
Abstract
We report here the first complete transcriptome analysis of the dorsal (dDG) and ventral dentate gyrus (vDG) of a rat epilepsy model presenting a hippocampal lesion with a strict resemblance to classical hippocampal sclerosis (HS). We collected the dDG and vDG by laser microdissection 15 days after electrical stimulation and performed high-throughput RNA-sequencing. There were many differentially regulated genes, some of which were specific to either of the two sub-regions in stimulated animals. Gene ontology analysis indicated an enrichment of inflammation-related processes in both sub-regions and of axonal guidance and calcium signaling processes exclusively in the vDG. There was also a differential regulation of genes encoding molecules involved in synaptic function, neural electrical activity and neuropeptides in stimulated rats. The data presented here suggests, in the time point analyzed, a remarkable interaction among several molecular components which takes place in the damaged hippocampi. Furthermore, even though similar mechanisms may function in different regions of the DG, the molecular components involved seem to be region specific.
Similar content being viewed by others


Introduction
Induced, or non-genetic epilepsy animal models, generally rely on the administration of chemoconvulsants or the use of electrical stimulation paradigms1,2,3. Such strategies generally lead to an episode of convulsive status epilepticus (SE), induce variable distribution and intensity of hippocampal neuronal loss, and display significant extra-hippocampal damage4,5,6. Animal models of chronic disorders such as mesial temporal lobe epilepsy (MTLE) provide the unique opportunity to observe the establishment and the development of the pathologic process over time, which is not feasible in patients. Therefore, one would prefer to study animal models displaying a phenotype as similar as possible to that of the human condition. Currently, it is well recognized that a significant proportion of patients with MTLE display the classical hallmarks of hippocampal sclerosis (HS), which is characterized by loss of pyramidal neurons, especially in layers CA1 and CA3 of the hippocampus, and is associated with limited extra-hippocampal lesions7. In 2010, Norwood et al.8 developed an electrical stimulation paradigm in awake rats leading to the induction of hippocampal lesions with a morphology similar to that associated with human HS. More interestingly, these animals develop spontaneous seizures after a latent period of approximately 21 days without the presence of acutely induced generalized tonic clonic SE. To date, this is the only animal model that displays neuronal damage that more closely resembles HS present in patients with MTLE. Even though this animal model is well characterized, the molecular mechanisms that are ultimately responsible for epileptogenesis remain largely undetermined8,9.
Transcriptome profiling allows the broad mapping of molecular constituents present in cells and tissues, resulting in the possible generation of wide-ranging hypotheses about the mechanisms underlying physiological and pathological conditions10. With the advent of massive parallel sequencing of short DNA sequences, it is now possible to estimate the abundance of mRNA molecules in a sample, as well as determine its precise nucleotide sequence10. This strategy has advantages such as: that there is no probe design; and a larger linear range of quantity estimation10,11,12. Furthermore, after acquiring large mRNA data sets it is feasible to use this information to identify signaling pathways and biological processes most affected by a pathological condition by using the Gene Ontology system of classification of enriched genes.
The hippocampus is a highly heterogeneous structure, presenting different patterns of gene expression in different cell populations13,14. Furthermore, it is also possible to observe differences in gene expression even within the same cell population throughout the dorsal-ventral axis of the hippocampus14. Considering these characteristics, the precise isolation of cell populations to preserve spatial information significantly improves the specificity of the information obtained using high-throughput techniques to identify regional differences in molecular mechanisms.
Therefore, the main objective of the present study was to analyze, in one time point during disease progression, the transcriptome profile of the dorsal (dDG) and the ventral dentate gyrus (dDG) of animals with hippocampal lesions induced by Perforant pathway (PP) stimulation, and to explore the molecular mechanisms altered in this animal model.
Results
The stimulation of the PP during electrode placement surgery evoked depolarization of DG granular cells, and electrode positioning was adjusted for maximal amplitude of recorded depolarization. Furthermore, stimulation induced the occurrence of populational spikes (Fig. 1A). After a one-week recovery period, rats were stimulated for 30 minutes on two consecutive days and for 8 hours in the third day as previously described8. The stimulation induced epileptiform activity throughout the stimulation period (Fig. 1B,C). Control rats had electrodes implanted, but were not stimulated after the recovery period. Rats were video monitored after stimulation and did not present spontaneous seizures. Fifteen days after the last stimulation session, rats were euthanized, and brains were collected and processed for laser microdissection. In the present study, we selected the time point, of 15 days after induction of lesion by electrical stimulation because this consists in the ‘silent phase’ of the animal model used. We considered that a time point close to the 8 h electrical stimulation, as for example 48 h after induction, would present gene expression more associated with immediate response to electrical stimulation and apoptosis. A time point later, as for example 30 days would be in a phase were rats would already present spontaneous seizures, that could potentially induce changes in gene expression in the analyzed region due to seizures per se, and not necessarily due to the epileptogenesis process. Therefore, the time point chosen was sufficiently late in relation to the initial stimulation, but before the onset of spontaneous seizures; therefore reflecting molecular processes that most likely are involved in epilptogenesis.

(A) Two evoked local field potentials in the DG after perforant pathway stimulation (40-ms interpulse interval) during electrode placement surgery. (B) Electrical recording of the DG during the perforant pathway stimulation protocol. (C) Detail of discharges observed after a 20-Hz train. (D) Nissl-stained sections used in the laser microdissection procedures. (D) 1 Section from control rat from the Sham group; (D) 2 Same section from control rat after microdissection; (D) 3 Section from an 8-h stimulated rat at 15 days after stimulation. Note extensive lesion in CA1 and CA3. (D) 4 Same section from stimulated rat after microdissection. Even though other hippocampus subfields were microdissected, only the DG was used in the present study. Scale bar = 500 μm. (E) Schematic drawing indicating the anatomical division adopted.
Extensive pyramidal neuronal loss was observed in CA1, CA2 and CA3 in tissue sections from rats that were stimulated when compared to Sham controls (Fig. 1D). The dorsal and ventral regions of the DG granular cell layer were laser microdissected. In Fig. 1E we show a schematic drawing indicating the anatomical division adopted (Fig. 1E).
Total RNA was extracted from each sample, converted into cDNA libraries using Illumina TruSeq Stranded mRNA LT and sequenced in a HISeq® 2500 platform. The sequencing run produced a total of 1,355,974,797 100-bp reads, with 94% of bases over Q30. An average of 84.7 million sequences were produced for each sample. Sequences were aligned with TopHat2 to Rattus norvegicus Ensembl Rnor 5.0 assemble. The average sequence alignment rate was 91.2%. HTSeqCount and DESeq2 packages were used for the estimation of gene expression and statistical analysis. After data processing, a list of differentially expressed genes with p < 0.05 (after correction for multiple tests) was generated. The EdgeR package was used to calculate the Biological coefficient of variance of the present data (BV = 0161), and the RNASeqPower15 package was used to calculate the statistical power of the present experiment, considering the number of biological replicates used. A power greater than 0.9 was found for genes that presented fold-change greater than 1,87 considering a depth of sequence of 20 and the Biological coefficient of variance calculated using the EdgeR package.
We found a total of 2,621 differentially expressed genes (p < 0.05) when comparing control dDG with stimulated dDG: 1,425 genes were up-regulated and 1,196 were down-regulated in stimulated samples. For the vDG we found 2,053 genes differentially expressed when comparing control vDG with stimulated vDG: 1,157 genes were up-regulated and 896 were down-regulated in the stimulated condition (for a complete list refer to supplementary Tables 1 and 2). The dimensionality reduction PCA analysis indicates a clear clustering of samples regarding the condition, control vs. stimulated, and in different regions of the DG, dDG vs. vDG (Fig. 2).

It is possible to observe segregation between control and stimulated groups and between the regions of the dentate gyrus analyzed.
Comparing the effect of stimulation on the dorsal and ventral regions of the DG, there were 1,490 differentially regulated genes in both regions: 590 up-regulated and 900 down-regulated. However, 1,131 genes were uniquely differentially regulated in the dDG: 607 up-regulated and 524 down-regulated. In addition, 563 genes were exclusively differentially regulated in the vDG: 306 up-regulated and 257 down-regulated as shown in Venn diagrams (Fig. 3). Furthermore, when directly comparing the dorsal to ventral portions of the DG from control rats a total of 2,449 genes were differentially expressed between regions, 1,142 up-regulated and 1,307 down-regulated. Comparing the dorsal to ventral portion of the DG from stimulated rats 2,133 genes were differentially expressed, 909 up-regulated and 1,224 down-regulated. The distribution of up and down-regulated genes in the dorsal and ventral regions was found to be homogenous as determined by a chi-square test (p = 0,1845).

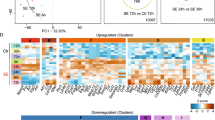
The left circle represents genes differentially expressed in the dorsal dentate gyrus (DG). The right circle represents genes differentially expressed in the ventral dentate gyrus, and the intersection contains differentially expressed genes common to both structures. Tables indicate gene onthologies enriched in each subset of genes represented in the Venn diagram. The X axis is the –log (adjusted p value).
We calculated the enrichment of pathways/biological processes for the set of differentially expressed genes using Metacore® (Thomas-Reuter) (for a complete list refer to supplementary Tables 3 and 4). For the up-regulated genes in the dDG, 67 pathways and 53 biological processes were significantly enriched. Most significant pathways were those involved in immune response such as Inflammosome in inflammatory response, IL-10 signaling, IL-5 signaling, classical complement pathway, and processes such as phagocytosis, inflammosome, interferon signaling and IL-10 inflammatory response. For the down-regulated genes in the dDG, cholesterol biosynthesis was the only highly enriched pathway. This pathway had 16 of its 19 components down-regulated in the dDG of stimulated animals. For the up-regulated genes in the vDG, 45 pathways and 43 biological processes were significantly enriched. In the vDG, there was also a high frequency of immune response-related pathways such as IL-10 and -5 signaling, classical complement activation and processes such as phagocytosis and interferon signaling. For the down-regulated genes in the vDG, cholesterol biosynthesis and regulation of lipid metabolism were the enriched pathways. However, two biological processes showed significant enrichments in the vDG: calcium transport and axonal guidance.
Enrichment pathways/biological process for genes, which were exclusively differentially regulated in stimulated animals were also identified (Fig. 3). For uniquely up-regulated genes in the dDG there were 14 enriched processes, all of which are involved in inflammatory response such as IL-12,15,18 signaling, IL-10 anti-inflammatory response, interferon signaling, IL-6 and leptin signaling. Whereas, for the genes exclusively up-regulated in the vDG, there were six enriched pathways and four biological processes: cell cycle pathways such as spindle assembly and chromosome separation, role of APC in cell cycle regulation, chromosome condensation in prometaphase; cell cycle processes such as mitosis G2-M and spindle microtubules. Also enriched were the cell adhesion – cell matrix interaction process and the role of cell-cell and extracellular matrix interaction in oligodendrocyte differentiation and myelination and MAG-dependent inhibition of neurite outgrowth.
Enriched pathways and biological processes that did not reach statistical significance are shown in Table 1 (P > 0.05); however, they include sets of related genes that were significantly differentially regulated (corrected p values < 0.05 for individual genes).
A total of 18 genes that were identified as differentially regulated in the RNA sequencing experiments were selected for validation by real-time PCR relative quantification (Supplementary figure 1). These genes were selected either because they are components of the major molecular pathways found to be enriched in the transcritpome or because they are genes that were individually mentioned in the discussion. The results from the real-time PCR analysis confirmed the findings obtained by RNA sequencing for all 18 genes tested.
Discussion
The present study explores for the first time the transcriptome of the hippocampus DG during the latent phase of an experimental epilepsy model presenting pathologic findings resembling classical human HS. Furthermore, due to the well-known spatial molecular diversity of the hippocampus14, we separately examined the dorsal and ventral sub-regions of the DG granular cell layer. Therefore, we were able to explore putative molecular mechanisms most likely involved in structural and functional changes that take place in the hippocampus in our model of mesial temporal lobe epilepsy, preserving spatial information about different neuronal circuits.
The most prevalent pathways and biological processes containing differentially expressed genes in both the dorsal and ventral regions of the DG were those related to the immune response. Moreover, the most enriched biological process associated with up-regulated genes in the electrically stimulated rats was phagocytosis. Even though neurons in the DG are considered a resistant cell population in the context of HS, some degree of cell loss within these granular neurons has been observed in humans16 and in animal models17. Interestingly, Norwood et al.8 did not describe any cell loss in the DG in this electrical stimulation model. However, the data presented by these authors did not exclude the possibility of some degree of cell loss because they did not quantify the cells or immunolabel infiltrating macrophage/microglial cells in the DG. Our data indicate a significant increase in expression of genes typically present in immune system cells specialized in phagocytosis, as indicated by the various enriched gene ontologies, such as Inflammosome in inflammatory response pathways or the Phagocytosis process. Furthermore, a typical marker of macrophage/activated microglial cells, CD18 (also known as CR3 or ITGB2)18 showed a 7.6- and 6.4-fold increase in expression in stimulated rats in the dDG and the vDG, respectively. The macrophage/microglia marker Adgre1 (also known as EMR1)19 was also up-regulated in the dDG and vDG. Therefore, our results indicate a greater extent of damage in the dDG than previously reported8. This observation is supported by the immune system-related genes that were exclusively up-regulated in the dDG, such as those in the JAK3/STAT5 pathway, and the morphological characteristics of the tissue observed by Nissl staining (Fig. 2). The observation of exclusive up-regulation in the dDG of JAK3, STAT5, SOCS3 and genes related to NFkβ suggest a greater presence of immune system-related cells, such as lymphocytes infiltrating the dDG20,21. In an intrahippocampal kainic acid injection model in mice, an extensive degeneration of neurons is observed, which is associated with infiltrating microglia, circulating macrophages and lymphocytes in the degenerating hippocampus22. The infiltration of T lymphocytes in the dDG may be responsible for the observed up-regulation in the JAK3/STAT5 pathway, since these immune cells are known to present a higher expression of these genes20,21. However, it is important to point out that although the up-regulation in immune system-related genes may be associated with the infiltration of immune system cells in the DG, it is possible that some of these genes may have been up-regulated in the granule cells. Additional experiments to better characterize the cellular composition of the DG during the latent phase of this electric stimulation epilepsy model, as well as the analysis of gene expression in isolated granule cells, may help to clarify the role of the immune system in the induction of HS and epileptogenesis.
The presence of various differentially regulated genes involved in axonal guidance such as class 3 Semaphorins, Ephrins and Integrins is noteworthy in the present study. Moreover, different members of these families of molecules are uniquely differentially regulated in the dorsal or the ventral portions of the DG. Semaphorins consist of a family of more than 20 secreted and membrane-bound proteins that may function as guidance cues during development and in the adult brain23,24,25. Beyond these established roles, Semaphorins were already shown to be differentially regulated in the hippocampus of experimental epilepsy models26. In the present dataset, we identified a high number of Semaphorins potentially involved in sprouting and abnormal circuitry formation. Furthermore, different Semaphorins (Sema3a and Sema3b, also known as Semaphorins 3A and 3B exclusively up-regulated in the dDG; Sema3c and Sema5a exclusively up-regulated in the vDG; Sema3e exclusively down-regulated in the dDG) were differentially regulated in different portions of the DG, indicating distinct axon guidance mechanisms functioning throughout the dorsal ventral hippocampal axis. Eph receptors and their respective ligands, the Ephrins, are also an important class of cell navigation and axon guidance molecules in the nervous system27. Therefore, these molecules may also be associated with molecular mechanisms underlying epileptogenesis as previously reported28.
An increase in neurogenesis of DG granule cells is a well-known phenomenon associated with epilepsy29. In the present dataset, we observed significantly enriched pathways related to cell proliferation only in the vDG, with the up-regulated genes involved in cell cycle control and mitosis. Furthermore, increased expression of the newborn neuronal marker Dcx (doublecortin) in both the dDG and vDG indicates the presence of migrating newly generated neuroblasts30. These observations may indicate an overall increase in neurogenesis, a phenomenon previously reported in the context of epileptogenesis29. In addition, our gene expression analyses indicated that this proliferation was more intense in the ventral region of the DG. Furthermore, we found an up-regulation of important genes related to myelination, such as the Mag (Myelin associated glycoprotein), Mbp (Myelin basic protein) and Plp1, present only in the vDG. This may indicate a greater extent of sprouting and rewiring taking place in the vDG, leading to an increase in the myelination process.
A common hypothesis for the induction of epileptogenesis is a change in the balance between excitation and inhibition in neural networks. It has been proposed that one possible mechanism for this change in balance would be a preferential loss of GABAergic neurons31,32. Another possibility would be a change in the expression pattern of GABA receptor subunits33,34. In the present dataset, we observed an up-regulation of some GABA A receptor subunits (such as alpha 3 and beta 2) and a reduction of other subunits (such as the delta and rho-3) in both regions of the DG, as well as a down-regulation of the Gabra5 (GABA A receptor alpha-5 subunit) in the dDG. Furthermore, changes in voltage-gated ion channels may also have an important role in the increase of excitability in the context of epileptogenesis. We observed an increase in the expression of Scn4a (Nav1.4), in both the dDG and vDG, a specific increase in Scn9a (Nav1.7) in the dDG and an increase in Scn7a (Nav2.1) in the vDG. The sodium channel isoform Scn4a is typically found in skeletal muscle35, whereas Scn9a is associated with dorsal root ganglia36. Thus, these channels are not usually associated with the hippocampus. The expression of these channel genes was low in the DG compared to other typical hippocampal sodium channels. For example, Scn8a (Nav1.6) had an average of 9,285 counts; whereas, Scn4a had an average of 285 counts and Scn9a an average of 154 counts. Nevertheless, these channels were significantly up-regulated in stimulated animals, suggesting an aberrant expression of sodium channels. Furthermore, their differential regulation may indicate some participation in the epileptogenic process. Voltage-gated potassium channels are a diverse family of genes that also have a key role in the regulation of neuronal excitability37. We found several voltage-gated potassium channel associated-genes differentially regulated in the stimulated animals. For example, expression of the gene Kcnc2 (Kv3.2) was reduced in the dDG. It is worth noting that changes in the expression of such channels may have significant effects on neuronal firing, thus, rendering neurons more susceptible to seizures38.
Neuropeptides are an important class of molecules involved in the regulation of neural tissue excitability39. We observed changes in expression of neuropeptides and receptors, which can be of importance in the context of epilepsy. NPY is a neuropeptide with a significant role in the regulation of food-intake40, and it is associated with anticonvulsant activity41. Furthermore, intra-hippocampal administration of NPY is able to suppress seizures induced by perforant pathway stimulation42. Changes in the expression of NPY have previously been described in other experimental epilepsy models43, and the observed reduction in both the expression of NPY and two of its receptors (Y1 and Y5) in the dDG may play a role in a possible increase in excitability in this region. Interestingly, there was a concomitant reduction in the expression of other neuropeptides that may have an anticonvulsant role, such as Pdyn (also known as prodynorphin or proenkephalin B) a member of the opioid family44, and Sst (somatostatin) exclusively in the vDG45. It is worth noting that Pdyn is also associated with seizure suppression activity in the perforant pathway stimulation model42.
Nervous tissue contains high levels of cholesterol, and because cholesterol carrier proteins do not cross the intact blood brain barrier it is synthesized locally in the brain46. It is known that after a lesion, the level of cholesterol synthesis is reduced in nervous tissue, and cholesterol is recycled from degenerating terminals47. In the hippocampus, for example, damage to the enthorinal cortex leads to extensive degeneration of synaptic input to the DG, resulting in reduction of hippocampal cholesterol synthesis and an increase in the uptake of cholesterol from degenerating terminals via ApoE47. In epilepsy models, a change in cholesterol metabolism due to epilepsy induction was previously observed48. Our results show that the most significantly down-regulated process in both regions of the DG was cholesterol synthesis. Almost all enzymes participating in this process showed reduced expression. These observations indicate the presence of degenerating axons and dendrites in the DG, hence possibly triggering the recycling of cholesterol from such degenerating neuronal projections.
Conclusions
The transcriptome data produced and analyzed in the present study reveal the functional profiles and possible components of the molecular mechanisms that most likely are involved in epileptogenesis in an animal model that displays HS. The gene ontologies enriched in the present dataset implicate several biological processes such as immune-response, the interplay between neural and immune systems, cell death, neural circuitry re-wiring, changes in cell excitability, neurotransmission and metabolic changes. All of these processes are likely to play a role, probably simultaneously, in the induction of epileptogenesis. Of particular interest is the possibility of interplay between the immune system and the granule neurons. This may involve signaling pathways, trophic molecules, cell migration and cell projection guidance molecules, such as those that attract and guide the migration of immune-system cells. These mechanisms may change the migration and projection pattern of neural cells, contributing to the generation of ictogenic tissue.
Many individual processes mentioned above have been previously associated to some degree with epileptogenesis. However, the most important contribution of the present study is that by generating a large and comprehensive dataset, one can more precisely identify specific molecular components of the many biological systems likely contributing to the structural and functional changes that take place in the damaged hipppocampus. Furthermore, the present data also indicate that even though similar mechanisms may be found in different regions of the DG, the components involved seem to be region specific. Finally, our results identified a large number of novel potential targets that may ultimately help inhibit the epileptogenic process.
Methods
Animals
In the present study, three-month-old male Wistar rats (n = 8) were used. Rats were housed under a 12 h/12 h light cycle on a ventilated rack with ad libitum access to standard rodent chow and water. All procedures were performed in accordance with the ethical standards for animal experimentation at the University of Campinas-UNICAMP. The experimental protocol was approved by the UNICAMP animal research ethics committee, which evaluates experimental protocols according to current accepted ethical practices and legislation regarding animal research in Brazil (Brazilian federal law 11.794 (10/08/2008).
Electrode placement surgery and stimulation
Rats were anaesthetized with an isoflurane/oxygen mixture (2%/98%) at 2 L/min using an acrylic induction box. We maintained anesthesia throughout the entire surgical procedure with the aforementioned mixture and administration rate by using a mask adapted to a stereotaxic apparatus. After a deep anesthesia state was verified by the lack of pedal and corneal reflex, we positioned the animal on a stereotaxic apparatus (Leica, Angle Two). The skull was exposed by an incision on the scalp, the Bregma was localized and bone perforations were performed in the following coordinates: +4.5 mm lateral, −7.6 mm posterior; −4.5 mm lateral, −7.6 mm posterior; +2 mm lateral, −3 mm posterior and −2 mm lateral, −3 mm posterior. We performed three perforations in the spaces between the aforementioned coordinates for the fixation of stainless steel screws. One additional perforation was performed 2 mm posterior to the lambdoid suture, in the midline for the placement of a reference electrode (stainless steel, polymide covered, 0.25 mm, Plastics One), the reference electrode was lowered just over the surface of the dura mater.
Bipolar stimulation electrodes (stainless steel, polymide covered, 0.125 mm, Plastics One) with a 1-mm separation were positioned into the perforant pathway (+−4.5 mm lateral, −7.6 mm posterior, −3 mm ventral). Monopolar recording electrodes (stainless steel, polymide covered, 0.25 mm, Plastics One) were positioned into the DG (+−2 mm lateral, −3 mm posterior, −3.5 mm ventral). We optimized the final positioning of the recording and stimulation electrodes based on polarity, occurrence of population spike and maximum amplitude of evoked potentials in the DG (Fig. 1A)49. We performed electrical stimulations with a Grass Astro-Med S88 stimulus generator (paired pulses, 0.1-ms pulse duration, interpulse interval of 40 ms, and pulse amplitude of 20 V). Recordings were performed with a miniature pre-amplifier (ThomasRecording®, 2 channel, 10× gain, dc coupled, 0.06 Hz high-pass filter, 10 kHz low-pass filter) connected to a main amplifier (ThomasRocording®, 8 channel, 25× gain, 0.06 Hz high-pass filter, 10 kHz low-pass filter), and the signal was digitalized at 10 kHz (NIDaq). After electrode and screw positioning, dental acrylic cement was applied in order to hold and stabilize all elements.
Seven days after surgery of electrode placement, freely moving awake rats were stimulated as described previously8 briefly, two days with 30-min stimulation sessions, followed by an 8-h session on the third day. The following stimulation protocol was used during stimulation: continuous 2-Hz paired pulse, 40-ms interpulse interval stimulation, and once per minute a 10-s, 20-Hz single pulse train was delivered using a Grass® Astro-Med S88 stimulator. We recorded DG field potentials throughout the stimulation with the same configuration described for the electrode placement surgery (Fig. 1B,C). Control rats (n = 4) also had surgery for electrode implantation, they were handled and recorded using the same protocols as stimulated rats, but they did not receive any electric stimulation during or after the surgery of electrode placement.
Fifteen days after the last stimulation session, rats were deeply anaesthetized with an isoflurane/oxygen mixture (2%/98%). They were quickly decapitated and the brain was immediately removed and frozen at −60 °C using dry ice and n-hexane.
Laser microdissection
We processed previously frozen brains in a cryostat in order to obtain 60-μm serial sections covering the entire hippocampus. Subsequently, we mounted tissue sections in PEN membrane covered slides (Life Technologies®). These, were immediately stained with Cresyl Violet, dehydrated with an ethanol series and stored at −80 °C. The DG was divided into dorsal and ventral regions according to previously described morphological criteria14. The dDG and the vDG were delimited with a Palm (Zeiss®) system, and tissue was mechanically collected in separate tubes using a surgical microscope and micro-forceps (Fig. 2)
cDNA library preparation and next-generation sequencing
We extracted RNA from microdissected tissue samples with Trizol®, using the manufacturer instructions. cDNA libraries were produced from 200 ng of extracted RNA using the TruSeq Stranded mRNA LT (Illumina®) according to the manufacturer instructions. Each sample had different identifier barcodes, which allowed many samples to be sequenced in the same lane. We sequenced cDNA libraries in a HiSeq® 2500 (Illumina®) in High Output mode, producing 100-bp, single-read sequences. The sequencing run produced a total of 1,355,974,797 100-bp reads, with 94% of bases over Q30. An average of 84.7 million of sequences was produced for each sample. We aligned sequences with TopHat2 (http:// http://ccb.jhu.edu/software/tophat/index.shtml) to Rattus norvegicus Ensembl Rnor 5.0 assemble. The average sequence alignment rate was 91.2%. The HTSeqCount and DESeq2 packages (http://www-huber.embl.de/users/anders /HTSeq/doc/overview.html, as well as http://www.bioconductor.org/ packages/release/bioc/html/DESeq2.html) were used for the estimation of gene expression and statistical analysis. HTSeqCount performs the counting of aligned sequences to genome elements, such as exons and genes. DESeq2 employs a negative-binomial distribution for data normalization, corrects for outliers and the contribution of low expression genes for the analysis of variance, and it uses differential expression statistical Wald tests and corrects for multiple tests employing the Benjamini and Hochberg procedures. After data processing, a list of differentially expressed genes with a statistical significance set at a p < 0.05 (after correction for multiple tests) was generated. We used the differentially expressed genes list for gene ontology analysis with the Metacore® software (Thomas-Reuter).
Gene expression analysis by real time RT-PCR
Total RNA that was previously extracted from microdissected tissue samples (200 ng per sample) was used for cDNA synthesis using Vilo cDNA synthesis kit (Thermo®) following the manufacturer instructions. Relative gene expression was analysed using the 2−ΔctΔct method, for this analysis, primers pairs for the genes: c3, cd18, socs3, mbp, npy, snc4a, sema3a, sema3c, dcx, stat5, adger1, apoe, cdk1, cyp51a1, fdft1, pdyn, sst and y1, were designed using NCBI primer design tool (http://www.ncbi.nlm.nih.gov/tools/primer-blast/). As endogenous controls the geometric mean of the ct of the genes ssr2, ankyf and smntl2 were used. For the PCRs, the Sybr Power Plus Master Mix kit (Thermo®) was used following the manufacturer instructions in a Applied 7500 real-time PCR equipment (Applied Biosystems®). Following PCR amplifications, a melting curve was analysed for each gene, and a single TM peak was observed.
Additional Information
How to cite this article: Vieira, A. S. et al. RNA sequencing reveals region-specific molecular mechanisms associated with epileptogenesis in a model of classical hippocampal sclerosis. Sci. Rep. 6, 22416; doi: 10.1038/srep22416 (2016).
References
Turski, W. A. et al. Limbic seizures produced by pilocarpine in rats: behavioural, electroencephalographic and neuropathological study. Behav Brain Res 9, 315–335 (1983).
Tauck, D. L. & Nadler, J. V. Evidence of functional mossy fiber sprouting in hippocampal formation of kainic acid-treated rats. J Neurosci 5, 1016–1022 (1985).
Kienzler, F., Norwood, B. A. & Sloviter, R. S. Hippocampal injury, atrophy, synaptic reorganization, and epileptogenesis after perforant pathway stimulation-induced status epilepticus in the mouse. J Comp Neurol 515, 181–196, doi: 10.1002/cne.22059 (2009).
Nairismagi, J. et al. Progression of brain damage after status epilepticus and its association with epileptogenesis: a quantitative MRI study in a rat model of temporal lobe epilepsy. Epilepsia 45, 1024–1034, doi: 10.1111/j.0013-9580.2004.08904.x (2004).
Fabene, P. F., Marzola, P., Sbarbati, A. & Bentivoglio, M. Magnetic resonance imaging of changes elicited by status epilepticus in the rat brain: diffusion-weighted and T2-weighted images, regional blood volume maps, and direct correlation with tissue and cell damage. Neuroimage 18, 375–389 (2003).
Bumanglag, A. V. & Sloviter, R. S. Minimal latency to hippocampal epileptogenesis and clinical epilepsy after perforant pathway stimulation-induced status epilepticus in awake rats. J Comp Neurol 510, 561–580, doi: 10.1002/cne.21801 (2008).
Blumcke, I. et al. A new clinico-pathological classification system for mesial temporal sclerosis. Acta Neuropathol 113, 235–244, doi: 10.1007/s00401-006-0187-0 (2007).
Norwood, B. A. et al. Classic hippocampal sclerosis and hippocampal-onset epilepsy produced by a single “cryptic” episode of focal hippocampal excitation in awake rats. J Comp Neurol 518, 3381–3407, doi: 10.1002/cne.22406 (2010).
Will, J. L. et al. Enhanced sequential reaction time task performance in a rat model of mesial temporal lobe epilepsy with classic hippocampal sclerosis. Behav Brain Res 247, 65–72, doi: 10.1016/j.bbr.2013.03.019 (2013).
Wang, Z., Gerstein, M. & Snyder, M. RNA-Seq: a revolutionary tool for transcriptomics. Nat Rev Genet 10, 57–63, doi: 10.1038/nrg2484 (2009).
Bloom, J. S., Khan, Z., Kruglyak, L., Singh, M. & Caudy, A. A. Measuring differential gene expression by short read sequencing: quantitative comparison to 2-channel gene expression microarrays. BMC Genomics 10, 221, doi: 10.1186/1471-2164-10-221 (2009).
Denoeud, F. et al. Annotating genomes with massive-scale RNA sequencing. Genome Biol 9, R175, doi: 10.1186/gb-2008-9-12-r175 (2008).
Greene, J. G., Borges, K. & Dingledine, R. Quantitative transcriptional neuroanatomy of the rat hippocampus: evidence for wide-ranging, pathway-specific heterogeneity among three principal cell layers. Hippocampus 19, 253–264, doi: 10.1002/hipo.20502 (2009).
Fanselow, M. S. & Dong, H. W. Are the dorsal and ventral hippocampus functionally distinct structures? Neuron 65, 7–19, doi: 10.1016/j.neuron.2009.11.031 (2010).
Hart, S. N., Therneau, T. M., Zhang, Y., Poland, G. A. & Kocher, J. P. Calculating sample size estimates for RNA sequencing data. J Comput Biol 20, 970–978, doi: 10.1089/cmb.2012.0283 (2013).
Blumcke, I. et al. Towards a clinico-pathological classification of granule cell dispersion in human mesial temporal lobe epilepsies. Acta Neuropathol 117, 535–544, doi: 10.1007/s00401-009-0512-5 (2009).
Zhang, S., Khanna, S. & Tang, F. R. Patterns of hippocampal neuronal loss and axon reorganization of the dentate gyrus in the mouse pilocarpine model of temporal lobe epilepsy. J Neurosci Res 87, 1135–1149, doi: 10.1002/jnr.21941 (2009).
Kato, H., Kogure, K., Liu, X. H., Araki, T. & Itoyama, Y. Progressive expression of immunomolecules on activated microglia and invading leukocytes following focal cerebral ischemia in the rat. Brain Res 734, 203–212 (1996).
Eglitis, M. A. & Mezey, E. Hematopoietic cells differentiate into both microglia and macroglia in the brains of adult mice. Proc Natl Acad Sci USA 94, 4080–4085 (1997).
Liva, S. M., Kahn, M. A., Dopp, J. M. & de Vellis, J. Signal transduction pathways induced by GM-CSF in microglia: significance in the control of proliferation. Glia 26, 344–352 (1999).
Musso, T. et al. Regulation of JAK3 expression in human monocytes: phosphorylation in response to interleukins 2, 4, and 7. J Exp Med 181, 1425–1431 (1995).
Zattoni, M. et al. Brain infiltration of leukocytes contributes to the pathophysiology of temporal lobe epilepsy. J Neurosci 31, 4037–4050, doi: 10.1523/JNEUROSCI.6210-10.2011 (2011).
Mann, F., Chauvet, S. & Rougon, G. Semaphorins in development and adult brain: Implication for neurological diseases. Prog Neurobiol 82, 57–79, doi: 10.1016/j.pneurobio.2007.02.011 (2007).
Skaliora, I., Singer, W., Betz, H. & Puschel, A. W. Differential patterns of semaphorin expression in the developing rat brain. Eur J Neurosci 10, 1215–1229 (1998).
Sahay, A. et al. Secreted semaphorins modulate synaptic transmission in the adult hippocampus. J Neurosci 25, 3613–3620, doi: 10.1523/JNEUROSCI.5255-04.2005 (2005).
Holtmaat, A. J. et al. Transient downregulation of Sema3A mRNA in a rat model for temporal lobe epilepsy. A novel molecular event potentially contributing to mossy fiber sprouting. Exp Neurol 182, 142–150 (2003).
Goldshmit, Y., McLenachan, S. & Turnley, A. Roles of Eph receptors and ephrins in the normal and damaged adult CNS. Brain Res Rev 52, 327–345, doi: 10.1016/j.brainresrev.2006.04.006 (2006).
Xu, B. et al. EphA/ephrin-A interactions regulate epileptogenesis and activity-dependent axonal sprouting in adult rats. Mol Cell Neurosci 24, 984–999 (2003).
Parent, J. M. et al. Dentate granule cell neurogenesis is increased by seizures and contributes to aberrant network reorganization in the adult rat hippocampus. J Neurosci 17, 3727–3738 (1997).
Brown, J. P. et al. Transient expression of doublecortin during adult neurogenesis. J Comp Neurol 467, 1–10, doi: 10.1002/cne.10874 (2003).
Andre, V., Marescaux, C., Nehlig, A. & Fritschy, J. M. Alterations of hippocampal GAbaergic system contribute to development of spontaneous recurrent seizures in the rat lithium-pilocarpine model of temporal lobe epilepsy. Hippocampus 11, 452–468, doi: 10.1002/hipo.1060 (2001).
Dinocourt, C., Petanjek, Z., Freund, T. F., Ben-Ari, Y. & Esclapez, M. Loss of interneurons innervating pyramidal cell dendrites and axon initial segments in the CA1 region of the hippocampus following pilocarpine-induced seizures. J Comp Neurol 459, 407–425, doi: 10.1002/cne.10622 (2003).
Houser, C. R. & Esclapez, M. Downregulation of the alpha5 subunit of the GABA(A) receptor in the pilocarpine model of temporal lobe epilepsy. Hippocampus 13, 633–645, doi: 10.1002/hipo.10108 (2003).
Peng, Z., Huang, C. S., Stell, B. M., Mody, I. & Houser, C. R. Altered expression of the delta subunit of the GABAA receptor in a mouse model of temporal lobe epilepsy. J Neurosci 24, 8629–8639, doi: 10.1523/JNEUROSCI.2877-04.2004 (2004).
Wang, J. Z. et al. Sequence and genomic structure of the human adult skeletal muscle sodium channel alpha subunit gene on 17q. Biochem Biophys Res Commun 182, 794–801 (1992).
Sangameswaran, L. et al. A novel tetrodotoxin-sensitive, voltage-gated sodium channel expressed in rat and human dorsal root ganglia. J Biol Chem 272, 14805–14809 (1997).
Sandhiya, S. & Dkhar, S. A. Potassium channels in health, disease & development of channel modulators. Indian J Med Res 129, 223–232 (2009).
Lau, D. et al. Impaired fast-spiking, suppressed cortical inhibition, and increased susceptibility to seizures in mice lacking Kv3.2 K+channel proteins. J Neurosci 20, 9071–9085 (2000).
Baraban, S. C. & Tallent, M. K. Interneuron Diversity series: Interneuronal neuropeptides–endogenous regulators of neuronal excitability. Trends Neurosci 27, 135–142, doi: 10.1016/j.tins.2004.01.008 (2004).
Stanley, B. G., Magdalin, W., Seirafi, A., Nguyen, M. M. & Leibowitz, S. F. Evidence for neuropeptide Y mediation of eating produced by food deprivation and for a variant of the Y1 receptor mediating this peptide’s effect. Peptides 13, 581–587 (1992).
Baraban, S. C. & Neuropeptide, Y. and epilepsy: recent progress, prospects and controversies. Neuropeptides 38, 261–265, doi: 10.1016/j.npep.2004.04.006 (2004).
Mazarati, A. & Wasterlain, C. G. Anticonvulsant effects of four neuropeptides in the rat hippocampus during self-sustaining status epilepticus. Neurosci Lett 331, 123–127 (2002).
Lurton, D. & Cavalheiro, E. A. Neuropeptide-Y immunoreactivity in the pilocarpine model of temporal lobe epilepsy. Exp Brain Res 116, 186–190 (1997).
Loacker, S., Sayyah, M., Wittmann, W., Herzog, H. & Schwarzer, C. Endogenous dynorphin in epileptogenesis and epilepsy: anticonvulsant net effect via kappa opioid receptors. Brain 130, 1017–1028, doi: 10.1093/brain/awl384 (2007).
Sloviter, R. S. Decreased hippocampal inhibition and a selective loss of interneurons in experimental epilepsy. Science 235, 73–76 (1987).
Dietschy, J. M. & Turley, S. D. Cholesterol metabolism in the brain. Curr Opin Lipidol 12, 105–112 (2001).
Poirier, J., Baccichet, A., Dea, D. & Gauthier, S. Cholesterol synthesis and lipoprotein reuptake during synaptic remodelling in hippocampus in adult rats. Neuroscience 55, 81–90 (1993).
de Freitas, R. M., do Nascimento, K. G., Ferreira, P. M. & Jordan, J. Neurochemical changes on oxidative stress in rat hippocampus during acute phase of pilocarpine-induced seizures. Pharmacol Biochem Behav 94, 341–345, doi: 10.1016/j.pbb.2009.09.015 (2010).
Andersen, P., Holmqvist, B. & Voorhoeve, P. E. Entorhinal activation of dentate granule cells. Acta Physiol Scand 66, 448–460, doi: 10.1111/j.1748-1716.1966.tb03223.x (1966).
Acknowledgements
This work was supported by grants from Fundação de Amparo à Pesquisa do Estado de São Paulo (FAPESP), SP, BRAZIL. Dr. ASV is supported by a fellowship from FAPESP; ABM and AC are supported by studentships from FAPESP; Dr. CSR is supported by a fellowship from Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES), BRAZIL; Dr. BSC is supported by a fellowship from FAPESP, and Drs. F. Cendes and IL-C are supported by grants from FAPESP and Conselho Nacional de Pesquisa (CNPq), Brazil.
Author information
Authors and Affiliations
Contributions
I.L.C., F.C., V.D.B.P. and A.S.V. conceived the study, designed the experiments and drafted the manuscript. A.S.V. performed electrode placement surgeries and electrical stimulation/recordings. A.S.V., A.M. and A.C. performed, tissue collection, laser microdissection, analysed video recordings from rats and performed library preparation. A.S.V., C.S.R. and B.C. performed bioinformatics analysis of Rna-seq data. B.C., B.A.N., S.B., F.R. and R.G. offered advice on data analysis and manuscript preparation. I.L.C. supervised the project. All authors discussed and commented on the manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Supplementary information
Rights and permissions
This work is licensed under a Creative Commons Attribution 4.0 International License. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in the credit line; if the material is not included under the Creative Commons license, users will need to obtain permission from the license holder to reproduce the material. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/
About this article

Cite this article
Vieira, A., de Matos, A., do Canto, A. et al. RNA sequencing reveals region-specific molecular mechanisms associated with epileptogenesis in a model of classical hippocampal sclerosis. Sci Rep 6, 22416 (2016). https://doi.org/10.1038/srep22416
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/srep22416
This article is cited by
-
Transcriptome analysis of l-leucine-producing Corynebacterium glutamicum under the addition of trimethylglycine
Amino Acids (2022)
-
Laser microdissection-based microproteomics of the hippocampus of a rat epilepsy model reveals regional differences in protein abundances
Scientific Reports (2020)
-
Epileptogenese und Konsequenzen für die Therapie
Der Nervenarzt (2019)
-
New Insights on Temporal Lobe Epilepsy Based on Plasticity-Related Network Changes and High-Order Statistics
Molecular Neurobiology (2017)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
