
Abstract
Hypoxia-inducible factors (HIF) are heterodimeric transcription factors that allow cells to adapt and survive during hypoxia. Regulation of HIF1A and HIF2A mRNA is well characterized, whereas HIF3A mRNA regulation and function are less clear. Using RNA-Seq analysis of primary human umbilical vein endothelial cells, we found two isoforms of HIF3A were expressed, HIF3A2 and HIF3A3. Comparing HIF3A expression profiles to HIF1A mRNA during 48 hours of hypoxia revealed that HIF1A message peaked at 4 hours, whereas HIF3A expression increased while HIF1A was decreasing. Given that HIF1A mRNA is regulated by miR-429, we tested miR-429 effects on both HIF3A isoforms and found that they too were regulated by miR-429. Analysis of a HIF-3 target, DNA-damage-inducible transcript 4, a key survival gene, indicated that DDIT4 mRNA is induced by HIF-3 and negatively regulated by miR-429 through miR-429’s actions on HIF3A message. This provides a compelling model for how hypoxia-induced miR-429 regulates the switch between HIF-1 adaptive responses to HIF-3 survival responses by rapidly decreasing HIF1A levels while simultaneously slowing the progression of HIF3A expression until the miR-429 levels drop below normoxic levels. Since HIF-1 drives HIF3A and miR-429 expression, this establishes a regulatory network in which miR-429 plays a pivotal role.
Similar content being viewed by others


Introduction
Ischemia-reperfusion injury results in tissue damage in organs such as the heart where hypoxia and reperfusion lead to fibrotic remodeling and cardiomyocyte apoptosis1. During the initial stages of hypoxia, a cellular adaptive response is induced that minimizes cell death through activation of three transcription factors called hypoxia-inducible factors (HIFs) that respond to low tissue oxygen levels and stimulate the expression of a number of genes that promote angiogenesis, energy metabolism and cell survival2,3. The genes activated by HIF-1 enhance oxygen delivery to the tissues and/or promote cellular metabolic adaption to the reduced oxygen levels3. HIF-1, HIF-2 and HIF-3 are tightly regulated through changes in oxygen tension4,5. They consist of stable, constitutively expressed β subunits that associate with the oxygen destabilized α subunits6. During normoxia, postranslational hydroxylations of alpha subunits target them for polyubiquitination and degradation7,8. During hypoxia, however, the alpha subunits are stable and translocate to the nucleus to dimerize with the β subunits to create transcriptionally active HIF-1, HIF-2 and HIF-3 complexes5.
HIF-1 and HIF-2 are widely accepted as major mediators of the hypoxic response9. The HIF-1 and HIF-2 function as transcriptional activators and have both unique and overlapping target genes. HIF-1 governs initial adaptation to hypoxia, whereas HIF-2 expression begins after more prolonged oxygen depletion9,10. Understanding the exact roles of HIF-3 are complicated by the fact that there are alternatively spliced variants of HIF3A and HIF3A utilization of different promoters results in at least four different mRNA variants that code for six or more isoforms11,12,13. Furthermore, overexpression of the HIF-3α isoforms, suppresses HIF-1 and HIF-2 activity in cell culture14,15. Recent studies, however, indicate that human HIF-3 isoforms have transcriptional functions that partially overlap with that of HIF-1, indicating that HIF-3 is more than just a dominant-negative form that suppresses HIF-1 and HIF-2 function16,17.
Previous studies have demonstrated that a number of microRNAs (miRNAs) are induced during hypoxia and play critical roles in angiogenesis18,19,20. Some of these miRNAs, miR-210-3p and miR-485-5p, have been shown to regulate HIF-3A mRNA in soft tissue sarcoma cells21. Our previous studies demonstrated that miR-429 is induced during hypoxia in primary human umbilical vein endothelial cells (HUVECs) and regulates HIF-1 mRNA levels during the early stages of hypoxia10.
Here we examined whether miR-429 played any role in the later stages of hypoxia and interestingly, found an inverse relationship between HIF3A mRNA levels versus miR-429 and HIF1A expression. We analyzed this in more detail and found that miR-429 binds directly to HIF3A2 and HIF3A3 3′UTRs and regulates their expression. We also demonstrate that miR-429 regulates a transcriptional target of HIF-3, DNA-damage-inducible transcript 4 (DDIT4), a pro-survival protein that inhibits the mechanistic target of rapamycin (mTOR) complex 1 (mTORC1). Our studies suggest that miR-429, which is induced early in the hypoxia time-course, regulates the transitional switch between HIF-1 and HIF-3 responses in HUVECs during prolonged hypoxia by first attenuating HIF-1 responses during the early stages of hypoxia by destabilizing HIF1A message and then delaying the onset of HIF3A message stability in the later stages. Given that HIF-1 drives the expression of HIF3A and miR-429, this establishes a regulatory network in which miR-429 plays a critical role.
Results
HIF3A2 and HIF3A3 mRNAs are expressed in human endothelial cells
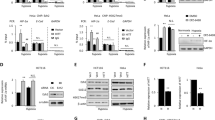
Given the complexity of the HIF3A locus, we used next generation sequencing analysis followed by qRT-PCR validation to determine which HIF3A isoforms were expressed in HUVECs. The data indicates that HIF3A is expressed at low levels and only isoforms HIF3A2 and HIF3A3 were detected (Supplemental Figure 1). These two HIF3A isoforms are annotated in the RefSeq database (ncbi.nlm.nih.gov/refseq/: NM_022462.4 and NM_152795.3, respectively). The protein expression supported the mRNA analyses and demonstrated that HIF-3α2 protein is expressed at a higher level than HIF-3α3 (64 kDa form versus the 72 kDa form, respectively (Fig. 1A). In our protein analysis, furthermore, it was difficult to the detect HIF3α3 isoform unless the blots were overexposed as shown in (Fig. 1A and therefore we focused our subsequent analyses on HIF-3α2. To follow the expression of HIF3A message levels during hypoxia, the HUVECs were treated from 0 to 48 h in 1% oxygen and the mRNA levels were measured (Fig. 1B). The results indicate that HIF3A mRNA, a measure of both HIF3A2 and HIF3A3, is reduced below normoxic levels after 4 hours of hypoxia and then gradually increases ~3-fold by 24 hours and remains elevated for 48 hours. To evaluate the consequences of hypoxia on the protein expression levels of the HIF-3α, we monitored expression by western blot analysis. The HIF-3α2 proteins levels mirrored the mRNA levels with the exception of the 48 hours time point that appeared similar to the normoxic start conditions (Fig. 1C,D). Interestingly, the HIF-3α2 protein levels do not begin to rise above normoxic levels until 8 hours of hypoxia. At 4 hours, the HIF-3α2 protein levels drop significantly and follow the mRNA changes, although the mRNA levels appear to rebound much faster than the corresponding protein changes.

Expression of HIF3α isoforms in HUVECs.
(A) The anti-HIF-3α (Sigma-Aldrich, AV39936) antibody allows detection of HIF-3α2 (~72 kDa) and when overexposed, HIF-3α1 (~72 kDa). The HUVECs cell lysate is shown in lane 1 (5 μq loaded) and the molecular weight markers (M) are shown in lane 2 (Precision plus Kaleidoscope, BioRad #161-0375). (B) Hypoxia induces dynamic changes in the mRNA expression profiles of the HIF3A2–3 variants in HUVECs. The mRNA levels were monitored in qRT-PCR experiments. The results from 3 independent experiments (n = 12) are plotted normalized to TBP mRNA levels and expressed as a fold-change over the normoxic control. Error bars represent standard deviations (*P < 0.05). (C) Hypoxia induces dynamic changes of protein levels of HIF-3α2 and (D) the bar graphs show the relative protein amounts at each time point. The protein levels of were detected with SDS-PAGE and Western Blot and related to total protein levels. 2 individual samples (4 μg of total protein per lane) were tested for each time point and the experiments were repeated twice. The protein levels (bar graphs) are normalized to the normoxic control. Significant changes (p < 0.05) are marked with an “*”.
Given that HIF-3α is regulated by HIF-1 but not HIF-213, we next examined the mRNA levels of HIF1A under these same conditions (Fig. 2A). The results indicate that HIF1A mRNA was rapidly elevated at 4 hours, but decreases dramatically below the normoxic levels after 12 hours of hypoxia. The protein profile of HIF-1α correlated with the mRNA analysis with the protein elevated from 2 to 8 hours (Fig. 2B,C). The HIF1A profile negatively correlated with the HIF3A mRNA, emphasizing the temporal segregation between the HIF1A and HIF3A expression patterns in HUVECs (Supplemental Figure 2).

(A) Hypoxia induces dynamic changes in the mRNA and protein expression profile of the HIF1A in HUVECs. The mRNA levels were monitored in qRT-PCR experiments. The results from 3 independent experiments (n = 12) are plotted normalized to TBP mRNA levels and expressed as a fold-change over the normoxic control. Error bars represent standard deviations. (B) Hypoxia induces dynamic changes of protein levels of HIF-1α and (C) the bar graphs show the relative protein amounts at each time point. The protein levels of were detected with SDS-PAGE and Western Blot and related to total protein levels. 2 individual samples (4 μg of total protein per lane) were tested for each time point and the experiments were repeated twice. The protein levels (bar graphs) are normalized to the normoxic control. Significant changes (p < 0.05) are marked with an “*”. (D) The predicted target site of miR-429 in HIF3A2-3. The localization of putative miR-429 sequence is shown aligned with the 3′UTR of HIF3A isoforms. The numbering is based on the human NCBI genomic sequence (RefSeq). (E) Hypoxia-induced changes in the expression profile of miR-429 in HUVECs. The miRNA level was monitored in quantitative real-time PCR experiments. The results from 3 independent experiments (n = 18) are plotted normalized to RNU44 and expressed as a fold change over the normoxic control (*P < 0.05).
In our previous studies, we found that miR-429 was induced by HIF-1 and that HIF1A mRNA was regulated by miR-429 in a negative feedback loop10. In examining the 3′UTR of the HIF3A isoforms using RNAhybrid software22, we identified a potential miR-429 targeting sequence in the HIF3A 3′UTRs (Fig. 2D). To establish whether there was a correlation between miR-429 and the HIF3A mRNA expression during hypoxia, we examined the same HUVEC samples used for the mRNA analyses and monitored for miR-429 expression. The results shown in Fig. 2E demonstrate that miR-429 was elevated ~13-fold at 4 hours and then rapidly declined below normoxic levels by 12 hours and remained low for the duration of the time course. Interestingly, the miR-429 profile negatively correlated with the HIF3A mRNA, suggesting that miR-429 expression could be responsible for initial reduction of HIF3A levels (Supplemental Figure 2).
miR-429 regulates HIF3A mRNA steady-state levels
We next tested whether miR-429 affected HIF3A mRNA and HIF-3α2 protein levels by transfecting the HUVECs with miR-429 mimics or antagomirs and followed their effects on HIF3A mRNA expression under both normoxic and hypoxic conditions. We first established that the transfection efficiencies promoted appropriate effects on the miR-429 levels in the HUVECs (Supplemental Figure 3). Examining the functional effects indicated that inhibition of miR-429 with antagomir 429 resulted in the accumulation of HIF3A mRNA, whereas miR-429 mimic expression reduced HIF3A mRNA levels (Fig. 3A). Furthermore under these same conditions, the HIF-3α2 protein levels followed the same pattern as the mRNA except for the miR-429 mimic transfection during normoxic conditions (Fig. 3B,C). Given the instability of the HIF3α-chains during normoxic conditions, however, this could explain the limited correlation here.

(A) miR-429 alters the expression of HIF3A. HUVECs were transfected with miR-429 mimic or antagomir and the mRNA levels were monitored in normoxic conditions and after 4 h and 8 h of exposure to hypoxia. HIF3A mRNA levels from 3 independent experiments (n = 12) are plotted normalized to 18S rRNA levels and expressed as a fold change over the transfection control. Significant changes (p < 0.05) are marked with an “*”. (B) The corresponding changes of HIF-3α2 protein levels of were detected with SDS-PAGE and Western Blot and normalized to the β-actin and total protein levels. (C) The bar graphs show the relative protein amounts at each condition normalized to the transfection control. (D) 2 individual samples (4 μg of total protein per lane) were tested for each treatment and the experiments were repeated twice. Mimic 429 (m429) and antagomir 429 (ant429) are shown for each condition.
In order to test if miR-429 directly interacted with the predicted HIF3A target sequence that we identified, we used a specific target protector to inhibit the interaction between miR-429 and the HIF3A mRNA. Target protectors are modified RNAs complementary to miRNA target sequences that bind to the specific mRNA 3′UTR target sequence and block miRNA binding, thus preventing the formation of the predicted miRNA-mRNA complex shown in Fig. 2B23. As shown in Fig. 4A, the HIF3A target protector (TP) blocked the miR-429 mimic effects on the HIF3A mRNA levels. The corresponding protein changes in these same samples showed that the HIF3A TP increased the HIF-3α2 levels, blocked the effect of mimic 429 and that mimic 429 alone dramatically decreased HIF-3α2 protein levels below normoxic control levels (Fig. 4B). The results suggested that the predicted target sequence identified was correct and that under hypoxic conditions miR-429 regulates the levels of the HIF3A mRNA and protein levels.

miR-429 binds to predicted target sequence in the HIF3A1-3 3′UTR.
(A) HUVECs were transfected with HIF3A target sequence-specific target protector and/or the miR-429 analog (Mimic 429). Two days after transfection, the cells were placed in hypoxia for 4 h and the HIF3A mRNA levels were monitored in qRT-PCR experiments. The HIF3A levels results from 3 independent experiments (n = 12) are plotted normalized to TBP mRNA levels and expressed as a fold change over the transfection control. Significant changes (p < 0.05) are marked with an “*”. The corresponding changes of HIF-3α2 protein levels were detected with SDS-PAGE and Western Blot and normalized to the β-actin levels and total protein levels. (B) 2 individual samples (3 μg of total protein per lane) were tested for each treatment and the experiments were repeated twice.
miR-429 affects the HIF3A target DDIT4 mRNA levels indirectly
In order to test if miR-429 has a functional influence on HIF-3α targets, we analyzed the hypoxia-driven expression of gene previously identified as a HIF-3α transcriptional target in HEK293 cells, DDIT416. DDIT4 regulates cell growth and survival via inhibition of the activity of mTORC1 and therefore the induction of this gene during hypoxia promotes cell survival24,25,26. First, we followed the hypoxia time-course for DDIT4 mRNA as described before and found that its expression was biphasic with a first peak at 4 hours (Fig. 5A). To test if miR-429 affected DDIT4 mRNA levels during normoxia or hypoxia, the cells were transfected with mimic 429 and antagomir 429. The results shown in Fig. 5B indicated that DDIT4 mRNA levels were affected significantly with both mimic and antagomir at 4 and 8 hours of hypoxia. During normoxic conditions, the antagomir, but not the mimic, had a significant effect.

Hypoxia induces dynamic changes in expression profiles of DDIT4 (A) in HUVECs. The results from 3 independent experiments (n = 18) are plotted normalized to TBP mRNA levels and expressed as a fold-change over the normoxic control. Error bars represent standard deviations. miR-429 mimic and antagomir modulate DDIT4 expression. (B) HUVECs were transfected with miR-429 mimic or antagomir and the mRNA levels were monitored in qRT-PCR experiments in normoxic conditions and after 4 and 8 hours of hypoxia. The mRNA levels from 3 independent experiments (n = 12) were normalized TBP mRNA levels and expressed as a fold change over the transfection control. miR-429 indirectly modulates the expression of DDIT4 mRNA levels during hypoxia through its actions on HIF3A mRNA. (C) mRNA levels were monitored in qRT-PCR experiments following transfection of HUVECs with HIF3A TP and HIF1A TP together with miR-429 analog (Mimic 429). The mRNA levels results from 3 independent experiments (n = 12) are normalized to TBP mRNA levels and expressed as a fold-change over the transfection control. Significant changes (p < 0.05) are marked with an “*”.
Given that the mimic did not affect the DDIT4 mRNA during normoxia, this suggested that the miR-429 effect on DDIT4 could be indirect. To test for this, we transfected the HUVECs with HIF3A and HIF1A target protectors and examined their ability to block the mimic 429 responses after 4 hours of hypoxia. The results shown in Fig. 5C show that the HIF3A target protector, but not HIF1A target protector, blocked the mimic 429 decrease in DDIT4 mRNA levels, indicating that miR-429 was inhibiting DDIT4 expression through its actions on HIF3A mRNA and therefore miR-429 effects on DDIT4 mRNA expression were indirect.
To confirm that HIF-1 enhances the expression of HIF3A and miR-429, we chemically enhanced HIF-1α stability and followed the levels of HIF3A mRNA and miR-429 during this treatment (Supplemental Figure 4). The HUVECs were treated with chemical hypoxia mimetics 100 μM desferoxamine (DFO) or 200 μM CoCl2 that inhibit proline hydroxylase activity during normoxia. This treatment stabilizes HIF-1α protein and leads to HIF-1 transcriptional activity27,28. The results show that the DFO treatment had the greatest effect in increasing HIF3A and miR-429 expression (Supplemental Figure 4, white bars). To confirm that the effects were due to HIF-1 activity, a functional inhibitor of HIF-1, topotecan (500 μM TPT)29, was used in combination with the two different hypoxia-mimetics and the results show that this inhibited the hypoxia-mimetic treatment (Supplemental Figure 4, black bars), confirming that HIF-1 was enhancing both HIF3A and miR-429 expression.
miR-429 regulates the levels of HIF3A and HIF1A during hypoxia
To establish the physiological effects of miR-429 during the hypoxia time course in HUVECs and its corresponding effects on HIF3A and HIF1A expression, the cells were transfected with 429 antagomir and then followed over a 48-hour hypoxia time course. The results demonstrate that miR-429 is normally active up to 36 hours since the antagomir dramatically increased the levels HIF3A message out to that time point (Fig. 6A). For HIF1A message, the antagomir only had pronounced effects during the early stages since by 8–12 hours; the HIF1A message was back to normoxic levels (~8 hours) and substantially below normoxic levels from 16 to 48 hours (Fig. 6B). These results indicate that miR-429 regulates both the early and late stages of the hypoxic response in HUVECs by regulating the mRNA levels of both HIF1A and HIF3A.

miR-429 regulates the levels of HIF3A and HIF1A mRNAs during normoxic and hypoxic conditions in HUVECs.
HUVECs were transfected with miR-429 antagomir and the mRNA levels were monitored over the 48 h time course for HIF3A (A) and HIF1A (B). The mRNA levels were monitored in qRT-PCR experiments. The results from 2 independent experiments (n = 6) are plotted normalized to 18S rRNA levels and expressed as a fold-change over the normoxic transfection control. Error bars represent standard deviation. Significant changes (p < 0.05) are marked with an “*”.
Discussion
HIF-1 is the master regulator for the expression of genes in response to hypoxia in most mammalian cells and its function and regulation are well established3. Much less is known about HIF-3 and it has been suggested that its expression is much more restricted7. Although HIF-1 and HIF-2 have 48% sequence identity, HIF-3 is very distinct from the other two hypoxia-induced transcription factors7. Previous studies in A549 cells (a human alveolar cell carcinoma cell line) indicated that HIF1A mRNA levels did not increase during hypoxia or when treated with 250 mM CoCl2, whereas HIF3A message increased at 4 hours and was maximal at 8 hours17. The differences between their studies and ours could be explained by differences in alveolar and endothelial cell regulation or because of the differences between transformed and primary cells. Our time course was much more extended, 48 hours versus 16 and an interesting feature in our studies was the clear demarcation between the HIF1A and HIF3A mRNA expression patterns and the fact that the HIF3A message was still elevated at 36 hours. We also demonstrate that a microRNA, miR-429, plays a key regulatory role in mediating this demarcation.
In previous studies, we demonstrated that HIF-1 drives the expression of miR-42910. Here we demonstrate that miR-429 attenuates the expression of HIF3A during the early stages of hypoxia. During normoxic conditions, the protein expression levels of HIF-1α and HIF-3α are regulated through postranslational hydroxylations of the alpha subunits that lead to subsequent polyubiquitination and degradation8. During hypoxia, the HIF-1α and HIF-3α proteins are stable. In the initial stages of hypoxia, the adaptive phase is driven by HIF-1. HIF-1 accumulation leads to induction of miR-429 expression that subsequently targets HIF1A mRNA in a negative feedback loop10. miR-429 also provides the switch between HIF-1 and HIF-2 activities since it reduces HIF1A message before HIF2A expression is induced at 12 h10. HIF2A does not have a miR-429 targeting sequence in its 3′UTR and therefore its message is not predicted to be affected by miR-429 expression, suggesting that miR-429 effects are limited to HIF1A and HIF3A mRNA.
HIF1A levels go rapidly up during hypoxia and then decline rapidly based on the actions of miR-42910. Here, we demonstrate that miR-429 also regulates HIF3A levels and miR-429 attenuates the expression until the later time points, thus regulating the transition from HIF1A to HIF3A and presumably the temporal expression of their target genes in HUVECs. During hypoxia, HIF1A is up at 4 hours, HIF2A is up by 12 hours10 and HIF3A is initially down at 4 hours and then gradually rises from 12 to 24 h at a time when the miR-429 levels are low. The role for miR-429 is best demonstrated when the cells are transfected with 429 antagomir. Under these conditions, HIF3A levels are elevated from the start, even during normoxic conditions. Clearly, part of this elevation is mediated by the increases in HIF1A expression (Fig. 6B); however, the HIF3A target protector studies demonstrate that this is a direct effect on HIF3A as well (Fig. 4A).
As shown in Supplemental Figure 1, the HIF3A mRNA levels are very low (up to 50-fold lower than that of HIF1A). Hence, the transcriptional increase of miR-429 levels during 4 h time point will not affect the HIF3A mRNA levels since these are already very low, in part due to the miR-429 steady-state normoxic levels. At later time points, the decrease in miR-429 levels in combination with the HIF-1-dependent induction of HIF3A expression and the long half-life of HIF3A mRNA17, drives the equilibrium towards HIF3A mRNA accumulation. This statement is supported by the miR-429 inhibitor data illustrating that HIF3A expression increases during both normoxia and hypoxia (Fig. 3). More importantly, HIF3A mRNA and miR-429 are only part of mechanism that controls HIF17 expression during hypoxia. The changes in HIF-3α expression result from the combined effects of HIF-1, the hypoxia impaired activity of PHDs and FIH, miRNAs and potentially other factors. Furthermore, miR-429 affects numerous other hypoxia-related targets including HIF-110. Therefore, the correlation between the HIF3A and miR-429 levels during the hypoxia time course, although is supported by miRNA analog studies, results from a number of multiple co-dependent factors.
The transition from HIF1A to HIF2A to HIF3A may be critical for cell survival during prolonged periods of hypoxia. This successful transition is further supported by the early HIF1A induction that drives the expression of HIF3A. HIF-1 and HIF-3 are both strongly connected by miR-429 regulation as well as by the fact that HIF-1, but not HIF-2, drives the expression of HIF3A13. The functional relationship between HIF-1 and HIF-3 has two main consequences. First, it allows for the switch between HIF-1 and HIF-3 signaling in the HUVECs. And secondly, it prevents the continuous HIF-1 transcriptional activity through the reported dominant negative function of HIF-3α15. In examining the functional consequences of HIF-3α expression, we tested a potential transcriptional target, DDIT4. Our studies support the view that HIF-3 induces the pro-survival gene DDIT4 and this induction is modulated by miR-429 through its direct interactions with HIF3A message. However, the physiological consequences of miR-429 interaction with HIF3A on DDIT4 expression require further studies.
In summary, our studies establish a model for how a microRNA that is induced during hypoxia can regulate the HIF-1/HIF-3 transition in HUVECs during hypoxia. In this model, normal oxygen levels in the HUVECs and moderate levels of miR-429 maintain low levels of HIF1A and HIF3A mRNA (Fig. 7). HIF-1α and HIF-3α protein levels remain low through posttranslational degradation of these transcription factors when the oxygen levels are normal (reviewed in)30. During the early stages of hypoxia, HIF1A message and HIF-1α protein are dramatically elevated, while the HIF3A mRNA isoforms and protein remain low. HIF-1 drives a number of pro-survival and angiogenic target genes and also promotes miR-429 and HIF3A up-regulation. miR-429 inhibits the message stabilities of both HIF1A and HIF3A. During prolonged hypoxia, HIF-1α levels and miR-429 are lower and HIF3A message and HIF-3α protein accumulate over time, leading to the induction of HIF-3 pro-survival gene expression. The model shown here provides a basis for how a miRNA, which is induced early in the hypoxia time-course, regulates the transitional switch between HIF-1 and HIF-3 responses in HUVECs during prolonged hypoxia and thus regulates the transition between the induction of the early response genes to the later stages of hypoxia where other targets are induced to provide the cell with a mechanism for surviving chronic stress conditions.

(A) Model of the miR-429 regulatory effects on the HIF-1/HIF-3 transition in HUVECs during hypoxia. During normoxia, miR-429 controls steady-state levels of HIF1A and HIF3A mRNA and keeps the levels low. Protein levels remain low through posttranslational modifications and subsequent degradation of these transcription factors when the oxygen levels are normal. During the early stages of hypoxia, HIF1A message and protein are dramatically elevated, while the HIF3A variants and protein remain low. HIF-1α drives a number of pro-survival and angiogenic target genes and also promotes miR-429 and HIF3A up regulation. miR-429 decreases the steady-state levels of both HIF1A and HIF3A mRNA. During prolonged hypoxia, HIF-1α levels and miR-429 are low and HIF3A mRNA and HIF-3α protein accumulate over time, leading to the induction of HIF-3 pro-survival gene expression. (B) The HIF1A and HIF3A relative expression levels (Fragments Per Kilobase of transcript per Million mapped reads) in HUVECs, analysis made base on 2 NGS analysis of 2 independent samples of normoxic HUVEC cells passage 3. Error bars represent SEM.
Methods
Cell lines and culture conditions
HUVECs were obtained from ATCC and maintained until passage six in EGM-2 Bulletkit medium (Lonza). Cells were split either into 6-well plates or 10 cm dishes and allowed to grow to 70–80% confluence prior to the start of the experiments.
Induction of hypoxia
Hypoxia was induced in a CO2/O2 incubator for hypoxia research (Binder CB160 with O2 control, trigas model) and oxygen levels additionally monitored with an independent oxygen meter (PCS GOX100). Briefly, cells were cultured in 10 cm dishes at 1% O2 for the time periods specified. Control cells were maintained in normoxic conditions in the same incubator and harvested at the specified times.
Isolation of RNA and microRNA
Total RNA containing the microRNA fraction was isolated using miRNeasy kit (Qiagen). RNA concentrations were calculated based on the absorbance at 260 nm. RNA samples were stored at −70 °C.
Next generation RNA sequencing analyses (RNA-Seq)
HUVECs (passage 3) were used for the RNA isolation and analyses. Following rRNA depletion, the remaining RNA fraction was used for library construction and subjected to 100 bp paired-end sequencing on an Illumina HiSeq 2000 instrument. Sequencing reads were aligned to the human reference genome assembly (hg19) using TopHat31. Transcript assembly and estimation of the relative abundances were carried out with Cufflinks32. The resulting data were validated with qRT-PCR.
Measurement of mRNA and miRNA levels using quantitative Real Time PCR (qRT-PCR)
We used TaqManOne-Step RT-PCR Master MixReagents (Applied Biosystems) as described previously33,34,35 using the manufacturer’s protocol. The relative expressions were calculated using the comparative relative standard curve method36. We used 18 S rRNA as the relative control for our studies. We also validated this relative control against another housekeeping gene, TATA-binding protein (TBP). As relative controls for miRNA quantification, we validated and used RNU44 and RNU48. TaqMan probes ids used were: 18 S - Hs99999901_s1; TBP - Hs4332659_m1; HIF1A - Hs00153153_m1; RNU44 - 001094; RNU48 - 001006; hsa-miR-429 - 001024, HIF3A - Hs00541711_m1; DDIT4 - Hs01111686_g1. In order to show that miR-429 levels undergo specific changes during hypoxia, we followed the expression of another miR~200 family member miR-141 (hsa -miR-141 - 000463) as shown in Supplemental Figure 5.
miRNA analogs and target protector transfections
miR-429 mimic (id MC10221) and antagomir (id MH10221) were purchased from Ambion. HUVECs were transfected using the Lipofectamine RNAiMax according to manufacturer’s protocol. miR-429 mimic and antagomir were used at final concentrations of 10 nM and 20 nM, respectively. The transfected cells were cultured for 2 days prior to further analysis. The degree of miRNA over-expression or knockdown was determined by qRT-PCR. Target protectorswere purchased from Qiagen and directed against the HIF3A (5′-GTAGAGACGGGGTTTTGCCAAGTTGGAGGGGCTGGTCTTG-3′) and HIF1A (5′-ACATAAATAATAATGCTTTGCCAGCAGTAC-3′) mRNAs. Target protectors were used at final concentration of 600 nM. cel-miR-67 was used as a control (Ambion assay id MC22484). The transfected cells were cultured for 2 days prior to further analysis. As an additional control, Ambion siRNA Negative Control 1 no. 4390843 was used as well.
Western Blots
The procedures were performed as described previously in10,33. The primary antibodies used were: HIF-1α (Abcam ab16066 (1:150); HIF-3α (Sigma-Aldrich, AV39936 (1:800)) and beta actin (Abcam, AB1801 (1:1000). After the washing steps, the membranes were incubated with goat anti-rabbit IgG (H + L) or with goat anti-mouse IgG (H + L) HRP-conjugated secondary antibodies (BioRad) and detected using ECL (Amresco). Since hypoxia affects a number of proteins commonly used as loading controls, finding the proper housekeeping control protein for normalization is problematic37. For example, endothelial cells during hypoxic exposure change their morphology, i.e., tube formation and this affects both beta actin and tubulin levels38,39,40. Furthermore, HIF1 changes affect glucose metabolism and thus GAPDH levels37,39 Prolonged hypoxia inhibits global cellular protein translation and transcription rates41,42. We have tested numerous protein controls for their stability during the time course of hypoxia including beta actin, cofilin, tubulin and GAPDH. Unfortunately, all of them showed variability during the hypoxia time course. Whereas, normalizing the blots to total protein levels allowed us to overcome this limitation. Densitometry was performed using Image Lab software v. 4.1 (BioRad) both prior to the transfer (gel) and after the transfer (membrane). No significant differences in protein distribution were detected between these two methods. The blots shown are representative and the calculations were based on 2 or more independent experiments.
Furthermore, when applicable, all analyzed samples were exposed to hypoxia for the same time period and we included beta actin-based normalization. The potential direct or indirect effects of miR-429 were eliminated since 3′UTR of ACTB does not have a miR-429 binding sequence and more importantly, miR-429 mimic and inhibitor did not affect ACTB levels in next generation sequencing experiments.
Statistical analysis
Results were expressed as means ± standard deviations (SD). Statistical significance among means was determined using the Student’s t-test (two samples, paired and unpaired). Pearson product-moment correlation tests43 were performed with SigmaPlot software.
Additional Information
How to cite this article: Janaszak-Jasiecka, A. et al. miR-429 regulates the transition between Hypoxia-Inducible Factor (HIF)1A and HIF3A expression in human endothelial cells. Sci. Rep. 6, 22775; doi: 10.1038/srep22775 (2016).
References
Hausenloy, D. J. & Yellon, D. M. Myocardial ischemia-reperfusion injury: a neglected therapeutic target. The Journal of clinical investigation 123, 92–100, doi: 10.1172/JCI62874 (2013).
Semenza, G. L. Perspectives on oxygen sensing. Cell 98, 281–284 (1999).
Semenza, G. L. Regulation of mammalian O2 homeostasis by hypoxia-inducible factor 1. Annual review of cell and developmental biology 15, 551–578, doi: 10.1146/annurev.cellbio.15.1.551 (1999).
Majmundar, A. J., Wong, W. J. & Simon, M. C. Hypoxia-inducible factors and the response to hypoxic stress. Molecular cell 40, 294–309, doi: 10.1016/j.molcel.2010.09.022 (2010).
Wang, G. L. & Semenza, G. L. General involvement of hypoxia-inducible factor 1 in transcriptional response to hypoxia. Proceedings of the National Academy of Sciences of the United States of America 90, 4304–4308 (1993).
Kaelin, W. G., Jr. & Ratcliffe, P. J. Oxygen sensing by metazoans: the central role of the HIF hydroxylase pathway. Molecular cell 30, 393–402, doi: 10.1016/j.molcel.2008.04.009 (2008).
Gu, Y. Z., Moran, S. M., Hogenesch, J. B., Wartman, L. & Bradfield, C. A. Molecular characterization and chromosomal localization of a third alpha-class hypoxia inducible factor subunit, HIF3alpha. Gene expression 7, 205–213 (1998).
Maxwell, P. H. et al. The tumour suppressor protein VHL targets hypoxia-inducible factors for oxygen-dependent proteolysis. Nature 399, 271–275, doi: 10.1038/20459 (1999).
Koh, M. Y. & Powis, G. Passing the baton: the HIF switch. Trends Biochem Sci 37, 364–372, doi: 10.1016/j.tibs.2012.06.004 (2012).
Bartoszewska, S. et al. The hypoxia-inducible miR-429 regulates hypoxia-inducible factor-1alpha expression in human endothelial cells through a negative feedback loop. FASEB journal: official publication of the Federation of American Societies for Experimental Biology 29, 1467–1479, doi: 10.1096/fj.14-267054 (2015).
Maynard, M. A. et al. Multiple splice variants of the human HIF-3 alpha locus are targets of the von Hippel-Lindau E3 ubiquitin ligase complex. The Journal of biological chemistry 278, 11032–11040, doi: 10.1074/jbc.M208681200 (2003).
Heikkila, M., Pasanen, A., Kivirikko, K. I. & Myllyharju, J. Roles of the human hypoxia-inducible factor (HIF)-3alpha variants in the hypoxia response. Cellular and molecular life sciences : CMLS 68, 3885–3901, doi: 10.1007/s00018-011-0679-5 (2011).
Pasanen, A. et al. Hypoxia-inducible factor (HIF)-3alpha is subject to extensive alternative splicing in human tissues and cancer cells and is regulated by HIF-1 but not HIF-2. The international journal of biochemistry & cell biology 42, 1189–1200, doi: 10.1016/j.biocel.2010.04.008 (2010).
Ando, H. et al. A hypoxia-inducible factor (HIF)-3alpha splicing variant, HIF-3alpha4 impairs angiogenesis in hypervascular malignant meningiomas with epigenetically silenced HIF-3alpha4. Biochemical and biophysical research communications 433, 139–144, doi: 10.1016/j.bbrc.2013.02.044 (2013).
Maynard, M. A. et al. Human HIF-3alpha4 is a dominant-negative regulator of HIF-1 and is down-regulated in renal cell carcinoma. FASEB journal: official publication of the Federation of American Societies for Experimental Biology 19, 1396–1406, doi: 10.1096/fj.05-3788com (2005).
Zhang, P. et al. Hypoxia-inducible factor 3 is an oxygen-dependent transcription activator and regulates a distinct transcriptional response to hypoxia. Cell reports 6, 1110–1121, doi: 10.1016/j.celrep.2014.02.011 (2014).
Li, Q. F., Wang, X. R., Yang, Y. W. & Lin, H. Hypoxia upregulates hypoxia inducible factor (HIF)-3alpha expression in lung epithelial cells: characterization and comparison with HIF-1alpha. Cell research 16, 548–558, doi: 10.1038/sj.cr.7310072 (2006).
Madanecki, P. et al. Regulation of angiogenesis by hypoxia: the role of microRNA. Cellular & molecular biology letters 18, 47–57, doi: 10.2478/s11658-012-0037-0 (2013).
Greco, S. & Martelli, F. MicroRNAs in Hypoxia Response. Antioxid Redox Signal 21, 1164–1166, doi: 10.1089/ars.2014.6083 (2014).
Greco, S., Gaetano, C. & Martelli, F. HypoxamiR regulation and function in ischemic cardiovascular diseases. Antioxidants & redox signaling 21, 1202–1219, doi: 10.1089/ars.2013.5403 (2014).
Gits, C. M. et al. MicroRNA response to hypoxic stress in soft tissue sarcoma cells: microRNA mediated regulation of HIF3alpha. BMC Cancer 14, 429, doi: 10.1186/1471-2407-14-429 (2014).
Kruger, J. & Rehmsmeier, M. RNAhybrid: microRNA target prediction easy, fast and flexible. Nucleic Acids Res 34, W451–454, doi: 10.1093/nar/gkl243 (2006).
Staton, A. A. & Giraldez, A. J. Use of target protector morpholinos to analyze the physiological roles of specific miRNA-mRNA pairs in vivo. Nature protocols 6, 2035–2049, doi: 10.1038/nprot.2011.423 (2011).
Schneider, A., Younis, R. H. & Gutkind, J. S. Hypoxia-induced energy stress inhibits the mTOR pathway by activating an AMPK/REDD1 signaling axis in head and neck squamous cell carcinoma. Neoplasia 10, 1295–1302 (2008).
Schwarzer, R. et al. REDD1 integrates hypoxia-mediated survival signaling downstream of phosphatidylinositol 3-kinase. Oncogene 24, 1138–1149, doi: 10.1038/sj.onc.1208236 (2005).
Brugarolas, J. et al. Regulation of mTOR function in response to hypoxia by REDD1 and the TSC1/TSC2 tumor suppressor complex. Genes Dev 18, 2893–2904, doi: 10.1101/gad.1256804 (2004).
Piret, J. P., Mottet, D., Raes, M. & Michiels, C. CoCl2, a chemical inducer of hypoxia-inducible factor-1 and hypoxia reduce apoptotic cell death in hepatoma cell line HepG2. Annals of the New York Academy of Sciences 973, 443–447 (2002).
Veschini, L. et al. Hypoxia-inducible transcription factor-1 alpha determines sensitivity of endothelial cells to the proteosome inhibitor bortezomib. Blood 109, 2565–2570, doi: 10.1182/blood-2006-06-032664 (2007).
Rapisarda, A. et al. Schedule-dependent inhibition of hypoxia-inducible factor-1alpha protein accumulation, angiogenesis and tumor growth by topotecan in U251-HRE glioblastoma xenografts. Cancer research 64, 6845–6848, doi: 10.1158/0008-5472.CAN-04-2116 (2004).
Koury, M. J. & Haase, V. H. Anaemia in kidney disease: harnessing hypoxia responses for therapy. Nature reviews. Nephrology 11, 394–410, doi: 10.1038/nrneph.2015.82 (2015).
Trapnell, C., Pachter, L. & Salzberg, S. L. TopHat: discovering splice junctions with RNA-Seq. Bioinformatics 25, 1105–1111, doi: 10.1093/bioinformatics/btp120 (2009).
Trapnell, C. et al. Transcript assembly and quantification by RNA-Seq reveals unannotated transcripts and isoform switching during cell differentiation. Nature biotechnology 28, 511–515, doi: 10.1038/nbt.1621 (2010).
Bartoszewski, R. et al. CFTR expression regulation by the unfolded protein response. Methods in enzymology 491, 3–24, doi: 10.1016/B978-0-12-385928-0.00001-8 (2011).
Bartoszewski, R. et al. Mangiferin has an additive effect on the apoptotic properties of hesperidin in Cyclopia sp. tea extracts. PloS one 9, e92128, doi: 10.1371/journal.pone.0092128 (2014).
Bartoszewski, R. et al. The unfolded protein response (UPR)-activated transcription factor X-box-binding protein 1 (XBP1) induces microRNA-346 expression that targets the human antigen peptide transporter 1 (TAP1) mRNA and governs immune regulatory genes. The Journal of biological chemistry 286, 41862–41870, doi: 10.1074/jbc.M111.304956 (2011).
Larionov, A., Krause, A. & Miller, W. A standard curve based method for relative real time PCR data processing. BMC Bioinformatics 6, 62, doi: 10.1186/1471-2105-6-62 (2005).
Bakhashab, S. et al. Reference Genes for Expression Studies in Hypoxia and Hyperglycemia Models in Human Umbilical Vein Endothelial Cells. G3: Genes|Genomes|Genetics 4, 2159–2165, doi: 10.1534/g3.114.013102 (2014).
Østergaard, L. et al. Proteomics reveals lowering oxygen alters cytoskeletal and endoplasmatic stress proteins in human endothelial cells. PROTEOMICS 9, 4457–4467, doi: 10.1002/pmic.200800130 (2009).
Yang, Y. et al. Effects of hypoxia on mRNA expression of housekeeping genes in rat brain tissue and primary cultured neural cells. Front. Med. China 2, 239–243, doi: 10.1007/s11684-008-0045-7 (2008).
Zhong, H. & Simons, J. W. Direct Comparison of GAPDH, β-Actin, Cyclophilin and 28S rRNA as Internal Standards for Quantifying RNA Levels under Hypoxia. Biochemical and Biophysical Research Communications 259, 523–526, doi: 10.1006/bbrc.1999.0815 (1999).
Liu, L. et al. Hypoxia-Induced Energy Stress Regulates mRNA Translation and Cell Growth. Molecular Cell 21, 521–531, doi: 10.1016/j.molcel.2006.01.010 (2006).
Fähling, M. Surviving hypoxia by modulation of mRNA translation rate. Journal of Cellular and Molecular Medicine 13, 2770–2779, doi: 10.1111/j.1582-4934.2009.00875.x (2009).
Derrick, T. R., Bates, B. T. & Dufek, J. S. Evaluation of time-series data sets using the Pearson product-moment correlation coefficient. Medicine and science in sports and exercise 26, 919–928 (1994).
Acknowledgements
This work has been supported by National Science Center OPUS Program under contract DEC-2011/03/B/NZ3/04387 (to R.B.) and an NIH grant 5R01DK060065 (to J.F.C.).
Author information
Authors and Affiliations
Contributions
Conceived and designed the experiments: R.B. and J.F.C. Performed the experiments: A.J.J., S.B., K.K. and R.B. Analyzed the data: A.R., R.B. and J.F.C. Contributed reagents/materials/analysis tools: R.J.O., L.K., W.K., R.B. and J.F.C. Wrote the paper: R.B. and J.F.C.
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Electronic supplementary material
Rights and permissions
This work is licensed under a Creative Commons Attribution 4.0 International License. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in the credit line; if the material is not included under the Creative Commons license, users will need to obtain permission from the license holder to reproduce the material. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/
About this article

Cite this article
Janaszak-Jasiecka, A., Bartoszewska, S., Kochan, K. et al. miR-429 regulates the transition between Hypoxia-Inducible Factor (HIF)1A and HIF3A expression in human endothelial cells. Sci Rep 6, 22775 (2016). https://doi.org/10.1038/srep22775
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/srep22775
This article is cited by
-
Differential expression of miRNAs associated with pectoral myopathies in young broilers: insights from a comparative transcriptome analysis
BMC Genomics (2024)
-
Methylation of hypoxia-inducible factor 3 subunit alpha contributes to poor prognosis in lung adenocarcinoma
Journal of Applied Genetics (2023)
-
The transition from HIF-1 to HIF-2 during prolonged hypoxia results from reactivation of PHDs and HIF1A mRNA instability
Cellular & Molecular Biology Letters (2022)
-
Unfolded protein response (UPR) integrated signaling networks determine cell fate during hypoxia
Cellular & Molecular Biology Letters (2020)
-
Editorial focus: understanding off-target effects as the key to successful RNAi therapy
Cellular & Molecular Biology Letters (2019)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
