
Abstract
By cultivating turf algae and aggressively defending their territories, territorial damselfishes in the genus Stegastes play a major role in shaping coral-algal dynamics on coral reefs. The epilithic algal matrix (EAM) inside Stegastes’ territories is known to harbor high abundances of potential coral disease pathogens. To determine the impact of territorial grazers on coral microbial assemblages, we established a coral transplant inside and outside of Stegastes’ territories. Over the course of one year, the percent mortality of transplanted corals was monitored and coral samples were collected for microbial analysis. As compared to outside damselfish territories, Stegastes were associated with a higher rate of mortality of transplanted corals. However, 16S rDNA sequencing revealed that territorial grazers do not differentially impact the microbial assemblage of corals exposed to the EAM. Regardless of Stegastes presence or absence, coral transplantation resulted in a shift in the coral-associated microbial community and an increase in coral disease associated potential pathogens. Further, transplanted corals that suffer low to high mortality undergo a microbial transition from a microbiome similar to that of healthy corals to that resembling the EAM. These findings demonstrate that coral transplantation significantly impacts coral microbial communities and transplantation may increase susceptibility to coral disease.
Similar content being viewed by others

Introduction
Microbial diversity is essential for the functioning and resilience of terrestrial and marine ecosystems1. On coral reefs, microbial communities influence biogeochemical and ecological processes such as nutrient cycling and larval recruitment2,3. Reef-building corals are host to well-studied obligate symbionts in the genus Symbiodinium, but less is known about the diverse assemblages of bacteria that associate with corals4. Microbial communities are hypothesized to confer many benefits to their coral hosts, such as energy provision, photosynthesis, nitrogen fixation and the prevention of infection. However, under disturbances and stressful environmental conditions, shifts in the coral microbiome have been linked to the degradation of coral reefs5. Yet, current research does not adequately address the potential impacts of trophic interactions on microbial communities on coral reefs.
In the marine environment, benthic microbial communities are influenced by an array of abiotic and biotic factors, including nutrient fluxes and benthic-feeding organisms6,7. Territorial damselfishes are abundant and important engineers of the reef benthos and play a key role in benthic dynamics8,9,10. These fishes exhibit several key behaviors that extensively modify benthic structure in their territories: grazing turf algae, weeding unpalatable macroalgae, coral-pecking to further propagate turf algae and engaging in constant aggression to protect their algal resources from intruders11,12,13. Although turf algae is omnipresent across the reef benthos, territorial damselfishes cultivate a notably thicker turf inside their territories and damselfishes in the genus Stegastes maintain low-diversity algal turfs on the benthos and coral skeletons9,14. Thus, damselfish territories are dominated by the epilithic algal matrix (EAM), a conglomeration of turf algae, juvenile macroalgae, detritus, invertebrates and bacterial assemblages15,16,17,18.
The EAM is known to have negative effects on coral growth and survival both inside and outside of damselfish territories19,20,21,22,23,24; therefore, due to their cultivation of a thick EAM, the impact of territorial grazers on corals is largely negative. Territorial grazers have been shown to inhibit coral recruitment25, negatively impact the abundance of juvenile corals26 and cause mortality in adult corals11. The EAM inside Stegastes’ territories has also been shown to harbour potential pathogens linked to black band disease (BBD)7. Due to the elevated levels of potential coral disease pathogens in the EAM, territorial grazers may indirectly promote disease and mortality in corals inside their territories. Direct contact-mediated coral-algal interactions may cause toxicity or hypoxia in coral tissues, thus facilitating the invasion of opportunistic pathogens. This stress-induced increase in potential pathogens may ultimately lead to coral disease or death18.
Although corals and algae have a highly antagonistic relationship on coral reefs, they are known to harbour distinct microbial communities20 and corals can outcompete turf algae in areas of low anthropogenic disturbance27. While territorial damselfishes in the genus Stegastes have been shown to negatively impact corals, the relationship between damselfishes and corals is variable depending on region and habitat and damselfishes have also been reported to increase coral recruitment28 and facilitate adult coral survival29,30,31,32,33. Therefore, the high occurrence of coral disease-associated bacteria in the EAM inside Stegastes’ territories is not necessarily indicative of the composition of neighboring coral microbial assemblages. Thus, this study aims to determine the coral survivorship and the microbial composition of corals within territories of damselfishes in the genus Stegastes. Specifically, we determined the impact of Stegastes on the survival of transplanted corals and analyzed how coral transplantation inside and outside of Stegastes’ territories affects coral microbial communities.
Methods
Study site and species
This study took place at Lizard Island in the northern GBR, Australia (14°41′5”S, 145°26′55”E) from February 2012 to August 2013. The study site was on the back reef in the lagoon between Palfrey and South Island at a depth of 1–3 m.
Stegastes nigricans and Stegastes apicalis (f. Pomacentridae), two intensive territorial grazers, were our damselfish study species. S. apicalis occurs at a depth of 1–15 m and reaches up to 15 cm (total length). S. nigricans occurs at the depth of 1–12 m and reaches up to 14 cm (total length). Both species form social groups (termed “colonies”) made up of several contiguous territories, where each territory belongs to an individual adult damselfish.
Acropora muricata was our coral study species. It is a common staghorn coral with aborescent branching in shallow reefs around Lizard Island, especially lagoon and back reef habitats. Stegastes typically form colonies within or around outcrops of A. muricata.
Coral baseline samples
To determine the impact of territorial damselfishes on coral microbial communities, we first collected coral branches from outside and inside damselfish territories. In the field, ten 15 cm coral branches were collected from control plots outside damselfish territories, ten from inside S. apicalis’ territories and ten from inside S. nigricans’ territories. For the collections from S. apicalis’ and S. nigricans’ territories, branches were taken across two damselfish colonies and each branch came from a different damselfish territory.
Coral transplant
To determine the effects of EAM exposure and damselfish farming behaviours on coral fragments over time, we set up a coral transplant outside and inside damselfish territories. Due to the higher success rate of coral transplantation of A. muricata when medium to large-sized fragments (10–20 cm) are used34, we sourced 120 15-cm fragments of A. muricata from outside damselfish territories. To minimize handling time, coral fragments were briefly transported to large seawater bins for labeling at the field site, then immediately transplanted using marine epoxy. Coral fragments were directly transplanted rather than allocating a recovery period in holding tanks because placing corals in holding tanks after fragmentation can substantially alter bacterial assemblages35 and experimental injuries (such as wounding from fragmentation) have been shown to have a limited impact on the coral immune response36. Forty fragments were transplanted in control plots outside of damselfish territories, forty fragments were transplanted inside S. apicalis’ territories and forty fragments were transplanted in S. nigricans’ territories. Coral fragments were distributed randomly across treatments with respect to the source colonies. Two transplanted corals were placed in each Stegastes’ territory and the transplanted corals were distributed across two S. apicalis’ colonies and two S. nigricans’ colonies, which were the same colonies that were used for baseline sample collections.
After six months, the percent mortality (percentage of tissue loss) of each coral fragment was estimated and ten coral fragments were sampled from each treatment for microbial analyses. Percent mortality estimates and microbial sampling (ten coral fragments from each treatment) were repeated after one year. Thus, we sampled a total of 90 coral fragments: 30 baseline samples, 30 samples after six months of transplantation and 30 samples after one year of transplantation. Of each 30 samples, ten were from control plots outside of damselfish territories, ten were from S. apicalis’ territories and ten were from S. nigricans’ territories.
Microbial processing
After collection, coral fragments were immediately snap-frozen in liquid nitrogen, stored at −80°C and transported to James Cook University for processing and DNA extractions. Samples were homogenized under liquid nitrogen. When tissue fragments suffered partial to high mortality, care was taken to homogenize only live tissue sections of the coral fragments (dead tissue and algae coverage was excluded). See Casey et al.7 for methods of DNA isolation, quality control, PCR and 16S rDNA sequencing.
Coral mortality data analysis
To analyze the effect of intensive territorial grazers on the mortality of transplanted corals, we used Akaike’s Information Criterion (AIC) to compare the fit of three generalized linear models (GLM) to our data (Table 1). Due to preferential removal of coral fragments with low mortality for microbial analysis after six months of transplantation, we focused on mortality only over the first six month period to avoid biases associated with sample removal. For each model, the response was multinomial (low mortality, partial mortality and high mortality in transplanted coral): low mortality was defined as 0–20% loss of tissue from the coral branch, partial mortality was defined as 20–80% loss of tissue from the coral branch and high mortality was defined as 80–99% loss of tissue from the coral branch (Fig. 1). In the first GLM, the fixed effect was the presence or absence of a damselfish territory, separating the effects of damselfish species (control plots with no damselfish territory versus S. apicalis’ territories and S. nigricans’ territories). In the second GLM, the fixed effect was also the presence or absence of a damselfish territory, without separating the effects of damselfish species (control plots with no damselfish territory versus damselfish territories). The third GLM was an intercept-only model with no treatment effects. The analysis was conducted with the package nnet37 using the software program R.

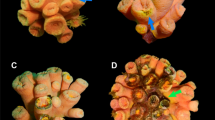
Mortality of transplanted corals in Stegastes’ territories after six months.
(a) Low mortality (0–20%), (b) partial mortality (20–80%) and (c) high mortality (80–99%). Photographs taken by J.M.C.
Microbial data analysis
The sequence data were processed using a proprietary analysis pipeline at MR DNA and re-analyzed with Quantitative Insights Into Microbial Ecology (QIIME)38. For both the proprietary analysis pipeline at MR DNA and QIIME, sequences were depleted of barcodes and primers and short sequences (<200 bp) and sequences with homopolymer runs exceeding 6 bp were removed. The average read length was 437 bp after primer and barcode removal. See Casey et al.7 for further methods of denoising, normalization, definition of operational taxonomic units (OTUs) and taxonomical assignments with QIIME.
We assessed the beta-diversity of the coral and EAM microbial communities (EAM samples were collected concurrently at the same study site; data published by Casey et al.7) inside and outside of damselfish territories with QIIME using a weighted UniFrac analysis. A principal coordinates analysis (PCoA) was generated from the UniFrac distances39. A PCoA was generated from weighted UniFrac distances and plotted in two dimensions.
Individual OTUs were assigned into three categories: autotrophs, heterotrophs and potential pathogens, based on literature reviews (Supplementary Table S1). All genera with a less than two percent relative abundance were excluded from the analyses (Supplementary Tables S2-S10). Our data included twenty-one genera that are considered potential pathogens for a broad array of hosts; however, among these genera, four are specifically linked to coral disease. These four coral-specific potential pathogens made up 26.4% percent relative abundance of the potential pathogen category and we focused on these coral disease associated potential pathogens in a further analysis.
We fit GLMs to analyze the differences in the relative abundances of autotrophs, heterotrophs, potential pathogens and potential coral disease pathogens in coral microbial communities outside of damselfish territories as compared to inside S. apicalis’ territories and S. nigricans’ territories and across the three sampling periods (baseline, six month of transplantation and one year of transplantation). We used a quasi-binomial error structure to account for the fact that the response variable (relative abundance) varied continuously between zero and one. The fixed effects included treatment (control plots outside of damselfish territories, inside S. apicalis’ territories and inside S. nigricans’ territories) and time (baseline, transplantation after six months and transplantation after one year) and their interactions. Thus, these models simultaneously analyzed the effects of damselfish presence as well as time of transplantation on the metabolic groupings of coral bacterial assemblages. Since we employed a quasi-binomial error structure in this analysis, likelihood-based model selection, such as AIC, could not be used. Instead, we employed a quasi-likelihood procedure based on adjusted model deviances, which utilizes the standard F distribution as the null distribution (Table 2, see Zuur et al.40). The analysis was conducted with the packages lme441 and arm using the software program R.
Results
Mortality of transplanted corals
Model selection indicated that the best model of coral mortality included the effects of both damselfish species by comparing control plots with no damselfish territory to S. apicalis’ territories and S. nigricans’ territories. Specifically, after six months, corals transplanted inside damselfish territories suffered a higher mortality (loss of tissue) than corals outside damselfish territories, with coral fragments inside S. nigricans’ territories exhibiting a stronger estimated response than fragments inside S. apicalis’ territories (Table 1, Fig. 1 and Fig. 2).

Percent of transplanted coral fragments (±SE) that suffered mortality after six months of transplantation.
Mortality is categorized as low mortality (0–20%), partial mortality (20–80%) and high mortality (80–99%). Treatments include control plots outside damselfish territories (35 fragments), inside S. apicalis’ territories (34 fragments) and inside S. nigricans’ territories (36 fragments). Bars represent means and standard errors of percentages of fragments in each mortality category across control plots outside damselfish territories and inside different damselfish territories (n = 20 territories in each case). Asterisks indicate significant values (p < 0.05).
Coral microbial communities
Bacterial 16S rDNA gene amplicon sequencing retrieved 235,388 high-quality sequence reads from 77 coral samples (DNA extractions and sequencing were successful for 77 coral samples out of the 90 collected coral fragments). Sequence reads were normalized to 960 reads per sample to allow for comparison between samples and bacterial community patterns, which further excluded two samples, resulting in 75 coral samples. In an analysis of all bacterial phylotypes, genera were assigned into the metabolic groupings of autotrophs, heterotrophs and potential pathogens (Supplementary Table S1), revealing that microbial communities in the coral fragments shift according to the presence of damselfish territories and over the course of one year of transplantation (Fig. 3). Model selection indicated that the best model for the relative abundance of autotrophs included the main effects of treatment (damselfish effects) and time, but no interaction (Table 2). As compared to baseline coral microbial communities, there were significant increases in the relative abundance of autotrophs after six months (p = 0.001) and one year (p = 0.008) of transplantation (Fig. 3). Further, as compared to corals transplanted outside of damselfish territories, corals inside S. nigricans’ territories had significantly lower relative abundances of autotrophs (p = 0008). For heterotrophs, model selection indicated that the best model included the effect of time only (Table 2). As compared to baseline coral microbial communities, there were significant decreases in the relative abundance of heterotrophs after six months (p < 0.001) and one year (p < 0.001) of transplantation (Fig. 3). Lastly, model selection indicated that the best model for potential pathogens included the full model: effects of treatment (damselfish effects), time and their interactions (Table 2). The relative abundance of potential pathogens was impacted by a significant interaction (p = 0.002). Visual inspection suggests that changes in potential pathogens across treatments and time were driven by larger increases over the first six months in control fragments relative to those in damselfish territories, particularly those of S. nigricans (Fig. 3).

Relative abundances of autotrophs, heterotrophs and potential pathogens in coral fragments.
The relative abundance (±SE) of (a) autotrophs, (b) heterotrophs and (c) potential pathogens according to damselfish presence (control plots outside damselfish territories, inside S. apicalis’ territories and inside S. nigricans’ territories) and time after transplantation (baseline coral fragments, transplanted coral fragments after six months and transplanted coral fragments after one year).
Potential coral disease pathogens
Model selection indicated that the best model for the relative abundance of potential coral disease pathogens included the main effects of treatment (damselfish effects) and time, but no interaction (Table 2). As compared to baseline coral fragments, acroporid fragments experienced significantly higher relative abundances in potential pathogens associated with coral disease after six months (p = 0.001) and to a lesser extent, after one year (p = 0.008) of transplantation (Fig. 4). The potential coral disease pathogens were in the genera Geitlerinema, Leptolyngbya, Oscillatoria, and Sphingomonas42,43. Of these coral-disease associated potential pathogens, Geitlerinema, Leptolyngbya and Oscillatoria are associated with BBD42. Also, as compared to no-damselfish controls, there were significantly lower relative abundances of potential pathogens associated with coral disease in coral fragments inside S. nigricans’ territories (p = 0.008), but the lower abundances inside S. apicalis’ territories did not differ significantly from controls (p = 0.073, Fig. 4).

Relative abundances of potential coral disease pathogens in coral fragments.
The relative abundance (±SE) of coral disease associated potential pathogens according to damselfish presence (control plots outside damselfish territories, inside S. apicalis’ territories and inside S. nigricans’ territories) and time after transplantation (baseline coral fragments, transplanted coral fragments after six months and transplanted coral fragments after one year).
Discussion
This study reveals that the presence of territorial damselfishes increases the rate of mortality of corals relocated to within territories. As compared to benthic plots outside damselfish territories, S. nigricans trigger the highest rate of mortality in transplanted corals, with S. apicalis causing an intermediate rate of mortality between plots outside of damselfish territories and S. nigricans’ territories. It has been shown that S. nigricans engage in more intensive farming behaviours than S. apicalis7; therefore, it is likely that S. nigricans’ intensive cultivation of the EAM or direct polyp mortality by coral-pecking may outcompete stressed, transplanted corals. Surprisingly, despite these higher rates of mortality, Stegastes do not have a differential impact on coral microbial communities. While previous work has shown that there are higher relative abundances of coral disease pathogens in the EAM inside Stegastes’ territories7, this study demonstrates that shifts in bacterial assemblages in corals after transplantation are not directly related to the impact of territorial damselfishes.
However, the damage caused by coral transplantation is found to impact the microbial community of corals. Six months after coral transplantation, over fifty percent of the coral fragments suffered partial to high mortality. As a result of this high rate of mortality, coral transplantation both inside and outside damselfish territories also triggered a shift in the coral microbiome that is evident up to a year after the initial transplantation. To further analyze how the microbiome of transplanted corals shift over time as a function of percent coral mortality, we examined the similarity of transplanted coral microbial communities to EAM microbial communities (EAM data published by Casey et al.7). This analysis reveals a transition in the microbial community of baseline coral samples and the healthy tissue of transplanted corals with low (0–20%) mortality to transplanted corals with partial (20–80%) mortality to transplanted corals with high (80–99%) mortality (Fig. 5). The microbial community of transplanted corals with high mortality is more similar to the EAM microbial community than to the baseline coral microbial community. Exposure to the benthos and consequently the EAM, has a transformative effect on microbial communities of transplanted corals that suffer partial to high mortality, as the coral tissue undergoes a microbial transition from an association with healthy corals to an association with the EAM. This microbial shift, in which coral microbial communities increasingly resemble the EAM microbial community, has major implications for benthic microbial assemblages. When benthic microbial communities become homogenous, from the EAM to corals, this loss of microbial diversity may lead to decreased resilience against potential coral disease pathogens44,45,46.

Principal coordinates analysis (PCoA) showing the percent of variation explained in the microbial community.
Treatments include baseline coral fragments, transplanted corals with low mortality (0–20%), transplanted corals with partial mortality (20–80%), transplanted corals with high mortality (80–99%) and EAM samples (data published in Casey et al.7) outside damselfish territories, inside S. apicalis’ territories and inside S. nigricans’ territories. The ellipses represent distinct clustering of the baseline corals, transplanted corals and EAM samples.
Previous work shows that there are higher relative abundances of BBD pathogens in the EAM inside Stegastes’ territories as well as a higher occurrence of BBD inside S. nigricans’ territories7. This study reveals that the higher relative abundance of potential coral disease pathogens in the EAM inside Stegastes’ territories may opportunistically cause BBD in acroporids, but it does not demonstrate nor predict shifts in the overall coral microbial assemblage in A. muricata inside Stegastes’ territories. The fact that BBD pathogens are omnipresent, albeit in lower abundances, across the reef benthos allows them to opportunistically colonize stressed transplanted corals, regardless of Stegastes presence or absence. It is known that even healthy corals have potential pathogens in their bacterial assemblages and under changing environmental conditions, a commensal may transition to a pathogenic state44. Here, we find that one prevalent taxon among our samples has been previously linked to disease in corals. Bacteria in the genus Ruegeria were consistently present within 50 percent of all coral fragments and previously have been associated with both healthy corals and coral lesions resulting from Yellow Band Disease46. Ruegeria also undergoes horizontal gene transfer, which may help hosts and microbial associates adapt to environmental challenges in short time periods47,48. Despite the suggestion of a possible link between Ruegeria and the promotion of coral disease46,49,50, the common occurrence of Ruegeria in baseline and transplanted corals suggests that the role of these bacteria is a commensal in the current study.
A considerable number of studies have investigated the efficacy of coral transplantation as a means for coral reef restoration by examining how transplantation affects coral growth, mortality and physiology51,52,53,54,55,56,57. While it is known that environmental stressors5,58,59,60,61 and coral disease49,50,62,63,64,65,66 cause changes in coral microbial communities, no previous study has analyzed the impact of coral transplantation on coral microbial communities. Due to the use of coral transplantation for coral reef restoration53,54,55,56,57,67,68,69,70,71,72, this paper demonstrates how transplantation may negatively impact the survival and health of corals. Since microbes are key players in coral health, it is imperative to consider microbial communities when examining the utility of conservation measures such as coral transplantation1.
Here, we show that coral fragments undergo higher rates of mortality inside damselfish territories, but territorial grazers do not differentially affect the microbial communities of transplanted corals. Rather, the damage caused by coral transplantation leads to a shift in the microbial community toward an increase in potential coral disease pathogens, especially those linked to BBD42, which is independent of territorial grazer presence or absence. The increase in potential pathogens in transplanted corals suggests that transplanted corals may be more susceptible to coral disease under certain stressful environmental conditions, such as an increase in sea surface temperature or nutrient fluxes. This study highlights the importance of examining ecological interactions beyond trends of macro-organisms and demonstrates how microbial communities provide essential information about coral health and resilience1,4,73.
Additional Information
Accession codes: The DNA sequence data appears in the National Center for Biotechnology Information (NCBI) under BioProject submission ID SUB924371. http://www.nature.com/srep
How to cite this article: Casey, J. M. et al. Coral transplantation triggers shift in microbiome and promotion of coral disease associated potential pathogens. Sci. Rep. 5, 11903; doi: 10.1038/srep11903 (2015).
References
Garren, M. & Azam, F. New directions in coral reef microbial ecology. Environ. Microbiol. 44, 833–833 (2012).
Ainsworth, T. D., Vega Thurber, R. & Gates, R. D. The future of coral reefs: a microbial perspective. Trends Ecol. Evol. 25, 233–240 (2009).
Webster, N. S. et al. Metamorphosis of a scleractinian coral in response to microbial biofilms. Appl. Environ. Microbiol. 70, 1213–1221 (2004).
Knowlton, N. & Rohwer, F. Multispecies microbial mutualisms on coral reefs: The host as a habitat. Am. Nat. 162, S51–S62 (2003).
Vega Thurber, R. et al. Metagenomic analysis of stressed coral holobionts. Environ. Microbiol. 11, 2148–2163 (2009).
Alongi, D. M. The role of bacteria in nutrient recycling in tropical mangrove and other coastal benthic ecosystems. Hydrobiologia. 285, 19–32 (1994).
Casey, J. M., Ainsworth, T. D., Choat, J. H. & Connolly, S. R. Farming behaviour of reef fishes increases the prevalence of coral disease associated microbes and black band disease. Proc. R. Soc. B. 281, 20141032 (2014).
Choat J. H. [The biology of herbivorous fishes on coral reefs] The ecology of fishes on coral reefs (ed. Sale, P. ) 120–155 (Academic Press, Inc, San Diego, 1991).
Hata, H. & Kato, M. Monoculture and mixed-species algal farms on a coral reef are maintained through intensive and extensive management by damselfishes. J. Exp. Mar. Biol. Ecol. 313, 285–296. (2004).
Ceccarelli, D. M., Jones, G. P. & McCook, L. J. Territorial damselfishes as determinants of the structure of benthic communities on coral reefs. Oceanogr. Mar. Biol. Annu. Rev. 39, 355–389 (2001).
Kaufman, L. The three spot damselfish: effects on benthic biota of Caribbean coral species. Proc. 3rd Int. Coral Reef Symp. 1, 559–564 (1977).
Hinds, P. A. & Ballantine, D. L. Effects of the Caribbean threespot damselfish, Stegastes planifrons (Cuvier), on algal lawn composition. Aquat. Bot. 27, 299–308 (1987).
Klumpp D. W. & Polunin, N. V. C. Partitioning among grazers of food resources within damselfish territories on a coral reef. J. Exp. Mar. Biol. Ecol. 125, 145–169 (1989).
Hata, H., Nishihira, M. & Kamura, S. Effects of habitat-conditioning by the damselfish Stegastes nigricans (Lacepede) on the community structure of benthic algae. J. Exp. Mar. Biol. Ecol. 280, 95–116 (2002).
Wilson, S. K. & Bellwood, D. R. Cryptic dietary components of territorial damselfishes (Pomacentridae, Labroidei). Mar. Ecol. Prog. Ser. 153, 299–310 (1997).
Wilson, S. K., Bellwood, D. R., Choat, J. H. & Furnas, M. J. Detritus in the epilithic algal matrix and its use by coral reef fishes. Oceangr. Mar. Biol. Annu. Rev. 41, 279–309 (2003).
Fricke, A., Teichberg, M., Beilfuss, S. & Bischof, K. Succession patterns in algal turf vegetation on a Caribbean coral reef. Bot. Mar. 54, 111–126 (2011).
Barott, K. L. & Rohwer, F. L. Unseen players shape benthic competition on coral reefs. Trends Microbiol. 20, 621–628 (2012).
Quan-Young, L. I. & Espinoza-Avalos, J. Reduction of zooxanthellae density, chlorophyll a concentration and tissue thickness of the coral Montastraea faveolata (Scleractinia) when competing with mixed turf algae. Limnol. Oceanogr. 51, 1159–1166 (2006).
Barott, K. L. et al. Microbial diversity associated with four functional groups of benthic reef algae and the reef building coral Montastraea annularis. Appl. Environ. Microbiol. 13, 1192–1204 (2011).
Smith, J. E. et al. Indirect effects of algae on coral: algae-mediated, microbe-induced coral mortality. Ecol. Lett. 9, 835–845 (2006).
Haas, A. F. et al. Effects of coral reef benthic primary producers on dissolved organic carbon and microbial activity. PLoS ONE 6, e27973 (2011).
Vega Thurber, R. et al. Macroalgae decrease growth and alter microbial community structure of the reef-building coral, Porites astreoides. PLoS ONE 7, e44246 (2012).
Sweet, M. J., Bythell, J. C. & Nugues, M. M. Algae as reservoirs for coral pathogens. PLoS ONE 8, 369717 (2013).
Arnold, S. N., Steneck, R. S. & Mumby, P. J. Running the gauntlet: inhibitory effects of algal turfs on the processes of coral recruitment. Mar. Ecol. Prog. Ser. 414, 91–105 (2010).
Casey, J. M., Choat, J. H. & Connolly, S. R. Coupled dynamics of territorial damselfishes and juvenile corals on the reef crest. Coral Reefs. 34, 1–11 (2015).
Barott, K. L. et al. Natural history of coral-algae competitors across a gradient of human activity in the Line Islands. Mar. Ecol. Prog. Ser. 460, 1–12 (2012).
Gleason, M. G. Coral recruitment in Moorea, French Polynesia: the importance of patch type and temporal variation. J. Exp. Mar. Biol. Ecol. 207, 79–101 (1996).
Glynn, P. W. & Colgan, M. W. Defense of corals and enhancement of coral diversity by territorial damselfishes. Proc. 6th Int. Coral Reef Symp. 2, 157–163 (1988).
Done, T. J., Dayton, P. K., Dayton, A. E. & Stegner, R. Regional and local variability in recovery of shallow coral reef communities. Coral Reefs. 9, 183–192 (1991).
Suefuji, M. & van Woesik, R. Coral recovery from the 1998 bleaching event is facilitated by Stegastes territories, Okinawa, Japan. Coral Reefs 20, 385–386 (2001).
White, J. S. S. & O’Donnell, J. L. Indirect effects of a key ecosystem engineer alter survival and growth of foundation coral species. Ecology 91, 3538–3548 (2010).
Gochfeld, D. J. Territorial damselfishes facilitate survival of corals by providing an associational defense against predators. Mar. Ecol. Prog. Ser. 398, 137–148 (2010).
Okubo, N., Taniguchi, H. & Motokawa, T. Successful methods for transplanting fragments of Acropora formosa and Acropora hyacinthus. Coral Reefs 24, 333–342 (2005).
Ainsworth, T. D. & Hoegh-Guldberg, O. Bacterial communities closely associated with coral tissues vary under experimental and natural reef conditions and thermal stress. Aquat. Biol. 4, 289–296 (2009).
van de Water, J. A. J. M. et al. Elevated seawater temperatures have a limited impact on the coral immune response following physical damage. Hydrobiologia. 10.1007/10750-015-2243-z. (2015).
Venables, W. N. & Ripley, B. D. Modern Applied Statistics with S 4th edn. (Springer, New York, 2002).
Caporaso, J. G. et al. QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 7, 335–336 (2010).
Lozupone, C. & Knight, R. UniFrac: a new phylogenetic method for comparing microbial communities. Appl. Environ. Microbiol. 71, 8228–8235 (2005).
Zuur, A. F., Ieno, E. N., Walker, N. J., Saveliev, A. A. & Smith, G. M. [Chapter 9: GLM and GAM for Count Data] Mixed Effects Models and Extensions in Ecology with R. (eds. Gail, M., Krickeberg, K., Samet, J. M., Tsiatis, A. & Wong, W. ) 227–228. (Springer, New York, 2009).
Bates, D., Maechler, M., Bolker, B. M. & Walker, S. lme4: Linear mixed-effects models using Eigen and S4. ArXiv e-print; submitted to J. Stat. Softw., Available at : http://arxiv.org/abs/1406.5823 (2014).
Myers, J. L., Sekar, R. & Richardson, L. L. Molecular detection and ecological significance of the cyanobacterial genera Geitlerinema and Leptolyngbya in black band disease of corals. Appl. Environ. Microbiol. 73, 5173 (2007).
Richardson, L. L. et al. Florida’s mystery coral-killer identified. Nature 392, 557–558 (1998).
Hube, B. From commensal to pathogen: stage- and tissue-specific gene expression of Candida albicans. Curr. Opinion Microbiol. 7, 336–341 (2004).
Sunagawa, S., Woodley, C. M. & Medina, M. Threatened corals provide underexplored microbial habitats. PLoS ONE 5, e9554 (2010).
Apprill, A., Hughen, K. & Mincer, T. Major similarities in the bacterial communities associated with lesioned a healthy Fungiidae corals. Environ. Microbiol. 15, 2063–2072 (2013).
McDaniel, L. D. et al. High frequency of horizontal gene transfer in the oceans. Science 330, 50 (2010).
McDaniel, L. D., Young, E. C., Ritchie, K. B. & Paul, J. H. Environmental factors influencing gene transfer agent (GTA) mediated transduction in the subtropical ocean. PLoS ONE 7, e43506 (2012).
Sekar, R., Kaczmarsky, L. T. & Richardson, L. L. Microbial community composition of black band disease on the coral host Siderastrea siderea from three regions of the wider Caribbean. Mar. Ecol. Prog. Ser. 362, 85–98 (2008).
Sunagawa, S. et al. Bacterial diversity and white plague disease-associated community changes in the Caribbean coral Montastrea faveolata. ISME J. 3, 512–521 (2009).
Pulcer-Rosario, G. & Randall, R. H. Preservation of rare coral species by transplantation and examination of their recruitment and growth. Bull. Mar. Sci. 41, 585–593 (1987).
Yap, H. T., Alvarez, R. M., Custodio III, H. M. & Dizon, R. M. Physiological and ecological aspects of coral transplantation. J. Exp. Mar. Biol. Ecol. 229, 69–84 (1998).
Ammar, M. S. A., Amin, E. M., Gundacker, D. & Mueller, W. E. G. One rational strategy for restoration of coral reefs: application of molecular biological tools to select sites for rehabilitation by asexual recruits. Mar. Poll. Bull. 40, 618–627 (2000).
Thornton, S. L., Dodge, R. E., Gilliam, D. S., DeVictor, R. & Cooke, P. Success and growth of corals transplanted to cement armor mat tiles in southeast Florida: Implications for reef restoration. Proc. 9th Int. Coral Reef Symp. 2, 955–962 (2000).
Soong, K. & Chen, T. Coral transplantation: Regeneration and growth of Acropora fragments in a nursery. Rest. Ecol. 11, 62–71 (2003).
Raymundo, L. J. & Maypa, A. P. Getting bigger faster: Mediation of size-specific mortality via fusion in juvenile coral transplants. Ecol. Appl. 14, 281–295 (2004).
Garrison, V. & Ward, G. Storm-generated coral fragments – a viable source of transplants for reef rehabilitation. Biol. Cons. 141, 3089–3100 (2008).
Geffen, Y., Ron, E. Z. & Rosenberg, E. Regulation of release of antibacterials from stressed scleractinian corals. FEMS Microbiol. Lett. 295, 103–109 (2009).
Littman, R. A., Bourne, D. G. & Willis, B. L. Responses of coral-associated bacterial communities to heat stress differ with Symbiodinium type on the same coral host. Mol. Ecol. 19, 1978–1990 (2010).
Meron, D. et al. Changes in coral microbial communities in response to a natural pH gradient. ISME J. 6, 1775–1785 (2012).
Jessen, C. et al. In-situ effects of eutrophication and overfishing on physiology and bacterial diversity of the Red Sea coral Acropora hemprichii. PLoS ONE 8, e62091 (2013).
Pantos, O. et al. The bacterial ecology of a plague-like disease affecting the Caribbean coral Montastrea annularis. Environ. Microbiol. 5, 370–382 (2003).
Pantos, O. & Bythell, J. C. Bacterial community structure associated with white band disease in the elkhorn coral Acropora palmata determined using culture-independent 16S rRNA techniques. Dis. Aquat. Org. 69, 79–88 (2006).
Harvell, D. et al. Coral disease, environmental drivers and the balance between coral and microbial associates. Oceanography 20, 172–195 (2007).
Kimes, N. E., Van Nostrand, J. D., Weil, E., Zhou, J. & Morris, P. J. Microbial functional structure of Montastrea faveolata, an important Caribbean reef-building coral, differs between healthy and yellow-band diseased colonies. Environ. Microbiol. 12, 541–556 (2010).
Sato, Y., Willis, B. L. & Bourne, D. G. Pyrosequencing-based profiling of archaeal and bacterial 16S rRNA genes identifies a novel archaeon associated with black band disease in corals. Environ. Microbiol. 15, 2994–3007 (2013).
Harriott, V. J. & Fisk, D. A. Coral transplantation as a reef management option. Proc. 6th Int. Coral Reef Symp. 2, 375–379 (1988).
Oren, U. & Benayahu, Y. Transplantation of juvenile corals: a new approach for enhancing colonization of artificial reefs. Mar. Biol. 127, 499–505 (1997).
Jaap, W. C. Coral reef restoration. Ecol. Engineer 15, 345–364 (2000).
Rinkevich, B. Conservation of coral reefs through active restoration measures: Recent approaches and last decade progress. Environ. Sci. Technol. 39, 4333–4342 (2005).
Yap, H. T. Local changes in community diversity after coral transplantation. Mar. Ecol. Prog. Ser. 374, 33–41 (2009).
Muko, S. & Iwasa, Y. Long-term effect of coral transplantation: Restoration goals and the choice of species. J. Theor. Biol. 280, 127–138 (2011).
Teplitski, M. & Ritchie, K. How feasible is the biological control of coral diseases? Trends Ecol. Evol. 24, 378–385 (2009).
Acknowledgements
We thank Lizard Island Research Station, K.D. Anderson, S.A. Blowes, S.J. Brandl, C. Heckathorn and M.O. Hoogenboom for field support. We also thank the Ecological Modelling Group at James Cook University for helpful comments that improved the manuscript. Funds were provided by the Australian Research Council Centre of Excellence for Coral Reef Studies and the Great Barrier Reef Marine Park Authority Science for Management Award. All work was carried out under the Great Barrier Reef Marine Parks Permit No: G11/34774.1.
Author information
Authors and Affiliations
Contributions
J.M.C., S.R.C. and T.D.A. conceived and designed the experiments. J.M.C. conducted the experiment, performed lab work and analyzed the data. All authors wrote the manuscript.
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Electronic supplementary material
Rights and permissions
This work is licensed under a Creative Commons Attribution 4.0 International License. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in the credit line; if the material is not included under the Creative Commons license, users will need to obtain permission from the license holder to reproduce the material. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/
About this article

Cite this article
Casey, J., Connolly, S. & Ainsworth, T. Coral transplantation triggers shift in microbiome and promotion of coral disease associated potential pathogens. Sci Rep 5, 11903 (2015). https://doi.org/10.1038/srep11903
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/srep11903
This article is cited by
-
Diversity and Distribution of Microbial Communities Associated with Reef Corals of the Malay Peninsula
Microbial Ecology (2023)
-
Bacterial communities associated with corals out-planted on the Great Barrier Reef are inherently dynamic over space and time
Marine Biology (2023)
-
Impacts of nursery-based propagation and out-planting on coral-associated bacterial communities
Coral Reefs (2022)
-
Variance of coral anti-pathogen defense in response to transplantation between coral- and macroalgal-dominated reefs
Coral Reefs (2022)
-
Differential Response of the Microbiome of Pocillopora acuta to Reciprocal Transplantation Within Singapore
Microbial Ecology (2022)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
