
Abstract
Willows (Salix) and poplars (Populus) are dioecious plants in Salicaceae family. Sex chromosome in poplar genome was consistently reported to be associated with chromosome XIX. In contrast to poplar, this study revealed that chromosome XV was sex chromosome in willow. Previous studies revealed that both ZZ/ZW and XX/XY sex-determining systems could be present in some species of Populus. In this study, sex of S. suchowensis was found to be determined by the ZW system in which the female was the heterogametic gender. Gene syntenic and collinear comparisons revealed macrosynteny between sex chromosomes and the corresponding autosomes between these two lineages. By contrast, no syntenic segments were found to be shared between poplar's and willow's sex chromosomes. Syntenic analysis also revealed substantial chromosome rearrangements between willow's alternate sex chromatids. Since willow and poplar originate from a common ancestor, we proposed that evolution of autosomes into sex chromosomes in these two lineages occurred after their divergence. Results of this study indicate that sex chromosomes in Salicaceae are still at the early stage of evolutionary divergence. Additionally, this study provided valuable information for better understanding the genetics and evolution of sex chromosome in dioecious plants.
Similar content being viewed by others
Introduction
Sex determination has the intriguing aspect in evolutionary and developmental biology. In contrast to the more completed sex chromosome evolution in animals, sex chromosomes in many plants are still at different evolutionary stages1,2,3,4 and thus afford the opportunity of investigating the early stage of sex chromosome evolution. It is believed that sex chromosomes originate from a pair of autosomes5,6 and the sex-determining systems in dioecious plants almost certainly evolved independently from ancestral hermaphrodites that lacked sex chromosomes2,3. Only about 4% of higher plants show full dioecism, with individuals of separate sexes7,8,9.
Salicaceae is a family of dioecious woody plants and the male or female flowers are arranged in morphologically different catkins (Figure 1) on the male or female trees10. Salix and Populus are the sister genera in Salicaceae (Figure 2). Genome analysis revealed that these two lineages originated from a common paleotetrapolyploid ancestor11,12,13. The chromosomes of Salicaceae are typically metacentric and small14,15. Based on cytological studies, there is no evidence of morphologically differentiated sex chromosomes in the Salicaceae species16,17,18. Various mechanisms have been proposed to explain the expression of gender in Salicaceae. Alström-Rapaport et al. proposed a multi-locus sex determination in the Salix and that the presence of sex chromosomes was unlikely19. In contrast, Semerikov et al. reported that a single locus governed the sex determination and the female was the heterogametic gender in basket willow20. More recently, several studies revealed that there was a single locus located on chromosome XIX of Populus, with a peritelomeric localization in members of the Aigeiros subgenera21,22 and a centromeric localization in subgenera of Leuce23,24,25,26, appeared to contain a gene (genes) that controlled gender determination. Multiple lines of evidence suggested that poplar's chromosome XIX was in the process of evolving into an incipient sex chromosome22,27. The sex-determining region in poplar shows some interesting features, including location of gender locus, highly divergent haplotypes, severe recombination suppression, distinctive pattern of sRNA occurrence and significantly fewer single nucleotide polymorphisms (SNP) than the rest of Populus genome11,22,27. Mapping studies on P. deltoides22 and P. alba26 revealed that sex determination occurred through a ZW system in which the female was the heterogametic gender22. However, the male sex was reported as the heterogametic gender in P. nigra21 and P. tremuloides23,24,25. Thus, it is possible that both ZZ/ZW (female heterogamety) and XX/XY (male heterogamety) sex-determining systems could be present in some members of the family Salicaceae27. It has been confirmed that the sister genera of Salix and Populus originate from a common ancestor11,12,13. Since different gender determining systems probably evolve separately and quite recently in species of this family, Salicaceae is a desirable system to study the genetics and evolution of sex chromosomes in dioecious plants.

Flowers of the female and male trees in Salicaceae species.
On willow and poplar, the male or female flowers are separately arranged in morphologically different catkins on the male or female trees. Photos taken by Tongming Yin and Jing Hou.

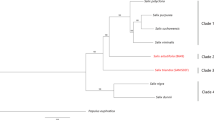
Phylogenetic tree of selected plant species.
The phylogenetic tree was constructed with 1,881 single-copy genes on 4-fold degenerate sites. The branch length represents the neutral divergence rate. The posterior probabilities (credibility of the topology) for inner nodes are all 100%.
S. suchowensis is an early flowering shrub willow that belongs to subgenus Vetrix28. Recently, its genome has been sequenced and publically available13. To gain further insight into the origin and evolution of sex chromosomes in Salicaceae species, we identified the sex chromosome in S. suchowensis, as well as compared the sex chromosomes between the sister genera of Salix and Populus.
Results and Discussion
To map the gender locus of S. suchowensis, we established a large mapping pedigree in which the sequenced individual was the maternal parent. Among the progeny, 374 individuals were randomly selected for mapping and locating the gender locus in the willow genome. Investigation of sex phenotype confirmed constant gender for all of the mapping progeny across three locations in two continuous years. Among these individuals, 183 were female, 188 were male and 3 remained sexually immature. Statistical analysis indicated that segregation of gender fit for the expected 1:1 Mendelian segregation ratio (χ2 = 0.067, where (χ2 = 3.84 for α ≤ 0.05). Subsequently, genetic maps were separately constructed for the maternal and paternal parents by using amplified fragment length polymorphism (AFLP) markers and the pseudo-testcross strategy29. Totally, 1,137 1:1 segregating markers were generated by 90 primer combinations with the 374 mapping progeny. Among these markers, 494 testcross markers from the maternal parent were mapped into 266 bins distributed on 19 linkage groups, covering a genetic distance of 1924.5 cM (Supplementary Figure 1); Alternatively, 549 testcross loci from the male were assigned into 263 bins on 19 linkage groups, spanning a genetic distance of 2223.8 cM (Supplementary Figure 2). The coverage of the female and male maps was estimated to be 99.99% and 99.97% at 20 cM of a marker, respectively. Thus, the established maps achieved nearly complete genome coverage, with linkage group numbers equaled to the 19 haploid chromosome numbers in willow. Based on the established genetic maps, the gender locus was mapped as a 1:1 segregating morphological marker. Mapping results showed that gender locus could only be mapped on the maternal map, but was unmappable on the paternal map, indicating that the female was the heterogametic gender in willow, which was in agreement with the findings in P. deltoides22 and P. alba26 that revealed sex determination occurred through a ZW system, in which females are heterogametic22. It was also noteworthy that marker density in the vicinity of the gender locus (Supplementary Figure 1) was significantly higher than that expected by chance alone (P ≤ 0.0001 with a Poisson calculator), which indicated severe recombination suppression around the gender locus. Recombination suppression has been recognized as a critical mechanism that triggered the divergence of the alternate sex chromosomes2,22,30,31.
To map the willow sequence scaffolds derived by Dai et al.13 along each chromosome of the willow genome, we carried out fine mapping with SNP markers that were generated by resequencing a subset of 80 progeny from the 374 mapping individuals. AFLP markers on the established maps were used as landmarks to anchor the SNP markers into the corresponding marker bins. In total, 2,548 testcross SNP markers from the maternal parent and 3,772 testcross SNP markers from the paternal parent were integrated into the female (Supplementary Figure 3) and male (Supplementary Figure 4) maps, respectively. Subsequently, SNP markers in each linkage group were used to map the willow sequence scaffolds derived by Dai et al.13 along the corresponding chromosome.
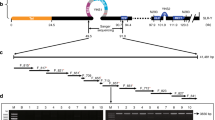
In total, 432 sequence scaffolds (covering a physical length of 194.1 Mb) and 630 sequence scaffolds (covering a physical length of 206.2 Mb) were mapped based on the female (Figure 3a) and male (Figure 3b) maps, respectively. There remained 7,009 unmapped sequence scaffolds larger than 2 kb, covering a total physical length of 38.4 Mb. Chromosome identities of the willow genome were designated by blasting willow sequences containing the mapped SNPs against the P. trichocarpa genome sequences. It was found that the linkage group containing the gender locus was chromosome XV in the willow genome (Figure 3a). This chromosome is an autosome in poplars. Thus, the willow's sex chromosome corresponds to a poplar's autosome. Previous studies revealed that chromosome XIX was the sex chromosome in poplars21,22,23,24,25,26, by contrast, chromosome XIX was identified as an autosome in willow.

Anchoring the willow sequence scaffolds along each chromosome in the genomes of Salix suchowensis.
(a) anchor with the female map (b) anchor with the male map. Note: willow sequence scaffolds were obtained from Dai et al.'s study13. Sequence scaffolds were mapped according to the integrated SNP markers on the maternal genetic map of S. suchowensis. The orange bars on the left of each chromosome represented the linkage groups and the discrete blue bars on the right represented the anchored sequence scaffolds. The mapped scaffolds were separated with evenly sized spaces, which did not represent the actual sizes of the uncaptured gaps. Chromosome identities were designated based on syntenic relationship between the willow's and poplar's chromosomes. As shown in Figure 3a, gender locus was mapped onto scaffold_64 on chromosome XV in the female S. suchowensis and markers cosegregated with gender were indicated with red lines.
On willow's chromosome XV and chromosome XIX, 18 and 40 sequence scaffolds were mapped, respectively, along the corresponding chromosome based on the maternal map (Figure 3a). Because the sequenced individuals of both P. trichocarpa and S. suchowensis were females11,13, we first conducted syntenic analysis between sex chromosomes and the corresponding autosomes in these two lineages based on the willow's female map. Syntenic analysis revealed high collinearity on chromosome XIX (Figure 4a) and on chromosome XV (Figure 4b) between willow and poplar. In P. trichocarpa, complex haplotype divergence was observed between its alternate sex chromatids11,27. To explore whether the Z and W sex chromatids diverged with each other in willow, we examined the sequence scaffolds anchored on willow's chromosome XV based on the male map, which included 45 sequence scaffolds (Figure 3b). Contrary to the consistency revealed based on the female map (Figure 4b), syntenic analysis based on the male map revealed substantial chromosome rearrangements (Figure 4c), suggesting the occurrence of significant chromosome divergence between the Z and W chromatids in willow. We further compared willow's sex chromosome (XV) with poplar's sex chromosome (XIX) and found that sex chromosomes between these two lineages shared no syntenic chromosome segments (Figure 4d), indicating that sex chromosomes of poplar and willow originated from different ancestral chromosomes. Since sex chromosomes act as autosomes in the alternate genera of Salicaceae, we propose that turnover of autosomes into sex chromosomes occurred after the divergence of Salix and Populus.

Syntenic analysis between sex chromosomes and the corresponding autosomes across Populus trichocarpa and Salix suchowensis.
(a) synteny between poplar's and willow's chromosome XIX based on the female map. (b) synteny between poplar's and willow's chromosome XV based on the female map. (c) synteny between poplar's and willow's chromosome XV based on the male map. (d) synteny between poplar's chromosome XIX and willow's chromosome XV based on the female map. Note: Bar on the top of each chromosome pair represents poplar's chromosome. Bar at the bottom corresponds to the anchored sequence scaffolds on willow's chromosome. Only sequence scaffolds containing coding genes were included in this analysis. The red portions of the chromosome bars represent the sex-determining regions.
Because evidence indicated that chromosome XV and chromosome XIX separately evolved into sex chromosomes after the divergence of willow and poplar, we proposed that turnover of different autosomes into sex chromosomes might be due to mobile of the sex-determining gene from chromosome XIX to chromosome XV in willow. Alternatively, the sex-determining gene may have translocated from chromosome XV to chromosome XIX in poplar. Therefore, there might be homologous sex-determining genes within the sex-determining regions between willow and poplar. Mapping of SSR markers revealed that the gender locus was in between SSR markers SSR151 (151,741 bp on scaffold 64) and SSR893 (893,817 bp on scaffold 64) (Figure 4b). In P. nigra21 and P. deltoides22, the close relatives to P. trichocarpa (the sequenced poplar species), gender locus was consistently mapped to the peritelomeric region upper the position of SSR marker O_206 (Figure 4a). We subsequently searched the homologous genes shared by these two regions between willow and poplar and nine willow genes were found to have a total of 52 homologous genes in the sex-determining region of P. trichocarpa. However, all these poplar genes had homologous genes in other regions of the poplar genome and none of these genes were found to be involved in sex determination based on gene annotation (Supplementary Table 1).
In recent studies, RNA-based sex-determining mechanisms have been found to play a significant role in higher plants27,32,33,34. For example, the tasselseed4 miRNA, i.e., miR172, was found to be involved in the sex determination of the male inflorescence in maize32,33. Interestingly, the miR172 family was conserved in Populus35 and located on the peritelomeric end of chromosome XIX27. We searched the homolgous sequences of miR172 in willow and none of them appeared in the sex-determining region. More recently, Akagi et al. reported that a sex-determining candidate (OGI) encoded a small RNA targeting a feminizing gene (MeGI) in persimmons (Diospyros lotus) and the encoded small RNA acted as a sex determinant in this plant34. Search homologous genes of OGI and MeGI in poplar and willow showed that no homologous genes were found to be distributed on the sex chromosomes in either lineage (Supplementary Table 2). With all of these efforts, we have not identified the likely sex determinants in Salicaceae. The involvement of small RNAs in sex determination might explain why they have been difficult to identify to date34. RNA-based sex-determining mechanisms maybe present in Salicaceae species as well, thus small RNAs will be the primary focus in our future studies.
In plants, species with heterogametic females are less common36. Therefore, Salix species should represent a unique system for studying dioecy evolution in flowering plants. Although the sex-determining gene (genes) has not been identified with the current data, this work provides unique insight into the sex determination in Salicaceae and also sheds new light on understanding the genetics and evolution of sex chromosome in dioecious plants.
Methods
Plant materials
An F1 mapping pedigree with 2,546 progeny was established in 2010. The maternal parent of this pedigree was described by Dai et al.13, whereas the paternal parent was a two-year-old S. suchowensis collected from Linshu, Shandong Province of China. To map the gender locus, 374 offspring were randomly selected and cuttings of the selected individuals were planted at three different locations in China: Sihong and Lishui in Jiangsu Province, as well as Shishou in Hubei Province. The field trials were established in 2012 and three replicates were planted for each progeny at each location.
Map construction
AFLP genotyping was conducted as described by Yin et al.37. GeneMapper Software (Version 3.7, Applied Biosystems) was used to score the amplicons in range of 50 ~ 500 bp. Map construction, genome length estimation and map coverage calculation were performed following the description in Yin et al.37. A Poisson probability test was conducted following the description in Remington et al.38. The map charts were produced with MapChart 2.139.
Generating SNP markers
SNP markers were generated with a subset of 80 progeny from the 374 mapping individuals. The combination of two restriction enzymes, EcoRI and MseI (Takara Inc.), were used to reduce the complexity of the willow genome. For PCR amplification, the EcoRI primer had no selective nucleotide and the MseI primer had a “G” selective nucleotide. The PCR amplification was conducted as described by Yin et al.37.
The sequencing library was constructed according to the manufacturer's standard protocol (Illumina, Inc.). Library quantification was carried out with a Quant-iTTM PicoGreen dsDNA Kit (Invitrogen, Inc.) and library qualify was evaluated with an Agilent 2100 Bioanalyzer (Agilent, Inc.). For each lane of a flow cell, 12 index-labeled samples were multiplexed, which enabled the sequencing of 84 samples (80 progeny and two replicates of the parents) in a full run and the eighth lane was used for the control. Sequencing was performed on an Illumina HiSeq 2000 (Illumina, Inc.) at Nanjing Agricultural University following the manufacturer's protocols (Illumina, Inc.).
Raw sequencing data generated by the Illumina platform were analyzed with an integrated bioinformatics pipeline. The SAMTOOLS (http://samtools.sourceforge.net)40 and Varscan (http://varscan.source-forge.net) were combined to call the SNPs with default parameters41.
Anchoring the sequence scaffolds
AFLP markers on the established genetic maps were used to pull out tightly linked SNP markers with an LOD threshold ≥10 and these SNPs were assigned to the corresponding marker bins that showed the strongest linkage on each map. Subsequently, the mapped SNPs were used to anchor willow sequence scaffolds that were produced by Dai et al.13. Finally, willow sequences containing these SNPs were blasted against the P. trichocarpa genome sequences (ftp://ftp.jgi-psf.org/pub/compgen/phytozome/v9.0/Ptrichocarpa/) to designate the chromosome identities for the obtained linkage groups on the male and female maps.
Synteny and collinearity analyses
Genome sequences, Gff3 file and the protein sequences of P. trichocarpa were retrieved from JGI website (ftp://ftp.jgi-psf.org/pub/compgen/phytozo- me/v9.0/Ptrichocarpa/). The corresponding information for willow was downloaded from the S. suchowensis website (115.29.234.170/willow). Synteny and collinearity analyses were performed with bioinformatics toolkit of MCScanX42. First, coding genes in willow and poplar genome were blasted by using BLASTP43 with an e-value cutoff of 1e−10, which produced a file in blast8 format. Then, an in-house perl script was used to filter the gff3 files and reformat the files to make them compatible with MCScanX. Finally, MCScanX produced the collinearity results, with the following parameters: −k 50, −m 25, −e 1e−5, where −k is the match score, −m is the maximum gaps and −e is the e value cutoff. Dual_synteny_plotter in the MCScanX package was used to draw the collinearity charts.
References
Jamsari, A., Nitz, I., Reamon-Büttner, S. M. & Jung, C. BAC-derived diagnostic markers for sex determination in asparagus. Theor. Appl. Genet. 108, 1140–1146 (2004).
Liu, Z. Y. et al. A primitive Y chromosome in papaya marks incipient sex chromosome evolution. Nature 427, 348–352 (2004).
Filatov, D. A. Evolutionary history of Silene latifolia sex chromosomes revealed by genetic mapping of four genes. Genetics 170, 975–979 (2005).
Alexandrov, O. S., Divashuk, M. G., Yakovin, N. A. & Karlov, G. I. Sex chromosome differentiation in Humulus japonicus Siebold & Zuccarini, 1846 (Cannabaceae) revealed by fluorescence in situ hybridization of subtelomeric repeat. Comp. Cytogenet. 6, 39–247 (2012).
Smith, C. A. & Sinclair, A. H. Sex determination: insights from the chicken. Bioessays 26, 120–132 (2004).
Carvalho, A. B. & Clark, A. G. Y chromosome of D. pseudoobscura is not homologous to the ancestral Drosophila Y. Science 307, 108–110 (2005).
Lloyd, D. G & Webb, C. J. Secondary sex characters in plants. Bot. Rev. 43, 177–216 (1977).
Guttman, D. S. & Charlesworth, D. An X-linked gene with a degenerate Y-linked homologue in a dioecious plant. Nature 393, 263–266 (1998).
Heslop-Harrison, J. S. & Schwarzacher, T. Organisation of the plant genome in chromosomes. Plant J. 66, 18–33 (2011).
Rechinger, K. H. Salix taxonomy in Europe-problems, interpretations, observations. Proceedings of the Royal Society of Edinburgh. Section B. Biological Sciences 98, 1–12 (1992).
Tuskan, G. A. et al. The genome of black cottonwood, Populus trichocarpa (Torr. & Gray). Science 313, 1596–1604 (2006).
Berlin, S. et al. High-density linkage mapping and evolution of paralogs and orthologs in Salix and Populus. BMC Genomics 11, 129 (2010).
Dai, X. G. et al. The willow genome and divergent evolution from poplar after the common genome duplication. Cell Res. 24, 1274–1277 (2014).
Blackburn, K. B. & Harrison, J. W. H. A preliminary account of the chromosomes and chromosome behavior in the Salicaceae. Ann. Bot. 38, 361–378 (1924).
Islam-Faridi, M. N., Nelson, C. D., DiFazio, S. P., Gunter, L. E. & Tuskan, G. A. Cytogenetic analysis of Populus trichocarpa-ribosomal DNA, telomere repeat sequence and marker-selected BACs. Cytogenet. Genome Res. 125, 74–80 (2009).
Peto, F. H. Cytology of poplar species and natural hybrids. Can. J. Forest. Res. 16, 445–455 (1938).
Van Dillewijn, C. Zytologische Studien in der Gattung Populus L. Genetica 22, 131–182 (1940).
Van Buijtenen, J. P. & Einspahr, D. W. Note on the presence of sex chromosomes in Populus tremuloides. Bot. Gaz. 121, 60–61 (1959).
Alström-Rapaport, C., Lascoux, M. & Gullberg, U. Sex determination and sex ratio in the dioecious shrub Salix viminalis L. Theor. Appl. Genet. 94, 493–497 (1997).
Semerikov, V. et al. Genetic mapping of sex-linked markers in Salix viminalis L. Heredity 91, 293–299 (2003).
Gaudet, M. et al. Genetic linkage maps of Populus nigra L. including AFLPs, SSRs, SNPs and sex trait. Tree Genet. Genomes 4, 25–36 (2008).
Yin, T. M. et al. Genome structure and emerging evidence of an incipient sex chromosome in Populus. Genome Res. 18, 422–430 (2008).
Markussen, T., Pakull, B. & Fladung, M. Positioning of sex correlated markers for Populus in an AFLP- and SSR-marker based genetic map of Populus tremula × tremuloides. Silvae Genet. 56, 180–184 (2007).
Pakull, B., Groppe, K., Meyer, M., Markussen, T. & Fladung, M. Genetic linkage mapping in aspen (Populus tremula L. and Populus tremuloides Michx.). Tree Genet. Genomes 5, 505–515 (2009).
Pakull, B. et al. Genetic mapping of linkage group XIX and identification of sex-linked SSR markers in a Populus tremula× Populus tremuloides cross. Can. J. Forest. Res. 41, 245–253 (2011).
Paolucci, I. et al. Genetic linkage maps of Populus alba L. and comparative mapping analysis of sex determination across Populus species. Tree Genet. Genomes 6, 863–875 (2010).
Tuskan, G. A. et al. The obscure events contributing to the evolution of an incipient sex chromosome in Populus: a retrospective working hypothesis. Tree Genet. Genomes 8, 559–571 (2012).
Smart, L. B. & Cameron, K. D. Genetic improvement of willow (Salix spp.) as a dedicated bioenergy crop. In: Vermerris W editors. Genetic improvement of bioenergy crops. New York: Springer (2008).
Grattapaglia, D. & Sederoff, R. R. Genetic-linkage maps of Eucalyptus grandis and Eucalyptus urophylla using a pseudotestcross-mapping strategy and RAPD markers. Genetics 137, 1121–1137 (1994).
Ohno, S. Sex chromosomes and sex-linked genes. Berlin: Springer (1967).
Nei, M. Linkage modification and sex difference in recombination. Genetics 63, 681–699 (1969).
Chuck, G., Meeley, R., Irish, E., Sakai, H. & Hake, S. The maize tasselseed4 microRNA controls sex determination and meristem cell fate by targeting Tasselseed6/indeterminate spikelet1. Nat. Genet. 39, 1517–1521 (2007).
Banks, J. MicroRNA, sex determination and floral meristem determinacy in maize. Genome Biol. 9, 204 (2008).
Akagi, T. et al. A Y-chromosome–encoded small RNA acts as a sex determinant in persimmons. Science 346, 646–650 (2014).
Barakat, A., Wall, P. K., Diloreto, S., Depamphilis, C. W. & Carlson, J. E. Conservation and divergence of microRNAs in Populus. BMC Genomics 8, 481–496 (2007).
Charlesworth, D. Plant sex chromosome evolution. J. Exp. Bot. 64, 405–420 (2013).
Yin, T. M., DiFazio, S. P., Gunter, L. E., Riemenschneider, D. & Tuskan, G. A. Large-scale heterospecific segregation distortion in Populus revealed by a dense genetic map. Theor. Appl. Genet. 109, 451–463 (2004).
Remington, D. L., Whetten, R. W., Liu, B. H. & O′Malley, D. M. Construction of an AFLP genetic map with nearly complete genome coverage in Pinus taeda. Theor. Appl. Genet. 98, 1279–1292 (1999).
Voorrips, R. E. MapChart: software for the graphical presentation of linkage maps and QTLs. J. Hered. 93, 77–78 (2002).
Li, H. et al. The sequence alignment/map format and SAMtools. Bioinformatics 25, 2078–2079 (2009).
Koboldt, D. C. et al. VarScan: variant detection in massively parallel sequencing of individual and pooled samples. Bioinformatics 25, 2283–2285 (2009).
Wang, Y. P. et al. MCScanX: a toolkit for detection and evolutionary analysis of gene synteny and collinearity. Nucleic Acids Res 40, e49–e49 (2012).
Altschul, S. F., Gish, W., Miller, W., Myers, E. W. & Lipman, D. J. Basic local alignment search tool. J. Mol. Biol. 215, 403–410 (1990).
Acknowledgements
This work was supported by the National Basic Research Project (2012CB114505), the Natural Science Foundation of China (31125008, 31400564) and Jiangsu Province (BK20130968). It was also enabled by the Program for Innovative Research Team (the Educational Department of China, the Universities of Jiangsu Province), the Doctorate Fellowship Foundation and PAPD (Priority Academic Program Development) program at Nanjing Forestry University.
Author information
Authors and Affiliations
Contributions
J.H., D.Z. and L.F. conducted the experiments. J.H., N.Y., D.Z. and X.D. performed the data analysis. J.H. and Y.C. prepared the manuscript. T.Y. conceived this work and critically reformulated the manuscript. All authors have read and approved the final manuscript.
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Electronic supplementary material
Supplementary Information
Supplementary information
Rights and permissions
This work is licensed under a Creative Commons Attribution 4.0 International License. The images or other third party material in this article are included in the article's Creative Commons license, unless indicated otherwise in the credit line; if the material is not included under the Creative Commons license, users will need to obtain permission from the license holder in order to reproduce the material. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/
About this article

Cite this article
Hou, J., Ye, N., Zhang, D. et al. Different autosomes evolved into sex chromosomes in the sister genera of Salix and Populus. Sci Rep 5, 9076 (2015). https://doi.org/10.1038/srep09076
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/srep09076
This article is cited by
-
The male-heterogametic sex determination system on chromosome 15 of Salix triandra and Salix arbutifolia reveals ancestral male heterogamety and subsequent turnover events in the genus Salix
Heredity (2023)
-
Sex effect on growth performance and marker-aided sex discrimination of seedlings of Populus deltoides
Journal of Forestry Research (2023)
-
Repeated turnovers keep sex chromosomes young in willows
Genome Biology (2022)
-
Genome-wide association mapping uncovers sex-associated copy number variation markers and female hemizygous regions on the W chromosome in Salix viminalis
BMC Genomics (2021)
-
Sex determination through X–Y heterogamety in Salix nigra
Heredity (2021)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
