinoma: is targeting MDM2 a new therapeutic option? Virchows Archiv: an international journal of pathology 458, 197–203, doi: 10.1007/s00428-010-1012-7 (2011).
Gajjar, M. et al. The p53 mRNA-Mdm2 interaction controls Mdm2 nuclear trafficking and is required for p53 activation following DNA damage. Cancer cell 21, 25–35, doi: 10.1016/j.ccr.2011.11.016 (2012).
Bond, G. L., Hu, W. & Levine, A. A single nucleotide polymorphism in the MDM2 gene: from a molecular and cellular explanation to clinical effect. Cancer research 65, 5481–5484, doi: 10.1158/0008-5472.CAN-05-0825 (2005).
Yoeli-Lerner, M. et al. Akt blocks breast cancer cell motility and invasion through the transcription factor NFAT. Molecular cell 20, 539–550, doi: 10.1016/j.molcel.2005.10.033 (2005).
Choschzick, M. et al. MDM2 amplification is an independent prognostic feature of node-negative, estrogen receptor-positive early-stage breast cancer. Cancer biomarkers: section A of Disease markers 8, 53–60, doi: 10.3233/DMA-2011-0806 (2010).
Abolhasani, M., Salarinejad, S. & Asgari, M. P53 and MDM2 Over-expression and Five-year Survival of Kidney Cancer Patients Undergoing Radical Nephrectomy-Iranian Experience. Asian Pacific journal of cancer prevention: APJCP 16, 5043–5047 (2015).
Li, J. Q. et al. Correlation of Skp2 with carcinogenesis, invasion, metastasis and prognosis in colorectal tumors. International journal of oncology 25, 87–95 (2004).
Iscovich, J. M. et al. Colon cancer in Argentina. II: Risk from fibre, fat and nutrients. International journal of cancer. Journal international du cancer 51, 858–861 (1992).
Bang, H. O., Dyerberg, J. & Sinclair, H. M. The composition of the Eskimo food in north western Greenland. The American journal of clinical nutrition 33, 2657–2661 (1980).
Bosetti, C., Rosato, V., Gallus, S., Cuzick, J. & La Vecchia, C. Aspirin and cancer risk: a quantitative review to 2011. Annals of oncology: official journal of the European Society for Medical Oncology/ESMO 23, 1403–1415, doi: 10.1093/annonc/mds113 (2012).
Rothwell, P. M. et al. Long-term effect of aspirin on colorectal cancer incidence and mortality: 20-year follow-up of five randomised trials. Lancet 376, 1741–1750, doi: 10.1016/S0140-6736(10)61543-7 (2010).
Lipkin, M., Reddy, B., Newmark, H. & Lamprecht, S. A. Dietary factors in human colorectal cancer. Annual review of nutrition 19, 545–586, doi: 10.1146/annurev.nutr.19.1.545 (1999).
Hill, M. J. Diet and chemoprevention of colorectal cancer. Tumori 81, 5–6 (1995).
Park, Y. et al. Dietary fiber intake and risk of colorectal cancer: a pooled analysis of prospective cohort studies. Jama 294, 2849–2857, doi: 10.1001/jama.294.22.2849 (2005).
Welcker, M. & Clurman, B. E. FBW7 ubiquitin ligase: a tumour suppressor at the crossroads of cell division, growth and differentiation. Nature reviews. Cancer 8, 83–93, doi: 10.1038/nrc2290 (2008).
Yumimoto, K. et al. F-box protein FBXW7 inhibits cancer metastasis in a non-cell-autonomous manner. The Journal of clinical investigation 125, 621–635, doi: 10.1172/JCI78782 (2015).
Chan, D. S. et al. Red and processed meat and colorectal cancer incidence: meta-analysis of prospective studies. PloS one 6, e20456, doi: 10.1371/journal.pone.0020456 (2011).
Delattre, O. et al. Multiple genetic alterations in distal and proximal colorectal cancer. Lancet 2, 353–356 (1989).
Chiaur, D. S. et al. Five human genes encoding F-box proteins: chromosome mapping and analysis in human tumors. Cytogenetics and cell genetics 88, 255–258, doi: 15532 (2000).
Reifenberger, J. et al. Molecular genetic analysis of malignant melanomas for aberrations of the WNT signaling pathway genes CTNNB1, APC, ICAT and BTRC. International journal of cancer. Journal international du cancer 100, 549–556, doi: 10.1002/ijc.10512 (2002).
Chen, C. & Matesic, L. E. The Nedd4-like family of E3 ubiquitin ligases and cancer. Cancer metastasis reviews 26, 587–604, doi: 10.1007/s10555-007-9091-x (2007).
Wang, X. et al. NEDD4-1 is a proto-oncogenic ubiquitin ligase for PTEN. Cell 128, 129–139, doi: 10.1016/j.cell.2006.11.039 (2007).
Liu, Y., Oppenheim, R. W., Sugiura, Y. & Lin, W. Abnormal development of the neuromuscular junction in Nedd4-deficient mice. Developmental biology 330, 153–166, doi: 10.1016/j.ydbio.2009.03.023 (2009).
Kawabe, H. et al. Regulation of Rap2A by the ubiquitin ligase Nedd4-1 controls neurite development. Neuron 65, 358–372, doi: 10.1016/j.neuron.2010.01.007 (2010).
Fouladkou, F. et al. The ubiquitin ligase Nedd4-1 is required for heart development and is a suppressor of thrombospondin-1. The Journal of biological chemistry 285, 6770–6780, doi: 10.1074/jbc.M109.082347 (2010).
Jung, S. et al. Oncogenic function of p34SEI-1 via NEDD41mediated PTEN ubiquitination/degradation and activation of the PI3K/AKT pathway. International journal of oncology 43, 1587–1595, doi: 10.3892/ijo.2013.2064 (2013).
Tanksley, J. P., Chen, X. & Coffey, R. J. NEDD4L is downregulated in colorectal cancer and inhibits canonical WNT signaling. PloS one 8, e81514, doi: 10.1371/journal.pone.0081514 (2013).
Wang, Y. Y., Ye, Z. Y., Zhao, Z. S., Tao, H. Q. & Li, S. G. Systems biology approach to identification of biomarkers for metastatic progression in gastric cancer. Journal of cancer research and clinical oncology 136, 135–141, doi: 10.1007/s00432-009-0644-y (2010).
Liu, J. et al. SCF(beta-TRCP)-mediated degradation of NEDD4 inhibits tumorigenesis through modulating the PTEN/Akt signaling pathway. Oncotarget 5, 1026–1037 (2014).
Acknowledgements
We sincerely thank Ryan J. Bohlender for reviewing and revising the manuscript during the manuscript revision. This work was supported by grants from Natural Science Foundation of China (Grant No. 81302483 and 30972539), the fifty-second batch of the Postdoctoral Science Foundation of P. R. China (Grant No. 2012M520773), the Postdoctoral Science Foundation of the government of Heilongjiang Province (Grant No. LBH-Z11070).
Author information
Authors and Affiliations
Contributions
F.H., Y.Z. and G.W. designed the study, directed its implementation, including quality assurance and control and reviewed the manuscript. H.B. and T.T. did the experiments. H.B. did the data analysis and wrote the manuscript. L.Z., H.Z. and X.L. helped the study’s analytic strategy. H.H. and Y.L. helped the questionnaire data collection and experiments conduction.
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Electronic supplementary material
Rights and permissions
This work is licensed under a Creative Commons Attribution 4.0 International License. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in the credit line; if the material is not included under the Creative Commons license, users will need to obtain permission from the license holder to reproduce the material. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/
About this article

Cite this article
Bi, H., Tian, T., Zhu, L. et al. Copy number variation of E3 ubiquitin ligase genes in peripheral blood leukocyte and colorectal cancer. Sci Rep 6, 29869 (2016). https://doi.org/10.1038/srep29869
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/srep29869
This article is cited by
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
