
Abstract
Wrinkle structures in rocks younger than the Permian-Triassic (P-Tr) extinction have been reported repeatedly in marine strata, but rarely mentioned in rocks recording land. Here, three newly studied terrestrial P-Tr boundary rock succession in North China have yielded diverse wrinkle structures. All of these wrinkles are preserved in barely bioturbated shore-shallow lacustrine siliciclastic deposits of the Liujiagou Formation. Conversely, both the lacustrine siliciclastic deposits of the underlying Sunjiagou Formation and the overlying Heshanggou Formation show rich bioturbation, but no wrinkle structures or other microbial-related structures. The occurrence of terrestrial wrinkle structures in the studied sections reflects abnormal hydrochemical and physical environments, presumably associated with the extinction of terrestrial organisms. Only very rare trace fossils occurred in the aftermath of the P-Tr extinction, but most of them were preserved together with the microbial mats. This suggests that microbial mats acted as potential oases for the surviving aquatic animals, as a source of food and oxygen. The new finds suggests that extreme environmental stresses were prevalent both in the sea and on land through most of the Early Triassic.
Similar content being viewed by others


Introduction
In ancient siliciclastic and carbonate depositional systems, the interaction between microbial activities and physical sedimentary processes can produce microbially related sedimentary structures. These were widespread during the Proterozoic and even extend back to the Early Archean1,2,3,4, but became restricted to highly stressed environments after the Cambrian4,5 because they were generally destroyed by burrowing organisms. In addition, occurrences of microbially related sedimentary structures provide the most valuable evidence for detecting early life and reestablishment of environments in the Precambrian6.
After the Cambrian, the most widespread microbially related sedimentary structures occur in shallow marine settings following the largest mass extinction of all time near the P-Tr boundary7,8,9. The P-Tr mass extinction event wiped out over 90% of marine species and most terrestrial vertebrate and plant species10. The diversity of marine invertebrates declined dramatically11 and bioturbation returned to Precambrian levels12. Microbial buildups and mats dominated the seabed after the P-Tr crisis, in the form of microbialites7,9 and wrinkle structures8,9. Cyanobacteria- dominated marine ecosystems lasted for about 5 Myr in the Early Triassic, linked to the depression of bioturbation and/or to long-term unusual environmental conditions9.
Current work suggests that the destruction of terrestrial ecosystems happened simultaneously with the decline in marine diversity during the P-Tr transition13,14. However, less is known about the specific changeover of terrestrial ecosystems during the P-Tr transition. So far, most attention has focused on tetrapods and plants15,16. Recent reviews indicate that tetrapods on land suffered a massive loss during the P-Tr crisis, as severe as that of life in the oceans, but the extinction seems to have been less profound for plants and insects17. Here we present new observations and investigations of wrinkle structures and trace fossils as well as their space relations in shallow-shore lacustrine siliciclastic deposits of Lower Triassic strata from North China that provides an exceptionally detailed record of terrestrial aquatic ecosystems during the P-Tr crisis.
Results
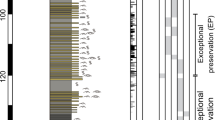
North China was located in the north tropical zone, between about 10°N and 20°N during the P-Tr transitional time (see Supplementary Fig. S1). Since the Middle Permian, North China had been largely land and rock successions are characterized by fluvial-lacustrine sedimentary facies. During the earliest Triassic, the central part of North China was dominated by east-west distributed lacustrine deposition, surrounded by dozens of lake deltas and flooding sandy plains (Supplementary Fig. S2b). The Upper Permian to Lower Triassic sedimentary sequence in North China is represented by the Shiqianfeng Group, which comprises, from oldest to youngest, the Sunjiagou, Liujiagou and Heshanggou formations. Wrinkle structures are observed in the Liujiagou Formation in the Dayulin, Sugou and Yuntouling sections, located within 100 km of each other in the central part of North China (Fig. 1).

Maps and columnar sections.
(a) Sketch map of China, showing Henan Province straddling the Yellow River (grey shading). (b) Locations of the studied sections, in Henan Province. (c) Stratigraphic sections showing lithologies and distributions of wrinkle structures; symbols (right) indicate major lithologies and fossils and sedimentary structures (the nine symbols indicated in light blue). Abbreviations: Fm, Formation; P, Permian; Tr, Triassic. D.L.C. created the figure using MapGIS6.7 and CorelDRAW14.
The lower to middle part of the Sunjiagou Formation is composed of fine-grained sandstones and thinly interbedded siltstones, which show rich and intensive bioturbation, all deposited in a shallow-shore lake environment. The upper part of the Sunjiagou Formation is composed of mudstones, siltstones and fine-grained sandstones. Typical Late Permian plant fossil elements assigned to Ullmannia occur at the upper part of the Sunjiagou Formation in the Dayulin and Sugou sections. The P-Tr boundary is identified at a horizon about 20 m below the top of the Sunjiagou Formation by the occurrence of the Lundbladispora-Aratrisporites-Taeniaesporites assemblage18, all of these representing typical earliest Triassic palynomorphs19. The Liujiagou Formation (100–150 m in thickness) consists of fine-grained sandstones with abundant ripple marks and cross beddings, bearing diverse wrinkle structures but infrequent bioturbation, deposited in a lake shore environment. Plant fossils assigned to Pleuromeia, a typical Early Triassic element20, occur in the upper part of the Liujiagou Formation in the Dayulin section and are sporadically distributed more widely within the Lower Triassic strata in North China21. In addition, oolitic beds accompanied with hummocky cross bedding, flat-pebble conglomerates and desiccation cracks with distinct geometries are observed in the Liujiagou Formation. The Heshanggou Formation (200–250 m in thickness) is mainly composed of purple siltstones and mudstones with diverse trace fossils, such as Planolites, Psilonichnus, Scoyenia, Skolithos and Taenidium, indicating a shallow-shore lake palaeoenvironment. The bioturbation is well developed in the Heshanggou Formation, with a maximum measured bioturbation index22 up to 5, but no microbial-related structures have been observed. The overlying Middle Triassic Ermaying Formation (450–600 m in thickness) consists of sandstones and mudstones with abundant invertebrate fossils and diverse trace fossils.
Four wrinkle structure types were recognized through studies of morphology and microscopic characteristics. Cross cutting wrinkle structures23 are preserved as bifurcating and frequently interconnected crests and intervening troughs on upper bedding surfaces of fine sandstones and one set of wrinkles cuts across another set at a low angle (Fig. 2a). The crests are 2–5 mm in height, 2–6 mm in width, irregular and round-topped. Microscopic features seen in longitudinal thin sections show 0.1–0.4 mm thick, wavy, crinkly, dark and opaque clay laminae on the surfaces of the wrinkle structures (Fig. 2d,h). Fine-grained quartz crystals are abundant in the surface laminae as floating grains and these differ from the underlying grains in size. The clay laminae and the floating quartz grains suggest that the microbial mat trapped sand grains from the surrounding environment, based upon comparative study between ancient and modern microbial mat growth1.

Photographs and thin section photomicrographs of wrinkle structures.
(a) Cross-cutting wrinkle structures. (b) Parallel wavy wrinkle structures. (c) Bulge-like wrinkle structure, with elongate to irregular bulges in close-up view (bottom left). (d) Thin section of wrinkle features in (a). (e,f) Thin section of the complete structure of the wrinkles. (g) Thin section of wrinkle features in Fig. 3a. (h-k) Close-up views of indicated portions of (d), (f) and (g). (l-n) SEM images showing the presence of the clay laminae and its detailed internal structure from (i-k). D.L.C. created this figure using CorelDRAW14.
Parallel wavy wrinkle structures are preserved as sinuously curved continuous crests, separated by parallel narrow troughs, extending over 15 cm or more on the upper surface of fine sandstone beds (Fig. 2b). The continuous crests of these structures are invariably flat-topped and the height of individual crests usually ranges from 1 to 2 mm and their spacing from 10 to 15 mm. Microscopic thin sections perpendicular to the structural trend show that the clay laminae are completely encased in sediment above and below, preserving the complete structure of the wrinkles (Fig. 2e,f,i). SEM images show that the quartz crystals are surrounded by clay laminae (Fig. 2l). This structure is considered to result from mat deformation, perhaps the crumpling of a disrupted and marginally detached mat24. Recent wave tank experiments suggest that microbial aggregates can form such wrinkle structures with morphological biosignatures at the sediment-water interface in wave-dominated environments25.
Bulge-like wrinkle structures are characterized by patches of elongate to irregular bulges measuring 2–5 mm in length on the upper surfaces of fine-grained sandstones (Fig. 2c). In appearance, bulges with similar geometry and orientation are preserved in discrete patches, presumably retained by covering microbial mats26. Preservation of the bulge-like wrinkles and their particular peripheries reflect subsurface morphological features developed beneath a microbial mat24 and suggest that at least two generations of microbial development occurred prior to burial.
“Old-elephant-skin textures”27 or briefly “elephant skin”28 wrinkle structures were found on the bedding surfaces of the fine-grained sandstones (Fig. 3a). The corrugated surface is characterized by an irregular and geometrically distinct polygonal texture. Individual networks are 3–10 mm in width, with a maximum vertical relief of 5 mm. Dark clay laminae overlying the surface have been observed by microscopic study of thin sections (Fig. 2g,j,k). In addition, SEM images show that the superposed clay laminae wrapped around the quartz grains (Fig. 2m,n). Previous studies suggested that this structure might form when mats provided a veneer between the lithologically identical overlying and underlying beds2.

Examples of trace fossils associated with mats.
(a) Back-filled tunnel cutting through biomat wrinkle structures in the Liujiagou Formation, indicating the trace maker may have mined the microbial mats for food resources. (b) Back-filled trace fossil associated with wrinkle structures (arrowed) in the Liujiagou Formation. Grey sketches in (a) and (b) are offset from the burrows and indicate their shape and size. D.L.C. created this figure using CorelDRAW14.
Discussion
Blooms of microbial-related sedimentary structures are usually caused by a combination of biotic and sedimentological factors, such as bioturbation, grazing pressure, nutrient levels and sediment dynamics. Modern studies show that the intensity of bioturbation directly determines the presence or absence of microbial mat communities29. The geological record suggests that microbial communities usually proliferated in the aftermath of mass extinctions in the post-Cambrian5,7,8,9, in both carbonate and siliciclastic settings. As the greatest crisis of the Phanerozoic, current studies show that the land ecosystem suffered marked biotic losses during the P-Tr mass extinction15,16,17. There is strong evidence for the absence of metazoans and of bioturbation in the Liujiagou Formation in North China and from the varying bioturbation index measured from the Dayulin section through the upper Sunjiagou Formation to the Ermaying Formation (Fig. 4). Decreased grazing pressure, coupled with low-level bioturbation following the P-Tr crisis, promoted the formation and preservation of microbial-related sedimentary structures. The abundant and diverse wrinkle structures in the Early Triassic of North China that we report here suggest the presence of microbe-dominated aquatic ecosystems in terrestrial facies as a response to the environmental crisis following the P-Tr mass extinction.

Variations in sedimentary structures, bioturbation index, ostracod volume, plants, ostracods and conchostracans from the Late Permian to Middle Triassic.
Wrinkle structures are most common in the Liujiagou Formation, but not present in the Sunjiagou, Heshangou and Ermaying formations. The bioturbation index was measured from the Dayulin section. Fossil data are from Supplementary Tables S1-4. The data on ostracod size is in Supplementary Table S5. Abbreviations: Di., Dienerian; Fm, Formation; Gr., Griesbachian; Ma, million years ago; Sm., Smithian. D.L.C. created this figure using CorelDRAW14.
Nutrient levels are significant in ecosystem construction. Studies of modern fresh waters demonstrate that microbial biomass responds positively to nutrient enrichment, with enhancement of the activities of extracellular enzymes and of metabolic rates30. Increased rates of terrestrial weathering have been proved near the P-Tr boundary31, which must have resulted in a significant increase in sediment fluxes32. Such large increases in sediment flux might have profoundly enhanced nutrient supply to rivers and oceans. Nutrient enrichment would promote the spread of sensitive microbial communities in both terrestrial (this study) and marine depositional systems7,9.
Physical sedimentary processes also play an important role in the formation of microbial mat-related sedimentary structures, according to the following sequence of events1,33: (1) binding, in which individual microbes actively assemble to form a microbial mat; (2) biostabilization, in which microbial mats escape destruction by erosion, but are remoulded to a different shape; (3) baffling and trapping, in which the mats accumulate sedimentary particles of a particular size on their surface; and (4) fossilization, in which the mats are buried by rapid pulses of fine-grained sediment deposition and preserved with atypical features in the rock record. Above all, extrinsic factors, including the size of the sediment and the effects of hydraulics and sediment dynamics, are closely linked with the formation of microbially-related sedimentary structures, among key physical sedimentary processes33. The occurrence of the oolitic beds, accompanied by hummocky cross beddings and flat-pebble conglomerates in the Liujiagou Formation in the studied sections, are similar to microbialites and oolites from the Buntsandstein Group in Germany, which are thought to have been deposited in an alkaline playa lake34, although other authors argued a marine origin35. The extensive occurrences of such sedimentary structures may imply strong water energy and potential geochemical anomalies in continental lakes worldwide, as another sedimentary response to the drastic upheavals in Early Triassic ecosystems and climates36.
Oceanic anoxia and lethal warming were thought to have been significant components of selective extinction during the P-Tr marine crisis37,38 and only those organisms with high tolerance levels to hypoxia and warming could survive in the mid-water refuge zone37. In lakes on land, the situation might have been worse. The rate of temperature increase would be greater on land than in the sea. Reduced atmospheric oxygen content38, elevated temperature39 and a large amount of organic input32 would dramatically decrease the solubility of oxygen and lead to hypoxic to anoxic lakes. This situation is in agreement with the finding of much-reduced bioturbation and the lack of body fossils of invertebrates in the early Early Triassic of North China (Fig. 4). Furthermore, our investigation of size variation in the terrestrial ostracod Darwinula from the Late Permian to Middle Triassic shows a sharp decrease during the P-Tr crisis (Fig. 4). The Lilliput effect40, well known among Early Triassic marine invertebrates41, also occurred among aquatic invertebrates on land, implying similarly stressed environments.
Finally, we identify evidence that certain lake-dwellers exploited the microbial mats as oases of oxygen and nutrients. This is based on the occurrence of rare trace fossils in the Liujiagou Formation, running parallel to the surface of the microbial mats (Fig. 3). The burrow cutting through the biomat might indicate that the trace maker may have mined the microbial mats for food resources (Fig. 3a). This evidence suggests that trace-making animals inhabited settings with microbial populations and may have mined microbially bound sediments for food resources, a phenomenon that has been mentioned in the study of the relationship between the earliest mobile animals and settings with high microbial populations in the Precambrian26,42. A modern analogue is a warm, hypoxic, hypersaline lagoon in Venezuela in which it was found43 that O2 levels in the cyanobacterial, photosynthesizing microbial mats are four times higher than in the overlying hypoxic water column and so these provide a suitable environment for animals to exploit biomat oxygen sources whilst foraging for food. Here, we hypothesize that survivors of the P-Tr mass extinction might have evolved in similar environments during the Early Triassic, effectively exploiting the abundant microbial mats, which acted as oases rich in oxygen and food within otherwise poorly oxygenated settings.
Methods
Sedimentary structures were recorded in detail by field observations in the three studied sections. A total of 133 samples were collected. All the features were photographed with a Canon EOS 7D digital camera. Cut slabs of rock and thin sections were produced using standard techniques. Sixty-five thin sections were made and these were examined and photographed using a LEICA-DM-750P microscope. Three thin sections were employed for ultramicroscopic studies under a HITACHI-SU8010 SEM with a iXRF EDS.
Additional Information
How to cite this article: Chu, D. et al. Early Triassic wrinkle structures on land: stressed environments and oases for life. Sci. Rep. 5, 10109; doi: 10.1038/srep10109 (2015).
References
Hagadorn, J. W. & Bottjer, D. J. Wrinkle structures: microbially mediated sedimentary structures common in subtidal siliciclastic settings at the Proterozoic-Phanerozoic transition. Geology 25, 1047–1050 (1997).
Schieber, J. et al. Atlas of Microbial Mat Features Preserved within the Siliciclastic Rock Record Elsevier: Amsterdam, 2007).
Noffke, N., Eriksson, K. A., Hazen, R. M., & Simpson, E. L. A new window into Early Archean life: Microbial mats in Earth’s oldest siliciclastic tidal deposits (3.2 Ga Moodies Group, South Africa). Geology 34, 253–256 (2006).
Hagadorn, J. W. & Bottjer, D. J. Restriction of a late Neoproterozoic biotope suspect-microbial structures and trace fossils at the Vendian-Cambrian transition. Palaios 14, 73–85 (1999).
Sheehan, P. M. & Harris, M. T. Microbialite resurgence after the Late Ordovician extinction. Nature 430, 75–78 (2004).
Noffke, N., Hazen, R. & Nhleko, N. Earth’s earliest microbial mats in a siliciclastic marine environment (2.9 Ga Mozaan Group, South Africa). Geology 31, 673–676 (2003).
Kershaw, S., Zhang, T. & Lan, G. A ?microbialite carbonate crust at the Permian-Triassic boundary in South China and its palaeoenvironmental significance. Palaeogeogr. Palaeoclimatol. Palaeoecol. 146, 1–18 (1999).
Pruss, S., Fraiser, M. & Bottjer, D. J. Proliferation of Early Triassic wrinkle structures: implications for environmental stress following the end-Permian mass extinction. Geology 32, 461–464 (2004).
Pruss, S. B., Bottjer, D. J., Corsetti, F. A. & Baud, A. A global marine sedimentary response to the end-Permian mass extinction: examples from southern Turkey and the western United States. Earth-Sci. Rev. 78, 193–206 (2006).
Erwin, D. H. The Great Paleozoic Crisis: Life and Death in the Permian Columbia Univ. Press: New York, 1993).
Alroy, J. et al. Phanerozoic trends in the global diversity of marine invertebrates. Science 321, 97–100 (2008).
Knaust, D. The end-Permian mass extinction and its aftermath on an equatorial carbonate platform: insights from ichnology. Terra Nova 22, 195–202 (2010).
Twitchett, R. J., Looy, C. V., Morante, R., Visscher, H. & Wignall, P. B. Rapid and synchronous collapse of marine and terrestrial ecosystems during the end-Permian biotic crisis. Geology 29, 351–354 (2001).
Shen, S. et al. Calibrating the end-Permian mass extinction. Science 334, 1367–1372 (2011).
Retallack, G. J. Permian-Triassic life crisis on land. Science 267, 77–80 (1995).
Benton, M. J., Tverdokhlebov, V. P. & Surkov, M. V. Ecosystem remodelling among vertebrates at the Permian-Triassic boundary in Russia. Nature 432, 97–100 (2004).
Benton, M. J. & Newell, A. J. Impacts of global warming on Permo-Triassic terrestrial ecosystems. Gondwana Res. 25, 1308–1337 (2014).
Ouyang, S. & Zhang, Z. Early Triassic palynological assemblage in Dengfeng, northwestern Henan. Acta Palaeontol. Sin. 21, 685–696 (1982).
Krassilov, V. & Karasev, E. Paleofloristic evidence of climate change near and beyond the Permian-Triassic boundary. Palaeogeogr. Palaeoclimatol. Palaeoecol. 284, 326–336 (2009).
Grauvogel-Stamm, L. & Ash, S. R. Recovery of the Triassic land flora from the end-Permian life crisis. C. R. Palevol. 4, 593–608 (2005).
Wang, Z. Recovery of vegetation from the terminal Permian mass extinction in North China. Rev. Palaeobot. Palynol. 91, 121–142 (1996).
Taylor, A. M. & Goldring, R. Description and analysis of bioturbation and ichnofabric. J. Geol. Soc. London 150, 141–148 (1993).
Banerjee, S. & Jeevankumar, S. Microbially originated wrinkle structures on sandstone and their stratigraphic context: Palaeoproterozoic Koldaha Shale, central India. Sediment. Geol. 176, 211–224 (2005)
Porada, H. & Bouougri, E. H. Wrinkle structures—a critical review. Earth-Sci. Rev. 81, 199–215 (2007).
Mariotti, G., Pruss, S. B., Perron, J. T. & Bosak, T. Microbial shaping of sedimentary wrinkle structures. Nat. Geosci. 7, 736–740 (2014).
Sarkar, S., Banerjee, S., Samanta, P. & Jeevankumar, S. Microbial mat-induced sedimentary structures in siliciclastic sediments: examples from the 1.6 Ga Chorhat Sandstone, Vindhyan Supergroup, MP, India. J. Earth Syst. Sci. 115, 49–60 (2006).
Runnegar, B. & Fedonkin, M. in The Proterozoic Biosphere: a Multidisciplinary Study (eds Schopf, J. & Klein, C. ) 369–388 Cambridge Univ. Press: Cambridge, 1992).
Gehling, J. G. Microbial mats in terminal Proterozoic siliciclastics: Ediacaran death masks. Palaios 14, 40–57 (1999).
Seckbach, J. & Oren, A. Microbial Mats: Modern and Ancient Microorganisms in Stratified Systems Springer, Berlin, 2010).
Sánchez-Carrillo, S., Reddy, K. R., Inglett, K. S., Álvarez-Cobelas, M. & Sánchez-Andrés, R. in Eutrophication: Causes, Consequences and Control (eds Ansari, A. A. & Gill, S. S. ) 203–222 Springer, Berlin, 2014).
Newell, A. J., Tverdokhlebov, V. P. & Benton, M. J. Interplay of tectonics and climate on a transverse fluvial system, Upper Permian, Southern Uralian Foreland Basin, Russia. Sediment. Geol. 127, 11–29 (1999).
Algeo, T. J. & Twitchett, R. J. Anomalous Early Triassic sediment fluxes due to elevated weathering rates and their biological consequences. Geology 38, 1023–1026 (2010).
Noffke, N. & Awramik, S. Stromatolites and MISS—Differences between relatives. GSA Today 23, 4–9 (2013).
Paul, J. & Peryt, T. M. Kalkowsky’s stromatolites revisited (Lower Triassic Buntsandstein, Harz Mountains, Germany). Palaeogeogr. Palaeoclimatol. Palaeoecol. 161, 435–458 (2000).
Weidlich, O. PTB mass extinction and earliest Triassic recovery overlooked? New evidence for a marine origin of Lower Triassic mixed carbonate-siliciclastic sediments (Rogenstein Member), Germany. Palaeogeogr. Palaeoclimatol. Palaeoecol. 252, 259–269 (2007).
Kidder, D. L. & Worsley, T. R. Causes and consequences of extreme Permo-Triassic warming to globally equable climate and relation to the Permo-Triassic extinction and recovery. Palaeogeogr. Palaeoclimatol. Palaeoecol. 203, 207–237 (2004).
Song, H. et al. Anoxia/high temperature double whammy during the Permian-Triassic marine crisis and its aftermath. Sci. Rep. 4, 4132 (2014).
Berner, R. A., VandenBrooks, J. M. & Ward, P. D. Oxygen and evolution. Science 316, 557–558 (2007).
Sun, Y. et al. Lethally hot temperatures during the early Triassic greenhouse. Science 338, 366–370 (2012)
Urbanek, A. Biotic crises in the history of Upper Silurian graptoloids: a palaeobiological model. Hist. Biol. 7, 29–50 (1993).
Twitchett, R. J. The Lilliput effect in the aftermath of the end-Permian extinction event. Palaeogeogr. Palaeoclimatol. Palaeoecol. 252, 132–144 (2007).
Seilacher, A., Buatois, L. A. & Mangano, M. G. Trace fossils in the Ediacaran-Cambrian transition: behavioural diversification, ecological turnover and environmental shift. Palaeogeogr. Palaeoclimatol. Palaeoecol. 227, 323–356 (2005).
Gingras, M. et al. Possible evolution of mobile animals in association with microbial mats. Nat. Geosci. 4, 372–375 (2011).
Acknowledgements
This study was supported by the 973 Program (National Basic Research Program of China; 2011CB808800), the National Natural Science Foundation of China (41272372, 41302271, 41172312), the 111 Project (B08030), BGEG (GBL11202, GBL11302) and the Fundamental Research Funds for the Central Universities (CUG130407, CUG120117, CUG1410491T01, CUGL140828).
Author information
Authors and Affiliations
Contributions
J.N.T. and H.J.S. led the analysis. D.L.C. and H.J.S. wrote the initial manuscript. M.J.B. and D.J.B. provided substantial comments and editorial revisions to the manuscript. H.Y.S. worked closely with L.T. on all aspects of the analysis and writing. All authors contributed to the discussions.
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Electronic supplementary material
Rights and permissions
This work is licensed under a Creative Commons Attribution 4.0 International License. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in the credit line; if the material is not included under the Creative Commons license, users will need to obtain permission from the license holder to reproduce the material. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/
About this article

Cite this article
Chu, D., Tong, J., Song, H. et al. Early Triassic wrinkle structures on land: stressed environments and oases for life. Sci Rep 5, 10109 (2015). https://doi.org/10.1038/srep10109
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/srep10109
This article is cited by
-
Carboniferous and Permian integrative stratigraphy and timescale of North China Block
Science China Earth Sciences (2022)
-
Altered fluvial patterns in North China indicate rapid climate change linked to the Permian-Triassic mass extinction
Scientific Reports (2019)
-
Boreal earliest Triassic biotas elucidate globally depauperate hard substrate communities after the end-Permian mass extinction
Scientific Reports (2016)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
