
Abstract
On March 11th, 2011 the Mw 9.0 2011 Tōhoku-Oki earthquake resulted in a tsunami which caused major devastation in coastal areas. Along the Japanese NE coast, tsunami waves reached maximum run-ups of 40 m and travelled kilometers inland. Whereas devastation was clearly visible on land, underwater impact is much more difficult to assess. Here, we report unexpected results obtained during a research cruise targeting the seafloor off Shimokita (NE Japan), shortly (five months) after the disaster. The geography of the studied area is characterized by smooth coastline and a gradually descending shelf slope. Although high-energy tsunami waves caused major sediment reworking in shallow-water environments, investigated shelf ecosystems were characterized by surprisingly high benthic diversity and showed no evidence of mass mortality. Conversely, just beyond the shelf break, the benthic ecosystem was dominated by a low-diversity, opportunistic fauna indicating ongoing colonization of massive sand-bed deposits.
Similar content being viewed by others
Introduction
The Mw 9.0 2011 Tōhoku-Oki earthquake offshore of Sendai (East Japan) occurred on March 11th 2011 (05:46 UTC) and is considered the most destructive natural hazard recorded in the modern history of Japan1. With the hypocenter located at an underwater depth of approximately 24 km, 70 km east off the Oshika Peninsula of Tōhoku, this earthquake generated a tsunami that struck Japanese coastlines within minutes2. Surging waters were far more deadly and destructive than the earthquake itself, with tsunami surges recorded along all Pacific coasts including Antarctica3. In northern Honshu, the coastline around Shimokita (Fig. 1) suffered tsunami waves reaching run-up heights of 10 m. Although dramatic effects of this catastrophe were clearly visible inland, environmental consequences on the seafloor have only been documented in a limited number of studies on bathyal environments deeper than 300 m off Sendai4. Considering the scarcity of studies regarding modern tsunami impacts on shallow-water benthic marine environments5,6,7, offshore imprints related to tsunami run-up and backwash are complex to decipher in terms of both sedimentary disturbance and biodiversity response. Especially as the sedimentary and biological features observed at the seafloor are already the product of many interacting factors. Still, expected consequences of a tsunami consist of major sediment reworking at the seafloor, erosion and mass-deposition, probably resulting in mass mortality of the benthos6,7,8. Beside the inherent complexity of ecological responses to tsunami deposits, these effects are difficult to assess in the field since post-tsunami observations of shallow-water benthic ecosystems may be distorted within week or months by additional extreme meteorological events, potentially generating strong hydro-sedimentary processes5. Furthermore, paleo-tsunami records offshore are inherently difficult to interpret because ongoing short-term and long-lasting processes (e.g. bioturbation, winnowing, liquefaction, diagenetic alteration) are recorded in sedimentary archives9. Five months after the 2011 Tōhoku-Oki tsunami, we anticipated recovering sediments devastated by a catastophic event.

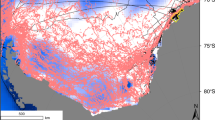
Study area off Shimokita (NE Japan).
The location of the 2011 Tōhoku earthquake epicenter is pictured in the upper map by a violet star. Both middle and lower figures show the local area and bathymetry of the four stations relative to the shoreline. In the middle figure, colored dots represent the tsunami wave height above marine sea level along the Shimokita coast47. Maps are drawn by The Generic Mapping Tools (GMT) at JAMSTEC with bathymetry data from NOAA/NGDC and Japan Oceanographic Data Center (JODC). The topographic profile are drawn by KaleidaGraph with bathymetry data recorded during our cruise.
The coast of the Shimokita Peninsula is characterized by smooth beaches and a gently sloping shelf. This setting contrasts sharply with the coastline most severely damaged by the 2011 Tōhoku-Oki tsunami at Sanriku, which possesses steep rocky shores and a sawtooth-cliff coast. For evaluating impacts on benthic ecosystems, the coast of Shimokita, with its sandy to muddy substrates, provides a more valuable reference area. The material flows generated by tsunami and their impact on benthic ecosystems are controlled by a number of factors including tsunami energy, distance from the shore, coastal bathymetry, prevailing currents and the resilience of existing ecosystems. Our interpretations are based on a multidisciplinary approach involving collection of sedimentary, geochemical and faunal data, which are supported by numerical simulations of tsunami propagation and compared with historical observations.
Results
Observations five months after the 2011 Tōhoku-Oki earthquake and tsunami
Data and samples were collected off Shimokita (NE Japan) in August 2011, five months after the 2011 Tōhoku-Oki tsunami and three months after the category 5 Typhoon Songda (Fig. 1). The study area is situated ca. 300 km north of the earthquake's epicenter and approximately 400 km from the path of Typhoon Songda10. Sediment cores from four different stations, located between 55 and 211 m depth (40°41′–40°50′N, 141°31′–141°45′E), have been studied.
Station 1 (depth 55 m) is characterized by fine to medium sands (D10 > 100 μm; D90 > 200 μm). Comparing this with historical sediment mapping11 shows no major changes in terms of sedimentary facies. However, a close inspection of our data (Fig. 2) reveals that Station 1 possesses a 4.5 cm thick mixed layer (210Pb profile) where terrigeneous particles (e.g., quartz, feldspar, rocky fragments) and volcanic material (e.g., pumice and glass) create variable grain sizes from fine to coarse sediment (290 μm < D90 < 550 μm). The sedimentary record recovered from Station 2 (depth 81 m) is topped by a 4 cm thick, coarsening-up layer composed of very poorly sorted, very coarse sand, with large (0.5–2 cm) shelly fragments. Station 3 (depth 105 m) is characterized by a 3 cm thick uppermost layer of medium to coarse sands (after decalcification; D90 > 300 μm with values around 600 μm in some intervals) (Fig. 2). A comparison with historical data off Shimokita11 shows that modern surface sediments from both stations are composed of larger particles than the fine-to-medium-sandy facies described in the 1970s. The coarse sediment layer overlaying finer sediments is stratigraphically consistent with heterogeneous layers characterized by mixed 210Pb profiles (Fig. 2). Clearly, a substantial hydro-sedimentary process supplied large sedimentary particles at both Stations 2 and 3. The sediment properties recorded at Station 4 (depth 211 m) located just beyond the shelf-break, are very different. Whilst also composed of fine to medium sands (D10 > 100 μm; D90 > 200 μm), the total 210Pb profile is indicative of a mixed layer within the uppermost 10 cm. Sandy particles from the upper 5 cm of this mixed layer are coarser (D90 maxima of 470 μm) than sediments below (250 μm < D90 < 380 μm).

Sediment properties along the bathymetric transect.
Grain-size features (in μm) are expressed in D10 and D90 for all stations. Shaded zones are related to mixed sedimentary layers, determined from radionuclide profiles. Total 210Pb activity (expressed in Bq/g) is plotted. A video captured image of the seafloor at Station 2 (81 m depth) is inserted at the bottom. Shells belonging to shallow-water bivalves and gastropods (i.e. Amusium japonicum, Arca boureardi, Olivella fulgurata) are relatively abundant. Note also the spatial heterogeneity as clearly highlighted by the chaotic distribution of the numerous shell fragments and the coarsening upwards profile overlying finer sediments. Scale = external diameter of Plexiglas tube is ~8.5 cm.
Numerical simulation of the 2011 Tōhoku-Oki tsunami's velocity (see supplementary material) indicated that bottom-water velocities reached 78 cm/s at the outer shelf at Station 3 (105 m) (Fig. 3). In contrast at Shimokita, modelling of Typhoon Songda indicated that its recorded 11 m waves at 80 m water depth generated velocities of ~17 cm/s. The local Tsugaru Warm Current generates bottom water velocities of 0.05–26 cm/s12. Velocities of 80 cm/s were required to entrain the recovered coarse, bioclastic gravels (~2 mm) of quartz density.

Simulated maximum velocities and directions of incoming and returning flows at the four stations during the 2011 Tōhoku-Oki tsunami.
The relationship between grain size (in μm) and maximum tsunami wave velocity (cm/s) is pictured on the lower figure.
Shelly fragments observed at Station 2 are notably made of inarticulate, allochthonous bivalves from the inner shelf (i.e. Amusium japonicum, Arca boureardi, Olivella fulgurata) and autochthonous bivalves and gastropods (Fig. 2). Small pieces of organic tissue were found on recovered allochthonous bivalves and occasional burrows were obseved at Stations 1, 3 and 4. These burrows cut through the coarse surface sediment layer (Fig. 2) and therefore must have been created within five months after its deposition.
Analyses of C/N ratios and organic carbon isotopes revealed that sediments from all study locations contained substantial concentrations of organic matter with varying contributions of marine compounds. Sedimentary organic matter typically has C/N ratios atomic ratios ranging between 8.4 ± 0.9 and 8.7 ± 0.4 and δ13C values ranging between −23.5‰ and −22.3‰. Both of these parameters increase from Station 1 to Station 4 (Fig. S2). Land-derived particulate organic carbon (C3 land plants and lacustrine algea) usually presents a δ13C signature close to −27‰13,14,15 versus −19 to −20‰ (for fresh marine phytodetritus) and lower C/N ratios (5.5 to 7.5)16,17. Our sample values are common in coastal sediments caused by mixing of terrestrial matter and oceanic compounds17. Furthermore, these signatures are indicative of marine organic compounds in an advanced state of degradation and indicate an absence of fresh terrestrial biomarkers in surface sediements. Visual observations of all >150 μm sieve residues dedicated to foraminiferal investigation contained no continental macrophytic remains at any of the stations investigated. The freshness of chlorophyllic pigments ([Chl a/(Chl a + Pheo a) ratio] ranging between 6.3 ± 0.8% and 8.5 ± 1.9%) and nutritional values of bulk organics (EHAA/THAA ratio between 8.3% and 15.9%) showed fairly low values with only minor spatial variability along the overall bathymetric transect (Fig. S2). The shelf-break station (Station 4) is characterized by a deposit of more sorted sand that is fairly different from other stations where chlorophyllic pigments and bulk organics values are similar.
Living (rose bengal stained) and dead benthic foraminifera (Rhizaria) were investigated as they are relevant and reliable bio-indicators of marine benthic biodiversity and resilience18,19. Compared to the three other shelf stations where dead foraminiferal tests are relatively abundant in surficial sediments (1,644 Ind./50 cc, 14,300 Ind./50 cc and 16,700 Ind./50 cc in the first top half cm of Stations 1, 2 and 3 respectively), the content of dead foraminiferal tests at Station 4 is much lower (360 Ind./50 cc in the first half cm). Therefore, sediments characterized by a terrigeneous/volcanic matrix contain few dead foraminifera. The CT-scan images (Fig. S1) support the presence of a chaotically structured and unconsolidated fabric deposited at Station 4, different from that found at the other stations.
Living (stained) foraminifera were recorded at the three shelf stations (1, 2 and 3) (49–63 taxa, H′ ranging between 2.38 and 3.31) with relatively high diversity (Fig. 4). Most observed species (Table S1) are typical from shelf environments off Japan20,21. Living communities contained a few coastal taxa (i.e. Hanzawaia nipponica; living depth < 10 m) and many epilithic species that live attached to the surface of rock fragments and volcanic minerals22 (Fig. 5). Pioneer taxa were not observed at Stations 1–3, which precludes the occurence of on-going recolonization at the time of sample collection, i.e. five months after the tsunami. Epilithic species were buried several centimeters below the sediment-water interface. As depicted in Figures 2 and 4, the vertical distribution of epilithic foraminifera broadly corresponds with the mixed sedimentary layers. Station 4 (~200 m depth), located at the shelf-break where the sea-floor slope steepens, revealed living foraminiferal fauna dominated by opportunistic single-chambered agglutinated foraminifera (86% Psammosphaera spp.), which are documented to colonize sediments after major physical disturbance (e.g., turbidite, large storm) causing benthic reworking23,24. This group likely proliferated in surface sands following the tsunami, becoming the highest standing stock (4,500 ind./100 cm2) of the bathymetric transect, five months after the catastrophic event. This exceptionally high density is not related to the bioavailability of organic compounds (i.e. EHAA/THAA ratio), which is the lowest at this station. Moreover, diversity is strongly altered at this station with only 21 taxa (H′ = 0.64) compared to the shelf stations (H′ = 2.38–3.1). The other species which are present at this station are not restricted to shelf environments but were also documented along the upper slope20,21. Inspection of dead foraminiferal fauna in the uppermost sediments revealed a large proportion of allochthonous, broken tests, such as Buccella makiyamae.

Living foraminiferal faunas including standing stock (number of individuals/100 cm2), sample diversity (number of taxa) and community composition.

Conceptual scheme illustrating the benthic marine environments off Shimokita (NE Japan) before, during and after the 2011 Tōhoku-Oki tsunami.
Loci of water particle movement by the March 2011 Tōhoku-Oki tsunami and the May 2011 Typhoon Songda is indicated.
Discussion
Here, we have applied a multidisciplinary approach to investigate the impact of tsunami on benthic ecosystems. Our four-station transect revealed several sedimentary signatures indicating relatively recent disturbance. This is summarized in a conceptual scheme illustrated in Figure 5. These signatures include coarse, heterogeneous sediments with large shelly fragments overlying finer-grained beds, a coarsening upward sequence and 210Pb profiles indicating recent sediment mixing correlating with the identified disturbed beds (Fig. S1). Considering the timing of our cruise, only two substantial events (2011 Tōhoku-Oki tsunami or Typhoon Songda) qualify as likely to cause the signatures observed. Simulated modelling of the regular Tsugaru Warm Current, the 2011 Tōhoku-Oki tsunami and Typhoon Songda velocities indicated that bottom-water speeds of investigated sediments reached up to 80 cm/s at depths lower than 100 m. This is beyond the estimated velocity of Typhoon Songda and therefore only the 2011 Tōhoku-Oki tsunami generated sufficient force to entrain the recovered sediments. This conclusion is in line with other collected environmental data. Therefore, the sediments studied here are regarded as tsunamigenic deposits not modified by the subsequent passage of Typhoon Songda.
Offshore tsunami deposits of both the 2011 Tōhoku-Oki tsunami and earlier events illustrate their variety in size, nature and permanence7,25,26. Here, these deposits are most clearly indicated at Stations 1–3 by coarse beds overlying fine-grained beds. However, Station 2 and potentially Station 3 additionally display coarsening upward sequences (Fig. 2). These sequences are likely related to tsunami backwash carrying coarse material offshore from shallow-water settings. Uncertainty remains at Station 3 as to whether a coarsening upward sequence is present (Fig. 2), which is obscured by subsequent bioturbation (Fig. S1). Stations 1–3 are represented by heterogeneous, biogenic, terrigenous and volcanic sediments distributed along the shelf. At the shelf-break, Station 4 is fairly different, characterized by a deposit of more sorted sand. Tsunami backwash has great capacity to shift substantial amounts of onshore sediments out to sea26,27,28. The coarse, poorly sorted, offshore sedimentation found in this study may have occurred by retreating waves, entraining material to form dense sediment flows nearshow which then flowed out to sea27.
Five months after the disaster, the three shelf stations (Stations 1–3) are characterized by tsunamigenic deposits made of freshly reworked sediments consisting of poorly sorted sands and relatively degraded marine organic material. Marine organic material was likely redistributed uniformly over both the shelf and the upper-slope areas by tsunami waves. Because of the relatively uniform distribution of sediments by the tsunami across Stations 1–3, there are only slight variances in organic matter concentration.
Living foraminiferal communities from open marine environments are constrained by many physicochemical parameters amongst which sedimentary organic compounds, oxygenation level and hydrosedimentary processes constitute major ecological influences. Therefore, their diversity is used for bio-monitoring in marine environments impacted by anthropogenic disturbances and also for paleo-environmental reconstruction including tsunamis18,29,30. For instance, opportunistic taxa are able to colonize freshly deposited sediment layers during massive sedimentation events. Low-diversity colonizing assemblages have been recorded in shelf environments influenced by riverine flooding or in deep-sea ecosystems impacted by turbidites24,31,32. The four investigated stations show two alternative types of recovery following a catastrophic event. At Stations 1–3, original sediments were mixed and some components transported further down shelf towards the shelf break (Station 4 region). The original shelf setting of Stations 1–3 was characterized by a relatively low energy and dominance of fine-grained sediments. Post-tsunami, the now poorly sorted, coarser sediments, rich with organic material, support a flourishing benthic community with an unexpectedly high diversity. Foraminiferal abundance in both live and dead specimens were reported to increase over a similar depth transect, only three months after the 2003 Tokachi-oki earthquake and tsunami off Hokkaido5. Unlike here, change was not uniform across all sites as many post-tsunami sediments also recorded reductions in living and dead foraminiferal abundances. What is particularly noteworthy in our study is the contrasting abundance and distribution of epilithic species. Original burrows of these taxa (1–2 cm beneath sediment water interface) would have been likely destroyed by the tsunami, but the freshly deposited surficial sediments has resulted in them inhabiting deeper sediments (3–5 cm) where they continued to survive (Fig. 4). It is possible that the now bioturbated deposit is sufficiently oxygenated to sustain them at a deeper horizon. Vertical displacement to deeper sediments due to sediment disturbance in live benthic communities has been previously reported for meiofauna33,34,35 but not foraminifera.
The alternative recovery pathway was detected at the shelf-break (~200 m depth, Station 4) where the original foraminiferal assemblage was decimated by the impact of the tsunami, substantially decreasing diversity. This allowed colonization by the opportunistic, agglutinated taxa Psammosphaera spp., to establish a new assemblage with high dominance and low diversity. This microbenthic response was likely triggered by the tsunamigenic event causing distal, massive sand-bed deposition. Rapid recolonization of turbidite sediments after a tsunami event were similarly found in meiofaunal communities after the 2004 Indian Ocean tsunami, however it was not by single genus and this behaviour varied between different taxa34. The allochthonous dead foraminifera in the terrigenous/volcanic matrix at Station 4 further eludes to mass deposition of shallower sediments offshore commonly reported in similar studies5,27. Here, this is due to the presence of commonly shallow water taxa such as B. makiyamae which is not normally found in such high abundance at these depths. Furthermore, as found with coiled, moderately inflated morphologies, the flying-saucer shape of B. makiyamae facilitates selective transportation and sorting along the shelf and to the upper slope when backwash turbidity currents ran downslope over the shelf break36,37,38. The use of foraminifera in offshore tsunami studies includes isolating tsunami deposits25,27, gauging tsunami outflow strength and direction5,27 and monitoring the return to the original environmental setting for a region39. Tsunami studies using foraminifera rarely have the opportunity to investigate live vs. dead fauna nor note the recovery of the disturbed benthic communities18. This is understandable considering the inherent difficulties of such investigations in both paleo- (difficult to establish recovery timeline) and modern (difficult to return to deposit site, little faunal evidence of disturbance) tsunami investigations40,41,42. Furthermore, tsunami deposits are more commonly investigated on land or in sheltered shallow marine, brackish or marine proximal fresh water settings28,43. Yet here we present unusual insight into both the nature of tsunami deposits in deeper marine settings and the varying response of foraminiferal communities to tsunamigenic stress.
Studies regarding the composition of benthic communities post-tsunami are rare and report varying reactions over different periods of time27,39. The recovery of offshore foraminiferal assemblages to return to their original character can take one to two years27. Similar to Station 4, an entire new benthic assemblage can be established before the original returns39. Whilst such dramatic environmental changes did not occur off Shimokita, it is clear that the recovery/returning of benthic assemblages to their pre-tsunami character varies substantially in both timing and nature across different environmental settings. Caution must be employed with assessing post-tsunami assemblages. Live/dead investigations are encouraged to ensure that if assemblage diversity changes after an event, it is a product of recovery and not deposited tsunami sediments containing a different assemblage from another region44,45.
This study highlights that the response and resilience of benthic communities to tsunami in offshore settings varies due to a number of parameters. Living foraminiferal faunas have been found to both rapidly recover and flourish after a tsunami event or for a new community to replace the original. Future sampling in the investigated area could reveal long-term recovery from the tsunami where abundance and diversity may return to their pre- March 2011 levels. Work presented here expands the understanding of tsunami impact offshore and the varied reaction of foraminiferal communities in different marine settings.
Methods
The oceanographic cruise KT11-20 aboard the R/V Tansei Maru (AORI, University of Tokyo/JAMSTEC) took place off Shimokita (NE Japan) in August 2011, five months after the Tōhoku-Oki disaster and three months after Typhoon Songda (May, 2011). The study area is situated ca. 300 km north of the earthquake's epicenter. At this location, the marine shelf is swept from north to south by the Tsugaru Warm Current transporting warm and saline water from the Japan Sea into the Pacific Ocean through the Tsugaru Straight46. Four stations located between 55 and 211 m depth (40°41′–40°50′N, 141°31′–141°45′E) were sampled with a classical Barnett multi-corer designed to collect eight sediment cores per deployment (Fig. 1). At each investigated station, one sediment core (surface area = 53 cm2) was dedicated to organic matter analyses (C/N ratio atomic ratio, [EHAA/THAA] ratio, [Chl a/(Chl a + Pheo a)] ratio, organic matter δ13C, biomarkers) and to grain-size distribution (e.g., D10 and D90). A second core was used to perform 210Pb measurements. Sedimentological descriptions (lithological and mineralogical analyses and CT-scan) were performed on a third core. A final core was dedicated to quantify living (rose bengal stained) benthic foraminifera as they are relevant and reliable bio-indicators of marine benthic biodiversity and resilience18. All investigated cores were sliced horizontally in varying sediment intervals (see supplementary material online). A numerical simulation of the 2011 Tōhoku-Oki tsunami propagation effect on both water mass (e.g. bottom-current velocity) and sediments was performed to strengthen observation and interpretation. Further details regarding sampling and methods are provided in the online supplementary information with living foraminiferal census data. CT-scan pictures are presented in online Supplementary Table S1 and Figure S1.
References
Koketsu, K. et al. A unified source model for the 2011 Tohoku earthquake. Earth Planet. Sc. Lett. 310, 480–487 (2011).
Tsuji, Y. et al. Field surveys of tsunami heights from the 2011 off the Pacific Coast of Tohoku, Japan Earthquake. B. Earthq. Res. I. Tokyo 86, 29–279 (2011).
Brunt, K. M., Okal, E. A. & MacAyeal, D. R. Antarctic ice-shelf calving triggered by the Honshu (Japan) earthquake and tsunami, March 2011. J. Glaciol. 57, 785–788 (2011).
Arai, K. et al. Tsunami-generated turbidity current of the 2011 Tohoku-Oki earthquake. Geology 41, 1195–1198 (2013).
Noda, A. et al. Evaluation of tsunami impacts on shallow marine sediments: An example from the tsunami caused by the 2003 Tokachi-oki earthquake, northern Japan. Sediment. Geol. 200, 314–327 (2007).
Lomovaski, B. J., Firstater, F. N., Gamarra Salazar, A., Mendo, J. & Iribarne, O. O. Macro benthic community assemblage before and after the 2007 tsunami and earthquake at Paracas Bay, Peru. J. Sea Res. 65, 205–212 (2011).
Sakuna, D., Szczucinski, W., Feldens, P., Schwarzer, K. & Khokiattiwong, S. Sedimentary deposits left by the 2004 Indian Ocean tsunami on the inner continental shelf offshore of Khao Lak, Andaman Sea (Thailand). Earth Planets Space 64, 931–943 (2012).
Seike, K., Shirai, K. & Kogure, Y. Disturbance of Shallow Marine Soft-Bottom Environments and Megabenthos Assemblages by a Huge Tsunami Induced by the 2011 M9.0 Tohoku-Oki Earthquake. PLoS ONE 8, e65417, 10.1371/journal.pone.0065417 (2013).
Chague-Goff, C. Chemical signatures of palaeotsunamis: A forgotten proxy? Mar. Geol. 271, 67–71 (2010).
Kitamoto, A. Typhoon 201102 (SONGDA). (2011) Available at: http://agora.ex.nii.ac.jp/digital-typhoon/summary/wnp/s/201102.html.en. (Accessed: 7th May 2014).
Arita, M. & Kinoshita, Y. in Marine Geology Map series no. 9 (Geological Survey of Japan, Tsukuba, 1978).
Japan Oceanographic Data Centre. Oceanographic Data and Information: Ocean Current Data. (2014) Available at: http://www.jodc.go.jp/. (Accessed: 5th September 2014).
Meyers, P. A. Preservation of elemental and isotopic source identification of sedimentary organic matter. Chem. Geol. 114, 289–302 (1994).
Ogawa, N. & Ogura, N. Dynamics of particulate organic matter in the Tamagawa estuary in Inner Tokyo Basin. Estuar. Coast. Shelf Sci. 44, 263–273 (1997).
Barth, J. A. C., Veizer, J. & Mayer, B. Origin of particulate organic carbon in the upper St. Lawrence: isotopic constraints. Earth Planet. Sc. Lett. 162, 111–121, 10.1016/S0012-821X(98)00160-5 (1998).
Kitazato, H. et al. Seasonal phytodetritus deposition and responses of bathyal benthic foraminiferal populations in Sagami Bay, Japan:Preliminary results from “Project Sagami”. Mar. Micropaleontol. 40, 135–149 (2000).
Nakatsuka, T., Masuzawa, T., Kanda, J., Kitazato, H. & Shirayama, Y. Particle dynamics in the deep water column of Sagami Bay, Japan. I: origins of apparent flux of sinking particles. Prog. Oceanogr. 57, 31–45, 10.1016/S0079-6611(03)00049-1 (2003).
Mamo, B., Strotz, L. & Dominey-Howes, D. Tsunami sediments and their foraminiferal assemblages. Earth-Sci. Rev. 96, 263–278 (2009).
Murray, J. W. Ecology and Applications of Benthic Foraminifera. (Cambridge University Press, 2006).
Ishiwada, Y. Benthonic foraminifera off the Pacific coast of Japan referred to biostratigraphy of the Kazusa group. Geol. Sur. Japan 205, 1–45 (1964).
Ikeya, N. in Reports of Faculty of Sciences Vol. 6 179–201 (Shizuoka University, 1971).
Matoba, Y. Recent foraminiferal assemblages off Sendai, northeast Japan. Mar. Sedimentol. Spec. Publ. 1, 205–220 (1976).
Kaminski, M. A. Evidence for control of abyssal agglutinated foraminiferal community structure by substrate disturbance. Mar. Geol. 66, 113–131 (1985).
Hess, S., Jorissen, F. J., Venet, V. & Abu-Zied, R. Benthic foraminiferal recovery after recent turbidite deposition in Cap Breton Canyon (Bay of Biscay). J. Foraminifer. Res. 35, 114–129 (2005).
Cita, M. B. & Aloisi, G. Deep-sea tsunami deposits triggered by the explosion of Santorini (3500 y BP), eastern Mediterranean. Sediment. Geol. 135, 181–203 (2000).
Goto, K., Ikehara, K., Goff, J., Chagué-Goff, C. & Jaffe, B. The 2011 Tohoku-oki tsunami — Three years on. Mar. Geol. in press, 10.1016/j.margeo.2014.08.008 (2014).
Sugawara, D. et al. Foraminiferal evidence of submarine sediment transport and deposition by backwash during the 2004 Indian Ocean tsunami. Isl. Arc 18, 513–525 (2009).
Minoura, K. & Nakaya, S. Traces of tsunami preserved in inter-tidal lacustrine and marsh deposits; some examples from Northeast Japan. J. Geol. 99, 265–287 (1991).
Jorissen, F. J., Fontanier, C. & Thomas, E. in Paleoceanography of the Late Cenozoic Vol. 1: Methods in late Cenozoic Paleoceanography (eds Hillaire-Marcel, C. & de Vernal, A.) 875 (Elsevier, 2007).
Schönfeld, J. et al. The FOBIMO (FOraminiferal BIo-MOnitoring) initiative—Towards a standardised protocol for soft-bottom benthic foraminiferal monitoring studies. Mar. Micropaleontol. 94–95, 1–13 (2012).
Goineau, A. et al. Temporal variability of live (stained) benthic foraminiferal faunas in a river-dominated shelf – faunal response to rapid changes of the river influence (Rhône prodelta, NW Mediterranean). Biogeosci. Disc. 8, 9033–9086 (2012).
Fontanier, C. et al. Deep-sea foraminifera from the Cassidaigne Canyon (NW Mediterranean): Assessing the environmental impact of bauxite red mud disposal. Mar. Pollut. Bull. 64, 1895–1910 (2012).
Ingole, B. S., Ansari, Z. A., Rathod, V. & Rodrigues, N. Response of deep-sea macrobenthos to a small-scale environmental disturbance. Deep-Sea Res. Pt II 48, 3401–3410, 10.1016/S0967-0645(01)00048-0 (2001).
Altaff, K., Sugumaran, J. & Naveed, S. Impact of tsunami on meiofauna of Marina beach, Chennai, India. Curr. Sci. 89, 34–38 (2005).
Kitahashi, T. et al. Effect of the 2011 Tohoku Earthquake on deep-sea meiofaunal assemblages inhabiting the landward slope of the Japan Trench. Mar. Geol. in press, 10.1016/j.margeo.2014.05.004 (2014).
Duros, P. et al. Live and dead benthic foraminiferal faunas from Whittard Canyon (NE Atlantic): Focus on taphonomic processes and paleo-environmental applications. Mar. Micropaleontol. 94–95, 25–44, 10.1016/j.marmicro.2012.05.004 (2012).
Snyder, S. W., Hale, W. R. & Kontrovitz, M. Assessment of Postmortem Transportation of Modern Benthic Foraminifera of the Washington Continental Shelf. Micropaleontology 36, 259–282, 10.2307/1485509 (1990).
Duros, P. et al. Study of benthic foraminiferal thanatocoenoses from the Cap-Ferret Canyon area (NE Atlantic): A complex interplay between hydro-sedimentary and biological processes. Deep-Sea Res. Pt II (in press).
Clague, J. J., Hutchinson, I., Mathewes, R. W. & Patterson, R. T. Evidence for Late Holocene Tsunamis at Catal Lake, British Columbia. J. Coast. Res. 15, 45–60 (1999).
De Martini, P. M. et al. A unique 4000 year long geological record of multiple tsunami inundations in the Augusta Bay (eastern Sicily, Italy). Mar. Geol. 276, 42–57 (2010).
Dawson, A. Linking tsunami deposits, submarine slides and offshore earthquakes. Quatern. Int. 60, 119–126 (1999).
Sun, L. et al. Preliminary evidence for a 1000-year-old tsunami in the South China Sea. Sci. Rep. 3, 1–5 (2013).
Scheffers, A. & Kelletat, D. Sedimentologic and geomorphologic tsunami imprints worldwide—a review. Earth-Sci. Rev. 63, 83–92, 10.1016/S0012-8252(03)00018-7 (2003).
Gadi, S. D. & Rajashekhar, K. P. Changes in inter-tidal foraminifera following tsunami inundation of Indian coast. Indian J. Mar. Sci. 36, 35–42 (2007).
Mukherjee, N. et al. From Bathymetry to Bioshields: A Review of Post-Tsunami Ecological Research in India and its Implications for Policy. Environ. Manage. 46, 329–339, 10.1007/s00267-010-9523-1 (2010).
Ito, T. et al. Variation of velocity and volume transport of the Tsugaru Warm Current in the winter of 1999–2000. Geophys. Res. Lett. 30, 1678, 10.1029/2003gl017522 (2003).
Mori, N., Takahashi, T. & Group, T. T. E. T. J. S. Nationwide post event survey and analysis of the 2011 Tohoku earthquake and tsunami. Coast. Eng. J. 54, 1–27, 10.1142/S0578563412500015 (2012).
Acknowledgements
We thank crew members of R/V Tansei-Maru, which was supported by the Atmosphere and Ocean Research Institute, The University of Tokyo/JAMSTEC. T.T., K.O. and H.K. were supported by grants-in-aid for scientific research by MEXT/JSPS (20740301 to T.T., 23510022 to K.O. and 21244079, 25247085 to H.K., T.T. and K.O.). K.A.K. would like to acknowledge NWO-ALW (Dutch Earth and Life sciences council) for funding of her research (grant numbers 820.01.011). Financial support was provided to A.G. and B.D. by the French national programs ‘EC2CO’ and ‘LEFE-CYBER’ (BIOMIN project). C.F. thanks the Centre National de la Recherche Scientifique for their support of his 1.5 years of delegation at the EPOC laboratory.
Author information
Authors and Affiliations
Contributions
T.T., who was in charge of the ocean cruise KT11-20, coordinated Japanese research activities after the cruise and finalized the present manuscript. P.D. worked on foraminiferal faunas from Stations 1, 2 and 3 and generated both Figures 2 and 4 in the draft. C.F. participated in the cruise and supervised the investigation of foraminiferal faunas from the shelf-break Station 4. C.F. contributed intensively to the redaction of the draft and generated Figure 5. B.M. reviewed literature with which to compare our findings, revised the English of the draft and provided constructive discussions to improve the understanding of our story. S.B. and C.M. performed analyses of organic descriptors. A.G., B.D. and R.B. supervised these analyses and provided reliable correction on the draft. G.C. was in charge of grain-size analyses. S.G. completed the practical work on foraminifera from Station 4. M.F., A.S. and K.K. described sedimentological facis. A.N. and Y.N. constructed tsunami numerical models for Figure 3 and 5. K.O. and O.R. did radionuclide measurements on sediment cores and made a lot of useful comments on the draft. K.O. participated on the cruise. M.M. scaned all cores by CT. K.A.K., L.J.N. and G.J.R. participated susbstantially during cruise sampling, provided complementary geochemical observations and improved the draft and the revision. A.K. and N.O. processed on-board oceanographic data and compiled inland run-up height of tsunami to generate Figures 1 and 5. T.O. identified the bivalves to determine their original habitable depth. H.K., F.J., G.J.R. and C.F. organize the international collaborative project and planning of the cruise proposal.
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Electronic supplementary material
Supplementary Information
Supplementary information
Rights and permissions
This work is licensed under a Creative Commons Attribution-NonCommercial-ShareAlike 4.0 International License. The images or other third party material in this article are included in the article's Creative Commons license, unless indicated otherwise in the credit line; if the material is not included under the Creative Commons license, users will need to obtain permission from the license holder in order to reproduce the material. To view a copy of this license, visit http://creativecommons.org/licenses/by-nc-sa/4.0/
About this article

Cite this article
Toyofuku, T., Duros, P., Fontanier, C. et al. Unexpected biotic resilience on the Japanese seafloor caused by the 2011 Tōhoku-Oki tsunami. Sci Rep 4, 7517 (2014). https://doi.org/10.1038/srep07517
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/srep07517
This article is cited by
-
Geochemical approaches in tsunami research: current knowledge and challenges
Geoscience Letters (2021)
-
Repeated occurrence of surface-sediment remobilization along the landward slope of the Japan Trench by great earthquakes
Earth, Planets and Space (2020)
-
Impact of the huge 2011 Tohoku-oki tsunami on the phenotypes and genotypes of Japanese coastal threespine stickleback populations
Scientific Reports (2018)
-
Dissolved Platinum Concentrations in Coastal Seawater: Boso to Sanriku Areas, Japan
Archives of Environmental Contamination and Toxicology (2017)
-
Seawater influence monitored by NaCl on the growth of Trametes versicolor
Environmental Science and Pollution Research (2016)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
