
Abstract
Diel hysteresis occurs often between soil CO2 efflux (RS) and temperature, yet, little is known if diel hysteresis occurs in the two components of RS, i.e., autotrophic respiration (RA) and heterotrophic respiration (RH) and how diel hysteresis will respond to future rainfall change. We conducted a field experiment in a desert ecosystem in northern China simulating five different scenarios of future rain regimes. Diel variations of soil CO2 efflux and soil temperature were measured on Day 6 and Day 16 following the rain addition treatments each month during the growing season. We found contrasting responses in the diel hysteresis of RA and RH to soil temperature, with a clockwise hysteresis loop for RH but a counter-clockwise hysteresis loop for RA. Rain addition significantly increased the magnitude of diel hysteresis for both RH and RA on Day 6, but had no influence on either on Day 16 when soil moisture was much lower. These findings underline the different roles of biological (i.e. plant and microbial activities) and physical-chemical (e.g. heat transport and inorganic CO2 exchange) processes in regulating the diel hysteresis of RA and RH, which should be considered when estimating soil CO2 efflux in desert regions under future rainfall regime.
Similar content being viewed by others


Introduction
Soil CO2 efflux, the second largest terrestrial carbon flux following photosynthesis, has received widespread attention during the past decades due to its vulnerability and high sensitivity to climate change1,2,3. On a diel, seasonal and annual scale, soil temperature is widely recognized as a primary abiotic factor affecting soil CO2 efflux and the relationship between soil CO2 efflux and soil temperature is often expressed as Arrhenius or van’t Hoff exponential functions4,5,6. However, describing and predicting soil CO2 efflux using these simple reaction functions lead to many uncertainties, because various biotic and abiotic factors, such as plant physiological activity, soil physical properties and soil moisture4,7,8,9, can also considerably influence the relationship.
In recent years, many field studies have shown that, on the diel and seasonal scales, soil CO2 efflux and soil temperature are decoupled, showing an elliptical hysteresis loop10,11. For example, soil CO2 efflux may increase more quickly in response to rising soil temperature in the morning compared to decreasing soil temperature in the evening, resulting in a clockwise diel hysteresis. Diel hysteresis (in some references also as “diurnal hysteresis”) has been observed in the soils of forests12,13,14,15, savanna grasslands16,17 and crop fields18,19,20. The mechanistic explanation for such hysteresis phenomenon of soil CO2 efflux is still lacking, although two possible hypotheses have emerged. The first one focuses on the biological mechanism associated with photosynthate transport such that recent photosynthates are periodically transported to the roots zone, where they can alter the magnitude and timing of soil CO2 efflux, leading to the phenomenon that soil CO2 efflux is out of sync with soil temperature13,15,21. For example, by comparing the relationship between soil CO2 efflux and soil temperature under tree canopies and in an open area, Vargas and Allen13 proposed that photosynthesis is the main contributor to the diel hysteresis phenomenon of soil CO2 efflux. The second hypothesis relies on the physical mechanisms associated with heat transport and CO2 diffusion processes. The direction of heat transport is opposite in daytime to that in nighttime, which can influence the velocities of CO2 diffusion along soil profiles when soil warms and cools10,15. The biological and physical mechanisms are not mutually exclusive and both are likely to play important roles in formulating diel hysteresis of soil CO2 efflux10. In addition, many researchers have suggested that without considering diel hysteresis, a large bias will likely be present in the estimation of soil CO2 efflux and soil C loss15,17,22. In a montane conifer forest, for example, Riveros-Iregui et al.15 found that a representative model without considering diel hysteresis could overestimate the total soil CO2 efflux of the growing season by 19%. Thus, proper interpretation and evaluation of diel hysteresis of soil CO2 efflux is critical to accurate estimation and prediction of regional and global C budgets.
Soil rewetting following a rainfall event has been shown to be an important controlling factor for regulating soil CO2 efflux, especially in desert and semiarid regions8,23,24. Global climate models predict that the desert and semiarid regions such as those in northern China will undergo more extreme climate changes characterized by increasing amount of total precipitation and higher frequency of extreme rainfall events in the 21st century25,26,27. Given that water is the primary driver of biological activity in desert and semiarid areas28, such alterations of the hydrological cycle will potentially alter soil CO2 efflux. In addition, by changing soil moisture, precipitation increase can influence heat transport and gas diffusion processes, since diffusion of gas in the gas phase is much higher than in the liquid phase29. Therefore, any change in precipitation regions under the climate change scenarios can potentially influence diel hysteresis of soil CO2 efflux. Thus, it is very important to evaluate possible responses of diel hysteresis of soil CO2 efflux to changes in soil water availability associated with rainfall events, as any change in diel hysteresis of soil CO2 efflux may affect the evaluation of possible response of soil CO2 efflux to future precipitation change.
Soil CO2 efflux has two components: the autotrophic respiration (RA) of roots and the associated rhizosphere community and the heterotrophic respiration (RH) from the decomposition processes of soil microbes9,30. However, most previous field studies mainly focused on diel hysteresis between total soil CO2 efflux and soil temperature12,13,14,15,16,17,19. Research concerning the diel hysteresis of the two respiration components is still rare18 and represents a gap in our knowledge of the response of soil CO2 efflux to temperature. In addition, we are not aware of any study conducted to distinguish the differential response of diel hysteresis between the two components of soil CO2 efflux in situ during rain pulses over an entire growing season. Modeling studies have shown differential responses of RA and RH to climate change such as increasing temperature31, indicating that the responses would be accompanied by varying changes in the magnitude of diel hysteresis of the two respiration components. Thus, partitioning soil CO2 efflux into its two components and analyzing the different responses of diel hysteresis are urgently needed for more accurate modeling of regional and global C budgets under future climate change scenarios.
We conducted a manipulative field experiment to simulate five scenarios of future rain regimes (0%, 25%, 50%, 75% and 100% increase over local annual mean precipitation (115 mm)) in a desert ecosystem of northwestern China dominated by the shrub species Nitraria tangutorum. The rain addition treatments were applied equally each month during the growing season (May-September). The effects of rain addition amount and timing on soil CO2 efflux in this arid ecosystem have been reported previously8,32. Although this study used some of the same dataset as our previous study32, the current study focused on the diel hysteresis phenomenon of two soil respiration components and their differential responses to rain additions, which is quite different from our previous paper in term of the scientific questions we addressed and the methods we used to analyze the data. The main objectives of this paper were to evaluate: (1) the diel relationship of the two components of soil CO2 efflux with soil temperature and (2) the possible responses of the diel hysteresis of the two components to rain increase. We hypothesized that the responses of diel hysteresis to rain addition would differ between the two components of soil CO2 efflux because roots and microbes often have different response mechanisms to rain pulses of varying size, considering that small rainfall events can only enhance microbial respiration and larger rainfall events can increase both microbial and root respiration with different response time and duration24,32,33.
Materials and Methods
Study site description
The experimental site (38° 34′ N, 102° 58′ E) was located in a sandy land area between Badain Jaran Desert and Tengger Desert in Minqin County, Gansu province, China. This area has a temperate arid continental climate with a mean annual temperature of 7.8 °C. The temperature during the warmest and coldest months is 23.2 °C and −9.6 °C, respectively. Mean annual precipitation is 115 mm mainly from July to September. The dominant soil type is aeolian sandy soil (Entisols in the USDA soil taxonomy system) with pH about 8.6. The dominant species is Nitraria tangutorum Bobr. More detailed information on the physical and chemical properties of soil can be found in Song et al.8.
Experimental design
The experiment was carried out in a patchy landscape with N. tangutorum interspersed with sand dunes, oriented in a northwest-southeast direction and composed of two distinct types of soil cover. The northwest area was covered with N. tangutorum plants, while the southeast area was bare soil. The mean height of the dunes was 0.9 m and the area was 14.2 m2. Vegetation cover was approximately 35% at the entire study site.
A completely random design was used in this experiment, with five precipitation treatments and four replicates for each treatment (113 m2 per plot, 20 plots in total). During the growing season (from May to September) of 2009, five precipitation treatments were designed to simulate a rain increase of 0% (CK), 25% (+25%PPT), 50% (+50%PPT), 75% (+75%PPT) and 100% (+100%PPT) over long-term average annual precipitation at the study site (115 mm). The precipitation addition treatments were applied every month and the precipitation amount added each time was 0, 5.8, 11.5, 17.3 and 23.0 mm for the five addition treatments, respectively. Water was pumped into a tank from a well near the plots and used to irrigate the plots via an irrigation system composed of a water-pump, water meter and spraying arms. In order to reduce water evaporation, precipitation treatments were carried out only in the morning. More detailed information on this experiment is provided in Song et al.8.
Soil CO2 efflux measurements
Soil CO2 efflux was measured on Day 6 and Day 16 after each rain addition for each month during the growing season (May to September). According to the general model of ecosystem carbon exchange following rain pulses24,33,34, shrubs in desert ecosystems are generally characterized by long lag time and long duration relative to microbes in responding to an effective rainfall. Huang and Nobel35 found that desert plants initialized new root growth c. 7 days after rewetting. The rate of soil CO2 efflux of vegetated soils in water irrigated plots showed a significant difference from that in control plot only about 1 week after the water addition treatment8. In addition, the soil moisture data also showed that the effects of the rain addition treatments could last as long as 15 days after the rain addition treatment8. Thus, we chose Days 6 and 16 after the rain addition treatment for the soil CO2 efflux measurements in this study.
At the beginning of the experiment, two PVC soil collars (80 cm2 in area and 5 cm in height) were installed into the vegetated and bare soils to a depth of 3 cm in each plot for measuring soil CO2 efflux. The rate of soil CO2 efflux was determined using an automated soil CO2 flux system (LI-8100, LI-COR Inc., Lincoln, NE, USA). Soil CO2 efflux was measured on Days 6 and 16 after rain addition each month during the growing season (May to September). On each measurement day, soil CO2 efflux rate was measured on each pre-installed collar repeatedly at an interval of 3 h from 6:00 AM to 6:00 AM of the following day. To achieve a more accurate estimate of soil CO2 efflux, we visited the 20 plots in the same sequence on every occasion that measurements were taken. As the rate of soil CO2 efflux was being measured, soil temperature (Ts) was measured at a depth of 10 cm with a thermocouple connected to the LI-8100 system. On each given measurement day, we also measured daily soil gravimetric water content (SWC) of the top10 cm of the soil column in all plots by the oven drying method. We chose to measure soil temperature at 10 cm depth based on the compromise that most microbes were concentrated in the upper layers (0–10 cm). However, most roots biomass was found in the 10–30 cm soil layers.
Partition of soil CO2 efflux components
At the study site, no obvious root biomass existed in the bare soils and grasses and forbs were scarce enough to be negligible. Thus, the soil CO2 efflux in the bare soils would mainly come from the decomposition of soil organic carbon - microbial respiration or heterotrophic respiration (RH). There were no differences in soil organic matter, soil nitrogen content or microbial biomass between the two vegetation cover types8. In addition, there were no differences in soil temperature or moisture between the two vegetation cover types during the experimental period. Thus, autotrophic respiration (RA) could roughly be determined by the difference in soil CO2 efflux measured in the vegetated and bare soils:

where RS is soil CO2 efflux in the vegetated soil; RH is soil CO2 efflux in the bare soil; and RA is autotrophic respiration.
Quantification of the diel hysteresis for soil CO2 efflux
For each measurement day of the growing season, we fitted the relationship between soil CO2 efflux and soil temperature during the night time (21:00–06:00) using a van’t Hoff exponential function. The parameters of the function were then adopted to calculate soil CO2 efflux during the daytime (09:00–18:00). Diel hysteresis was determined by the maximum difference of the daytime soil CO2 efflux measured and calculated.

where the degree of diel hysteresis (DH) is the maximum difference between the soil CO2 efflux measured (RSm) during the daytime and the efflux calculated by the exponential function (RSc) that was extracted from the relationship between the nighttime soil CO2 efflux and soil temperature. In this study, the maximum difference occurred at 12:00 for each measurement day. Positive values of diel hysteresis represent clockwise loop (higher CO2 efflux during soil warming period).
Statistical analyses
We performed the repeated measurement analysis of variance (ANOVA) to test the differences in the degree of diel hysteresis of RS, RH and RA on Days 6 and 16 after different rain addition treatments. Two-way ANOVA tests were used to examine the effect of rain addition time and rain addition amount on diel hysteresis of RS, RH and RA on Days 6 and 16 after rain addition. Linear regression analysis was conducted to examine possible relationships between diel hysteresis and daily soil moisture. All statistical analyses were performed using SPSS 17.0 (SPSS for Windows, Version 17.0, Chicago, IL, USA).
Results
Soil water content and temperature under different rain additions
We measured the surface (0–10 cm) soil water content (SWC) on Days 6 and 16 following the rain addition treatments each month. As expected, SWC was strongly affected by rain addition treatments in both vegetated and bare soils (Fig. 1). The magnitude of change in SWC depended on the amount of rain added and the measurement day. Day 6 usually showed a higher increase in SWC than Day 16 during the growing season except for August due to a heavy natural rainfall event on 25thAugust (25.8 mm). There was no significant difference in SWC between the vegetated and bare soils under a given rain addition treatment on each measurement day32.

(a) Daily precipitation during the experimental period from 1 May to 1 October. (b,c): Soil water content (SWC) at 10 cm depth in vegetated soil and bare soil of a N tangutorum dominated desert ecosystem. (d) Daily mean air temperature (Ta) during the experimental period. (e,f): Soil temperature at 10 cm depth (Ts) in vegetated and bare soils. Long solid arrows represent the timing of rain addition treatments; short solid and dashed arrows represent the measurement time on Day 6 and Day 16 after the rain addition treatments, respectively.
The seasonal trend in daily mean surface (0–10 cm) soil temperature (Ts) was similar for both vegetation cover types during the entire growing season (Fig. 1). Daily mean Ts at each measurement day did not show any significant differences among the five rain addition treatments32. Moreover, there was no difference in daily mean Ts between the vegetated and bare soils under a given rain addition treatment on each measurement day32.
Relationship between diel soil CO2 efflux and soil temperature
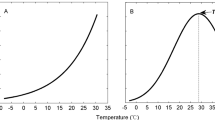
Plots of the diel dynamics of RS, RH and RA against soil temperature on Day 6 and Day 16 following the rain addition treatments are shown in Fig. 2. A clockwise diel pattern of hysteresis with respect to soil temperature was found for RS and RH on both measurement days, with higher rate of RS and RH when soil temperature was increasing during daytime and lower rate when temperature was decreasing during nighttime (Fig. 2a,b,d,e). The rate of RS and RH reached the maximum value between 12:00–15:00 and then gradually decreased to minimum value between 3:00–6:00. However, the maximum rate of RA occurred between 18:00–21:00 and the minimum occurred between 6:00–9:00 despite the increase in soil temperature (Fig. 2c,f). Diel rate of RA exhibited a counter-clockwise effect with soil temperature, that is, for a given temperature, the rate of RA at nighttime was higher than at daytime.

Relationships between diel soil CO2 efflux and soil temperature at 10 cm soil depth on Days 6 and 16 after the rain addition treatments during the growing season.
(a,d) Soil CO2 efflux in the vegetated soils (total soil CO2 efflux, RS), (b,e) soil CO2 efflux in the bare soils (heterotrophic respiration, RH), (c,f) calculated autotrophic respiration (RA). Dotted arrows indicate the directions of the hysteresis loop.
Effect of rain addition treatments on diel hysteresis of soil CO2 efflux
During the growing season, the diel hysteresis of RS and RH was positive while the diel hysteresis of RA was negative (Fig. 3). The values of diel hysteresis of RS and RH showed significant seasonal variations, with the minimum diel hysteresis of RS and RH occurring on June 26th and the maximum on August 26th (Fig. 3a,b). However, the diel hysteresis of RA were relatively stable during the growing season (Fig. 3c).

Seasonal variations in the diel hysteresis of soil CO2 efflux (RS) and its components (RH and RA).
Long solid arrows represent the timing of rain addition treatments; short solid and dashed arrows represent the measurement time on Day 6 and Day 16 after the rain addition treatments, respectively.
On Day 6, the rain addition had no influence on the diel hysteresis of RS (Fig. 4a), but significantly increased the amplitudes of diel hysteresis of the two respiration components (Fig. 4b,c). On Day 16, the rain addition had no significant effect on the diel hysteresis of RS and its components (Fig. 4d-f). There was no significant interaction effect of rain addition amount and rain addition time on the diel hysteresis of RA and RH on either Day 6 or 16 (Table S1). The value of diel hysteresis for RH was always higher than for RA on both measurement days. In addition, the amplitude of diel hysteresis positively corresponded with he increase in daily SWC for RS and its components on both measurement days (Fig. 5).

Mean values of diel hysteresis of soil CO2 efflux (RS) and its components (RH and RA) on Day 6 and Day 16 after different rain addition treatments during the growing season in 2009 (n = 4).
The different letters indicate significant differences between rain addition treatments at P < 0.05.

Relationships between absolute values of diel hysteresis and daily soil water content (SWC).
The method of using nighttime soil CO2 efflux data to predict daytime soil CO2 efflux, a common method in eddy covariance studies, resulted in significant error in the estimation of daily total soil CO2 efflux (Table 1). During the growing season, there was an underestimation of daily flux by 10–32% for RS and 41–62% for RH, but an overestimation of RA by 5–12% in this desert ecosystem. Rain addition changed the value of the error and the magnitude of the change depended on the date of measurement (Table 1). On Day 6, rain addition significantly decreased the magnitude of the error for RS and RH but had no influence for RA. On Day 16, however, there was a significant decrease in the magnitude of error for RS but had no change in the two respiration components.
Discussion
Numerous studies have investigated diel hysteresis between soil CO2 efflux and temperature, but reached contrasting conclusions on the mechanisms for the occurrence of such phenomenon12,13,14,15,16,17,18,19,20,36. In addition, these results mainly focused on diel hysteresis of total soil CO2 efflux and so far research concerning the diel hysteresis of two respiration components is still rare18. Furthermore, how diel hysteresis will respond to climate change, particularly precipitation change, is still poorly understood. To best of our knowledge, this is the first study focused on the differential responses of the diel hysteresis between the two components of soil CO2 efflux in situ to rain pulses over an entire growing season. Our results from this field manipulation experiment offer new insights into how soil CO2 efflux responds to temperature and soil moisture.
In this study, we found that a distinct diel hysteresis existed between RA and RH in terms of their responses to soil temperature. The rate of RA was higher when soil temperature was decreasing than when soil temperature was increasing, producing a counter-clockwise hysteresis loop. In contrast, the rate of RH increased more quickly in response to rising soil temperature in the morning compared to decreasing soil temperature in the evening, showing a clockwise hysteresis loop. These results suggest that both biological and physical mechanisms should be responsible for the diel hysteresis of soil CO2 efflux in this desert ecosystem and also indicate that variables that control diel behaviour of RA in response to changes in soil temperature are different from the variables that control diel behaviour of RH.
Several field studies have found a strong connection between plant photosynthesis and RA at diel and seasonal scales16,21,37. The time required to transport photosynthetic C to roots was found on the order of many hours to a few day10,13,15,38. For instance, Tang et al.16 reported that the time for photosynthetic products to be transported to the roots and be respired is less than 1 day. Similarly, by in situ13CO2 pulse labelling of shrubs, Carbone and Trunbore39 demonstrated that a part of photosynthetic C can be rapidly transported to belowground and be quickly metabolized by roots within a few hours. In this study, peak RA occurred at between 18:00–21:00, which was about 3 h after peak 10 cm soil temperature, 6 h after peak PAR. The high correlation between daily mean PAR and daily mean RA (Fig. S1) suggested that the C respired by roots may be related to the same day’s photosynthesis. We speculate that accumulated photosynthetic C during daytime was periodically transported to the roots and metabolized at night, which could have contributed to the counter-clockwise diel hysteresis of RA (Fig. 6a). However, in this study site, most roots biomass was concentrated in the 10–30 cm soil layers, which suggested that the diel hysteresis of RA could have also been modified by physical processes. According to model analysis, Phillips et al.10 demonstrated that the effect of CO2 diffusivity on diel hysteresis can be bypassed because it does little to shift diel oscillation of RA considering the high porosity of sandy soil, which is similar to the soil type in this study. In contrast to CO2 diffusivity, model based on physical first principles (Table 1 and Fig. 3 in Phillips et al.10) proved that thermal transport plays an important role in controlling the diel hysteresis of soil CO2 efflux by controlling the speed of air temperature propagated through soil and by changing the synchronicity of diel variations of soil temperature and efflux10. The asynchronous diel patterns between soil temperature and efflux could result in a clockwise hysteresis loop within an approximate rate (e.g. 1 × 10−7 m2 s−1 based on measurements of a sandy loam soil) of thermal diffusivity for sandy soils experiencing normal field moisture level10 (Fig. 6a). Thus, although the clockwise effect of physical processes had no influence on the rate of RA and the rotational direction of hysteresis loop, it could have modified the orientation of the principal axes of the hysteresis loop of RA (Fig. 6a). Consequently, the co-occurrence of biological mechanism (diel photosynthetic carbon supply) and physical processes, particularly thermal transport, could have contributed to the counter-clockwise diel hysteresis of RA in this desert ecosystem.

Conceptual illustration of factors controlling diel hysteresis patterns of soil CO2 efflux (RS) and its components (RA and RH) in responding to soil temperature in dry (a) and wet (b) soils.
The effect of physical (mainly heat) transport processes on the diel hysteresis of RA and RH is same. Besides physical process, the diel hysteresis of RH is also modified by chemical (inorganic CO2 efflux, RINO) process. In dry soil condition (a), the effects of physical and chemical processes to the diel relationship between soil CO2 efflux and soil temperature are pronounced, while the effects are limited under wet soil condition (b). See the text in the Discussion for more details.
On the other hand, the clockwise hysteresis effect of RH could not be associated with carbon supply mechanism because there were no obvious root biomass in the bare soils. The explanation for the diel hysteresis of RH is still unclear, but three possible mechanisms may account for it. First, physical processes associated with heat transport and CO2 diffusion mentioned above may be responsible for the diel hysteresis of RH (Fig. 6a). Second, many previous studies have reported that in arid ecosystems with saline/alkaline soils, there is a chemical (i.e. carbonate precipitation and dissolution) process (inorganic CO2 efflux, RINO) influencing the diel variation of soil CO2 efflux because RINO shows a positive value during daytime (CO2 emission) and a negative value (CO2 absorption) at nighttime40,41,42,43 (Fig. 6a). Based on the diel relationship between RINO process and soil temperate in Fig. 6a, which is redrawn based on results of Liu et al.43 and Ma et al.40 in saline/alkaline soil desert ecosystems located near our study site, we suggest that the diel variation in RINO could have affected the observed diel hysteresis of RH by modifying the rate of RH and the roundness of hysteresis loop. However, the diel variation RINO would have no influence on the orientation of the principal axes of the hysteresis loop of RH and the rotational direction of hysteresis loop. Third, in desert ecosystems, microbial communities such as bacteria and fungi usually have different vertical distribution to compete for available substrates44,45,46. The differential temperature sensitivities of microbial components47,48,49 may have contributed to the diel hysteresis of RH. In addition, the growth rates of bacteria and fungi usually have optimum temperatures around 25–30 °C49. The higher temperature (above 33 °C) at noon at this site could have inhibited the activity of microbes, leading to a clockwise hysteresis loops. We propose that the above three mechanisms together resulted in the clockwise diel hysteresis of RH (Fig. 6a). However, we found that RH had a strong temporal coincidence with photosynthetic active radiation (PAR) (Fig. S2), which reached a maximum at 12:00. This is earlier than the soil temperature maximum, which occurred around 15:00, indicating that diel pattern of RH in this desert ecosystem may be more closely related to near surface soil temperature. However, it does not necessarily imply that all microbial respiration occurs in surface soils, as microbial respiration at deep soils certainly occur.
In addition, a clockwise hysteresis loop of RS was observed in responding to diel change of soil temperature, which is similar to that of RH but distinct from that of RA. The clockwise diel pattern could be attributed to the high contribution of RH to RS, particularly when soil temperature was increasing. Many previous studies suggest that the diel hysteresis between soil CO2 efflux and soil temperature is an artifact due to an arbitrary selection of a reference soil depth, which can produce an unrealistic relationship between efflux and soil temperature19,50. By exploring the relationship between soil CO2 efflux and its two components with air temperature (Fig. S3), we found a distinct, elliptical diel hysteresis, suggesting a potential occurrence of diel hysteresis of soil CO2 efflux in responding to soil temperature at different depths and also highlighting the complexity of understanding the processes controlling the relationship between soil CO2 efflux and temperature. Thus, we argue that, in order to accurately understand the temperature dependence of RS, it is necessary to include measurements of soil temperature at several depths.
Rain addition resulted in a larger magnitude of diel hysteresis of RA on Day 6 when soil moisture was higher. Many studies have shown that an effective rainfall can infiltrate into deep soils and be absorbed by plants with deep-rooted systems, leading to an increase in photosynthesis, especially in desert and semiarid ecosystems24,51. In this study, photosynthesis of N. tangutorum significantly increased after rain addition treatments (unpublished data). This higher photosynthesis of plants implied that a larger portion of photosynthetic C was transported into belowground and metabolized by roots33,37, which was of sufficient magnitude to alter diel patterns of RA32,52. However, increased soil water moisture following rain addition could have altered the effects of physical processes by increasing the rate of heat diffusivity and decreasing the rate of CO2 transport. The change in the two physical transport processes have consistent effect on diel hysteresis that cause clockwise diel hysteresis to become less pronounced10. Therefore, the limited effect of physical mechanism could have contributed to the change of diel hysteresis of RA because physical processes had contrasting effect on the orientation of hysteresis loop. We found that rain addition had no effect on the diel hysteresis of RA on Day 16 when soil was drier. The different effect of rain addition for the two days was most likely a consequence of variation in soil water content. On day 16, the lower soil water availability could have weakened the activity of plants and reduced the coupling between plant photosynthesis and belowground processes by reducing the movement of recent photosynthates to phloem loading sites and by changing the flow rate of photosynthates from the shoots into the roots53,54. This would potentially stifle the effects of rain addition on diel hysteresis of RA on Day 16. In addition, the increasing effect of physical processes following soil drying could be responsible for the consistent values of diel hysteresis among treatments on Day 16.
Similarly, soil water availability is a vital factor in controlling microbial activity in desert and semiarid ecosystems55,56,57. After rewetting following simulated rain addition, microbial activity recovered quickly due to increased substrates availability and favorable environmental conditions23,51,58, leading to a large production of CO2 gas. In addition, the change in the two physical transport processes under rain addition treatments could have caused clockwise diel hysteresis to become less pronounced (Fig. 6b) because physical processes have a consistent effect on the orientation of hysteresis loop. Furthermore, increasing soil water content could decrease the effects of RINO on diel hysteresis because the wetter the saline/alkaline soils are, the less effect RINO will have on the diel pattern of RH40,41,42. Therefore, compared with dry soil (Fig. 6a) or control plots (CK), rain addition treatments could have diminished the effects of physical processes and RINO on the diel hysteresis of RH.
Why was there an increase in the diel hysteresis of RH on Day 6 after rain addition? We propose two possible mechanisms for this phenomenon. First, the high soil moisture enhanced the diffusion of organic molecules through soil water films and increased substrate availability for microbial mineralization23,59. This would have affected microbial activity differently in various soil layers because microbial communities have a vast vertical distribution and thus have different moisture sensitivities47,48,49, increasing the diel hysteresis of RH on Day 6. Second, rain addition eliminated environment stress and provided a favorable (cool, wet) microclimate for microbial activity at a diel scale, particularly at noon32. The higher RH rate and lower soil temperature at daytime resulted in an increase in the diel hysteresis of RH. Whereas on Day 16 when the soil was drier, the diel hysteresis of RH decreased and showed no significant difference among treatments. Previous studies have shown that an effective rainfall event could trigger a rapid pulse response of microbial activity and the response would gradually decrease due to exhaustion of accessible organic matter55,60,61. The declined substrate availability on Day 16 could have also inhibited the temperature response of microbial activity, particularly at midday when temperature reached maximum value, leading to a consistent diel hysteresis value of RH among treatments. In addition, the consistent value could be attributed to the increasing role of physical mechanism discussed above. Our findings are consistent with other research showing that soil water availability can modify the magnitude of diel hysteresis between soil CO2 efflux and soil temperature15. In addition, we found that the change in soil moisture would only change the diel magnitude of RH with no effect on orientation. These results derived from field experiment is consistent with theoretical analysis10.
Surprisingly, we found that rain addition had no influence on the diel hysteresis of RS on the two measurement days, although the daily mean rate of RS significantly increased by rain addition treatments on both days during the growing season32. In this study site, the increased size of clockwise hysteresis loop of RH would be offset by the increased size of counter-clockwise hysteresis loop of RA, which could have resulted in the consistent value for diel hysteresis of RS among the five rain addition treatments during the growing season.
In addition, we found that, without considering diel hysteresis, significant error would occur in the estimation of daily soil CO2 efflux based on a relationship between efflux and temperature. Thus, diel hysteresis should be considered in terrestrial ecosystem models for predicting regional C cycles in desert regions. The method of using nighttime soil CO2 efflux data to predict daytime soil CO2 efflux, a common method in eddy covariance studies, resulted in an underestimation of daily RS and RH by 10–32% and 41–62% respectively, but an overestimation of daily RA by 5–12% in this desert ecosystem. However, in a Chihuahuan desert shrubland, Hamerlynck et al.62 showed an overestimation of daily soil CO2 efflux by 9–19% based on temperature-derived functions that do not consider diel hysteresis. Similarly, Riveros-Iregui et al.15 found an overestimation of growing season total soil CO2 efflux by 19% in a montane conifer forest when diel hysteresis was not considered. These results indicate that in different ecosystems with varying relative contribution of the two respiration components to the total efflux, the value of the error would be different. Thus, in order to accurately estimate ecosystem C budgets under future climate scenarios, we should consider the different responses of RA and RH to climate change, particularly to rainfall variability.
Implications and limitations
The results from our field experimental studies have important implications for regional carbon cycle simulations. First of all, one can no longer confidently capture an entirety of temperature response in soil CO2 efflux by only measuring soil CO2 efflux late morning because of the present of hysteresis. At the ecosystem-scale, eddy covariance-based studies often use nighttime respiration data to predict daytime rates of respiratory efflux in accordance with exponential temperature extrapolation63. The results of this study underscore that the failure to account for possible hysteresis of soil CO2 efflux at diel and/or seasonal scales may result in under- or overestimation of ecosystem respiration36,64, which is composed of both soil and aboveground component efflux. In addition, the occurrence of diel hysteresis suggests a potential problem in the usage of constant temperature sensitivity of soil CO2 efflux for gap filling procedures and in ecosystem carbon cycling models65. Lastly, higher soil water content results in a larger amplitude of diel hysteresis, implying that the role of soil water content in controlling the relationship between soil CO2 efflux and soil temperature should be considered in the modeling of production and efflux of CO2 from the soils in desert and semiarid ecosystems subjected to severe seasonal changes in soil water availability.
Although we investigated the effects of experimental rain addition on diel hysteresis of soil CO2 efflux and its components (RH and RA) in a desert ecosystem, this study was limited by several uncertainties. First, the calculation of diel hysteresis was based on the relationship between soil CO2 efflux and temperature at a soil depth of 10 cm. It is worth noting that the soil temperature used in our analysis is crucial because the phase and peak of soil temperature in different soil layers vary due to altered heat transport rate19,50, despite finding a distinct diel hysteresis between soil CO2 efflux and air temperature. Therefore, to understand exactly the diel hysteresis, several soil depth samples should be taken into account in further experiments. Second, the method of calculating RA in our study was based on the difference in the rate of soil CO2 efflux between the vegetated soil and the bare soil; however, this method may ignore the presence of rhizosphere respiration. Thus our method may over- or under-estimate the diel response of RA to soil temperature and result in a calculated deviation in diel hysteresis of RA. However, it should be noted that among the different partitioning methods reviewed by Hanson et al.66 and Kuzyakov67, our approach seems to be the most appropriate method for separating RA and RH in this desert ecosystem. Third, several previous studies suggested that a rainfall pulse after a period of drought can result in a quick response in microbial activity within several hours and the response is often short-lived in desert and semiarid regions24,33. In this study, we measured diel variations in soil CO2 efflux on Days 6 and 16 after the rain addition, which may not capture such quick response of microbial activity to the rain addition. Therefore, to fully understand the response of diel hysteresis to rainfall pulse, continuous soil CO2 efflux measurements following rainfall events should be performed in further experiments. Finally, this study was conducted in a desert ecosystem and whether other ecosystems, such as grassland or forest ecosystems have a similar response in diel hysteresis of soil CO2 efflux and its components to rain addition is still unclear. Therefore, more experimental studies with better approaches (e.g. stable isotope labeling) are needed to better understand the diel hysteresis of soil CO2 efflux and its components as well as to accurately model soil C loss from terrestrial ecosystems under future climate scenarios.
Additional Information
How to cite this article: Song, W. et al. Contrasting diel hysteresis between soil autotrophic and heterotrophic respiration in a desert ecosystem under different rainfall scenarios. Sci. Rep. 5, 16779; doi: 10.1038/srep16779 (2015).
References
Valentini, R. et al. Respiration as the main determinant of carbon balance in European forests. Nature 404, 861–865 (2000).
Davidson, E. A., Janssens, I. A. & Luo, Y. Q. On the variability of respiration in terrestrial ecosystems: moving beyond Q10 . Global Change Biol. 12, 154–164 (2006).
Luo, Y. Q., Wan, S. Q., Hui, D. F. & Wallace, L. L. Acclimatization of soil respiration to warming in a tall grass prairie. Nature 413, 622–625 (2001).
Davidson, E. A., Belk, E. & Boone, R. D. Soil water content and temperature as independent or confounded factors controlling soil respiration in a temperate mixed hardwood forest. Global Change Biol. 4, 217–227 (1998).
Curiel Yuste, J., Janssens, I. A., Carrara, A. & Ceulemans, R. Annual Q10 of soil respiration reflects plant phenological patterns as well as temperature sensitivity. Global Change Biol. 10, 161–169 (2004).
Lloyd, J. & Taylor, J. On the temperature dependence of soil respiration. Funct. Ecol. 8, 315–323 (1994).
Carbone, M. S., Winston, G. C. & Trumbore, S. E. Soil respiration in perennial grass and shrub ecosystems: Linking environmental controls with plant and microbial sources on seasonal and diel timescales. J. Geophys. Res. 113, G09022; 10.1029/2007JG000611 (2008).
Song, W. et al. Vegetation cover and rain timing co-regulate the responses of soil CO2 efflux to rain increase in an arid desert ecosystem. Soil Biol. Biochem. 49, 114–123 (2012).
Raich, J. W. & Tufekcioglu, A. Vegetation and soil respiration: correlations and controls. Biogeochemistry 48, 71–90 (2000).
Phillips, C. L., Nickerson, N., Risk, D. & Bond, B. J. Interpreting diel hysteresis between soil respiration and temperature. Global Change Biol. 17, 515–527 (2011).
Niu, S. et al. Seasonal hysteresis of net ecosystem exchange in response to temperature change: patterns and causes. Global Change Biol. 17, 3102–3114 (2011).
Gaumont-Guay, D. et al. Interpreting the dependence of soil respiration on soil temperature and water content in a boreal aspen stand. Agr. Forest Meteorol. 140, 220–235 (2006).
Vargas, R. & Allen, M. F. Diel patterns of soil respiration in a tropical forest after Hurricane Wilma. J. Geophys. Res. 113, G03021; 10.1029/2007JG000620 (2008).
Vargas, R. & Allen, M. F. Environmental controls and the influence of vegetation type, fine roots and rhizomorphs on diel and seasonal variatin in soil respiration. New Phytol. 179, 460–471 (2008).
Riveros-Iregui, D. A. et al. Diurnal hysteresis between soil CO2 and soil temperature is controlled by soil water content. Geophys. Res. Lett. 34, L17404; 10.1029/2007GL030938 (2007)
Tang, J. W., Baldocchi, D. D. & Xu, L. Tree photosynthesis modulates soil respiration on a diurnal time scale. Global Change Biol. 11, 1298–1304 (2005).
Barron-Gafford, G. A., Scott, R. L., Jenerette, G. D. & Huxman, T. E. The relative controls of temperature, soil moisture and plant functional group on soil CO2 efflux at diel, seasonal and annual scales. J. Geophys. Res. 116, G01023; 10.1029/2010JG001442 (2011).
Li, Z., Wang, X., Zhang, R., Zhang, J. & Tian, C. Contrasting diurnal variations in soil organic carbon decomposition and root respiration due to a hysteresis effect with soil temperature in a Gossypium s. (cotton) plantation. Plant Soil 343, 347–355 (2011).
Parkin, T. B. & Kaspar, T. C. Temperature controls on diurnal carbon dioxide flux: Implications for estimating soil carbon loss. Soil Sci. Soc. Am. J. 67, 1763–1772 (2003).
Oikawa, P. et al. Unifying soil respiration pulses, inhibition and temperature hysteresis through dynamics of labile soil carbon and O2 . J. Geophys. Res. 119, 521–536; 10.1002/2013JG002434 (2014).
Kuzyakov, Y. & Gavrichkova, O. Time lag between photosynthesis and carbon dioxide efflux from soil: a review of mechanisms and controls. Global Change Biol. 16, 3386–3406 (2010).
Sampson, D. A., Janssens, I. A., Yuste, J. C. & Ceulemans, R. Basal rates of soil respiration are correlated with photosynthesis in a mixed temperate forest. Global Change Biol. 13, 2008–2017 (2007).
Sponseller, R. A. Precipitation pulses and soil CO2 flux in a Sonoran Desert ecosystem. Global Change Biol. 13, 426–436 (2007).
Huxman, T. E. et al. Precipitation pulses and carbon fluxes in semiarid and arid ecosystems. Oecologia 141, 254–268 (2004).
Chen, H. Projected change in extreme rainfall events in China by the end of the 21st century using CMIP5 models. Chin. Sci. Bull. 58, 1462–1472 (2013).
Gao, X., Shi, Y., Zhang, D. & Giorgi, F. Climate change in China in the 21st century as simulated by a high resolution regional climate model. Chin. Sci. Bull. 57, 1188–1195 (2012).
IPCC. Summary for policymakers. In Climate Change 2013: The Physical Science Basis. Contribution of Working Group I to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change (eds Stocker, T. F. et al. ) (Cambridge Univerisity Press, Cambridge, United Kingdom and New York, NY, USA., 2013).
Noy-Meir, I. Desert ecosystems: enviroment and producters. Annu. Rev. Ecol. Syst. 4, 25–51 (1973).
Welsch, D. L. & Hornberger, G. M. Spatial and temporal simulation of soil CO2 concentrations in a small forested catchment in Virginia. Biogeochemistry 71, 415–436 (2004).
Ryan, M. G. & law, B. E. Interpreting, measuring and modeling soil respiration. Biogeochemistry 73, 3–27 (2005).
Li, J. W. et al. Modeling permafrost thaw and ecosystem carbon cycle under annual and seasonal warming at an Arctic tundra site in Alaska. J. Geophys. Res. 119, 1129–1146; 10.1002/2013JG002569 (2014).
Song, W. et al. Simulated rain addition modifies diurnal patterns and temperature sensitivities of autotrophic and heterotrophic soil respiration in an arid desert ecosystem. Soil Biol. Biochem. 82, 143–152 (2015).
Chen, S. P., Lin, G. H., Huang, J. H. & Jenerette, G. D. Dependence of carbon sequestration on the differential responses of ecosystem photosynthesis and respiration to rain pulses in a semiarid steppe. Global Change Biol. 15, 2450–2461 (2009).
Illeris, L., Michelsen, A. & Jonasson, S. Soil plus root respiration and microbial biomass following water, nitrogen and phosphorus application at a high arctic semi desert. Biogeochemistry 65, 15–29 (2003).
Huang, B. & Nobel, P. S. Hydraulic conductivity and anatomy along lateral roots of cacti: changes with soil-water status. New Phytol. 123, 499–507 (1993).
Phillips, S. C. et al. Interannual, seasonal and diel variation in soil respiration relative to ecosystem respiration at a wetland to upland slope at Harvard Forest. J. Geophys. Res. 115, 10.1029/2008JG000858 (2010).
Yan, L. M., Chen, S. P., Huang, J. H. & Lin, G. H. Water regulated effects of photosynthetic substrate supply on soil respiration in a semiarid steppe. Global Change Biol. 17, 1990–2001 (2011).
Bahn, M., Schmitt, M., Siegwolf, R., Richter, A. & Brueggemann, N. Does photosynthesis affect grassland soil-respired CO2 and its carbon isotope composition on a diurnal timescale? New Phytol. 182, 451–460 (2009).
Carbone, M. S. & Trumbore, S. E. Contribution of new photosynthetic assimilates to respiration by perennial grasses and shrubs: residence times and allocation patterns. New Phytol. 176, 124–135 (2007).
Ma, J., Wang, Z.-Y., Stevenson, B. A., Zheng, X.-J. & Li, Y. An inorganic CO2 diffusion and dissolution process explains negative CO2 fluxes in saline/alkaline soils. Sci. Rep. 3, 2025; 10.1038/srep02025 (2013).
Shanhun, F. L., Almond, P. C., Clough, T. J. & Smith, C. M. S. Abiotic processes dominate CO2 fluxes in Antarctic soils. Soil Biol. Biochem. 53, 99–111 (2012).
Ball, B. A., Virginia, R. A., Barrett, J. E., Parsons, A. N. & Wall, D. H. Interactions between physical and biotic factors influence CO2 flux in Antarctic dry valley soils. Soil Biol. Biochem. 41, 1510–1517 (2009).
Liu, J. et al. Abiotic CO2 exchange between soil and atmosphere and its response to temperature. Environ. Earth Sci. 73, 2463–2471 (2015).
Yu, J. & Steinberger, Y. Vertical distribution of soil microbial biomass and its association with shrubs from the Negev Desert. J. Arid Environ. 78, 110–118 (2012).
Rodriguez-Zaraqoza, S. et al. Vertical distribution of microbial communities under the canopy of two legume bushes in the Tehuacan Desert, Mexico. Eur. J. Soil Biol. 44, 373–380 (2008).
Shamir, I. & Steinberger, Y. Vertical distribution and activity of soil microbial population in a sandy desert ecosystem. Microb. Ecol. 53, 340–347 (2007).
Barcenas-Moreno, G., Rousk, J. & Baath, E. Fungal and bacterial recolonisation of acid and alkaline forest soils following artificial heat treatments. Soil Biol. Biochem. 43, 1023–1033 (2011).
Yuste, J. C. et al. Drought-resistant fungi control soil organic matter decomposition and its response to temperature. Global Change Biol. 17, 1475–1486 (2011).
Pietikäinen, J., Pettersson, M. & Bååth, E. Comparison of temperature effects on soil respiration and bacterial and fungal growth rates. FEMS Microb. Ecol. 52, 49–58 (2005).
Subke, J. A. & Bahn, M. On the ‘temperature sensitivity’ of soil respiration: Can we use the immeasurable to predict the unknown? Soil Biol. Biochem. 42, 1653–1656 (2010).
Austin, A. T. et al. Water pulses and biogeochemical cycles in arid and semiarid ecosystems. Oecologia 141, 221–235 (2004).
Savage, K., Davidson, E. A. & Tang, J. Diel patterns of autotrophic and heterotrophic respiration among phenological stages. Global Change Biol. 19, 1151–1159 (2013).
Li, X. M., Feng, Y. S. & Boersma, L. Partition of photosynthates between shoot and root in spring wheat (Triticum aestivum L.) as a function of soil water potential and root temperature. Plant Soil 164, 43–50 (1994).
Ruehr, N. K. et al. Drought effects on allocation of recent carbon: from beech leaves to soil CO2 efflux. New Phytol. 184, 950–961 (2009).
Fierer, N. & Schimel, J. P. A proposed mechanism for the pulse in carbon dioxide production commonly observed following the rapid rewetting of a dry soil. Soil Sci. Soc. Am. J. 67, 798–805 (2003).
Cable, J. M. & Huxman, T. E. Precipitation pulse size effects on Sonoran Desert soil microbial crusts. Oecologia 141, 317–324 (2004).
Liu, W., Zhang, Z. & Wan, S. Predominant role of water in regulating soil and microbial respiration and their responses to climate change in a semiarid grassland. Global Change Biol. 15, 184–195 (2009).
Boot, C. M. Microbial response to drying and rewetting: osmotic and matric effects. Plant Soil 348, 99–102 (2011).
Janssens, I. A. & Pilegaard, K. Large seasonal changes in Q10 of soil respiration in a beech forest. Global Change Biol. 9, 911–918 (2003).
Borken, W. & Matzner, E. Reappraisal of drying and wetting effects on C and N mineralization and fluxes in soils. Global Change Biol. 15, 808–824 (2009).
Fierer, N. & Schimel, J. P. Effects of drying-rewetting frequency on soil carbon and nitrogen transformations. Soil Biol. Biochem. 34, 777–787 (2002).
Hamerlynck, E. P., Scott, R. L., Sanchez-Canete, E. P. & Barron-Gafford, G. A. Nocturnal soil CO2 uptake and its relationship to subsurface soil and ecosystem carbon fluxes in a Chihuahuan Desert shrubland. J. Geophys. Res. 118, 1593–1603; 10.1002/2013JG002495 (2013).
Zhang, W. L. et al. Biophysical regulations of carbon fluxes of a steppe and a cultivated cropland in semiarid Inner Mongolia. Agr. Forest Meteorol. 146, 216–229 (2007).
Davidson, E. A., Richardson, A. D., Savage, K. E. & Hollinger, D. Y. A distinct seasonal pattern of the ratio of soil respiration to total ecosystem respiration in a spruce-dominated forest. Global Change Biol. 12, 230–239 (2006).
Richardson, A. D. et al. Comparing simple respiration models for eddy flux and dynamic chamber data. Agr. Forest Meteorol. 141, 219–234 (2006).
Hanson, P. J., Edwards, N. T., Garten, C. T. & Andrews, J. A. Separating root and soil microbial contributions to soil respiration: A review of methods and observations. Biogeochemistry 48, 115–146 (2000).
Kuzyakov, Y. Separating microbial respiration of exudates from root respiration in non-sterile soils: a comparison of four methods. Soil Biol. Biochem. 34, 1621–1631 (2002).
Acknowledgements
This study was supported in part by grants from the Science and Technologies Foundation of the Chinese Academy of Forestry (CAFYBB2007008), the “973” program of Ministry of Science and Technology, China (2013CB956601), the “Strategic Priority Research Program” of the Chinese Academy of Sciences, Climate Change: Carbon Budget and Relevant Issues (XDA05050402), the National Natural Science Foundation of China (31170453) and a Selected Young Scientist Program of the State Key Laboratory of Vegetation and Environment Change. We also thank Ms. Tianyi Zhang for her help with valuable comments on the manuscript.
Author information
Authors and Affiliations
Contributions
S.P.C., B.W., Q.L. and G.H.L. conceived and designed the experiments. W.M.S., S.P.C., Y.D.Z. and Y.J.Z. performed the experiments. W.M.S., S.P.C. and G.H.L. analysed the data and wrote the manuscript.
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Electronic supplementary material
Rights and permissions
This work is licensed under a Creative Commons Attribution 4.0 International License. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in the credit line; if the material is not included under the Creative Commons license, users will need to obtain permission from the license holder to reproduce the material. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/
About this article

Cite this article
Song, W., Chen, S., Zhou, Y. et al. Contrasting diel hysteresis between soil autotrophic and heterotrophic respiration in a desert ecosystem under different rainfall scenarios. Sci Rep 5, 16779 (2015). https://doi.org/10.1038/srep16779
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/srep16779
This article is cited by
-
Temperature controls diel oscillation of the CO2 concentration in a desert soil
Biogeochemistry (2021)
-
Biotic soil-plant interaction processes explain most of hysteretic soil CO2 efflux response to temperature in cross-factorial mesocosm experiment
Scientific Reports (2020)
-
Soil respiration in an irrigated oasis agroecosystem: linking environmental controls with plant activities on hourly, daily and monthly timescales
Plant and Soil (2020)
-
Livestock induces strong spatial heterogeneity of soil CO2, N2O and CH4 emissions within a semi-arid sylvo-pastoral landscape in West Africa
Journal of Arid Land (2017)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
