
Abstract
Different types of sediments in salt marsh have different physical and chemical characters. Thus sediment type plays a role in plant competition and growth in salt marsh ecosystems. Spartina anglica populations have been increasingly confined to upper elevation gradients of clay and the niche sediment has changed. Because the niches of S. anglica and the native species Scirpus triqueter overlap, we conducted a greenhouse experiment to test the hypothesis that plant competition has changed under different types of sediments. Biomass and asexual reproduction were analyzed and inter- and intraspecific competition was measured by log response ratio for the two species in both monoculture and combination under three sediment types (sand, clay and mixture of sand and clay). For S. anglica, biomass, ramet number and rhizome length in combination declined significantly compared with those in monoculture and the intensity of interspecific competition was significantly higher than that of intraspecific competition under all sediments. For S. triqueter, the intensities of intra- and interspecific competition were not significantly different. This indicates that S. triqueter exerts an asymmetric competitive advantage over S. anglica across all sediments, but especially clay. Thus the sediment type changes competition between S. anglica and S. triqueter.
Similar content being viewed by others
Introduction
Zonal distribution of plant communities is an obvious characteristic of salt marsh ecosystems1,2,3,4. The plant zonation pattern is typically correlated with a combination of multiple factors, such as the saturated or unsaturated flow in the soil2 and sediment accretion5,6. In fact, plant community spatial distribution and abundance of species are both an indication of the relative physiological tolerance of an individual species to abiotic conditions and competition among species1,4,7,8,9.
Plant competition can be affected not only by resource availability10, but also by environmental factors within a salt marsh4,11,12,13, such as sediment9,14,15, sea level4,5 and salinity7,16. It has been shown that competition between species, especially between exotic and native species, often depend on nutrient supply9,17,18, flooding19, elevation4 and other factors20. Elevation, which varies with plant zonation, is highly correlated with the redox potential in salt marsh sediment21. Also, interspecific competition has also identified as always disproportionate, resulting in the replacement of one species by another16. However, there is typically little information available about competition between exotic and native species in different sediments types.
Salt marsh sediments are mainly influenced by plant activity, elevation and tidal flooding14,22,23,24,25. Because much of the organic matter in tidal flooding originates from external sources or from benthic microalgae, sediments are usually rich in organic matter and phosphorus26,27,28. Also, sediments of salt marshes are characterized with high concentrations of sulfides and ammonium originating from the high rates of anaerobic reduction5,23, which can affect plant root respiration29. Research has shown that poor water drainage of marsh sediments can be a critical factor limiting plant root growth and causing decay23. In addition, moderate levels of sediment slurry enrichment can have beneficial effects on the soil by increasing elevation and soil bulk density30, which could assist with the restoration of the macro-invertebrate community and its related habitat31. Thus supplementary sediments from tides ameliorate environmental factors for plant growth for some species27, including Spartina anglica32,33.
Spartina anglica C. E. Hubbard (hereafter called Spartina) is an exotic species originating in England that was first introduced in China in 196334,35. It developed populations successfully for 30 years on Chinese salt marsh36 and provided significant economic benefits, including protection for dams and feed for livestock. Yet, in recent years, Spartina populations have been in decline on the Chinese coast and the cover has decreased to less than 50 ha34,36. Spartina has been pushed to a higher elevation gradient due to the recovery of the salt marsh and the expansion of Spartina alterniflora, which is another exotic and invasive species36. Also there are differences in physical and chemical characters between sediments at low and high tide zones.
Scirpus triqueter L. (hereafter called Scirpus) is species native to China37. It grows on low-lying moist sites, where an overlapping niche exists with Spartina and it likely results in competition between the two plant species. Although research has been conducted regarding the ecological interaction between the two plant species with different nitrogen levels38, little is known of competition between the two plant species under different types of sediment.
In this experiment we used three kinds of sediment, i.e. clay, sand and a mixture of those two sediments, to test competition outcomes between the native species, Scirpus and the exotic species, Spartina. As sediment structure is known to vary within saltmarshes, we investigated how such variation might influence competitive outcomes between Spartina and Scirpus. Here, we quantified competition using various growth measurements when plants were grown with and without their competitor present.
Results
Competition intensity
The more negative the values of the log response ratio (LogRR) were, the greater was competition intensity. Sediment type significantly affected competition intensity, as measured by LogRR, in both species (Table 1). For Spartina, both inter- and intraspecific competitions were greater when it grew in clay than when it grew in sand or in the sand-clay mixture (Figure 1A). For Scirpus, competition in sand and clay did not differ significantly, but it was greater than that in the sand-clay mixture (Figure 1B).

Interspecific and intraspecific log response ratio (LogRR) of Spartina anglica (A) and Scirpus triqueter (B).
Means ± SE are presented.
The competition type significantly affected LogRR of Spartina, but not that of Scirpus (Table 1B). For Spartina, interspecific competition was significantly greater than intraspecific competition (Figure 1A). For Scirpus, intraspecific competition did not differ significantly from interspecific competition (Figure 1B).
Growth measures
Sediment type and competition between species affected significantly (P < 0.05) or tended to affect (P < 0.1) all growth measures of both Spartina and Scirpus (Table 2). The interaction between sediment type and competition type was statistically significant for all measures of growth for Spartina (except rhizome length), however the interaction term was never statistically significant for Scirpus (Table 2).
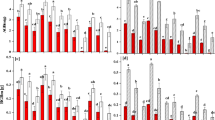
For Spartina, all growth measures were significantly larger when the sediment was clay than when it was sand or the mixture of sand and clay; they did not differ significantly between the sand and sand-clay mixture treatments (Table 2A, Figure 2, SNK tests). Overall, growth measures of Spartina were the largest in SA2, smallest in SA2 + ST2 and intermediate in SA4 (Table 2A, Figure 2). Such effects were larger when the sediment was clay than when it was sand or the sand-clay mixture (Table 2A, Figure 2).

Biomass and asexual characteristics of Spartina anglica in different sediment types and species competition experiments.
Means ± SE are presented.
For Scirpus, all growth measures except number of ramets were significantly larger when the sediment was clay than when it was sand or the sand-clay mixture (Table 2B, Figure 3, SNK tests). The number of ramets of Scirpus in clay was significantly higher than that in the sand-clay mixture, but did not differ from that in sand (Figure 3B). All growth measures except number of ramets were significantly larger in ST2 than in SA2 + ST2 and ST4, but they did not differ between SA2 + ST2 and ST4 (Table 2B, Figure 3).

Biomass and asexual characteristics of Scirpus triqueter in different sediment types and species competition experiments.
Means ± SE are presented.
Discussion
Our results suggest that sediment type has significant effects on the growth of Spartina. Although Spartina could adapt to different sediments1,24,29, it still performs better in more nutrient-rich sediments such as clay39, likely because the ability of clay to preserve moisture and nutrients is greater than sand. Scirpus had good growth performance in all sediments, perhaps because of its low nutrient needs38.
The sediment type in salt marsh depends on factors such as plant species27, sea level40, tide32, tidal creeks15 and elevation21. Tidal flooding gives salt marsh increased vigor because of the sediment component of the substrate, which increases the soil mineral matter and decreases nutrient deficiency30,32. Tidal creeks shape sequential geomorphic features, which receive different types of sediment (coarse or fine) from tides and channels15. The clay is a fine-type sediment driven by hydro-geomorphic processes with low bulk density, which yields poor drainage conditions and limits plant root growth and causing decay15,23. The clay can also change the redox conditions and sulphide concentrations, which can affect plant growth39. For instance, the health of plants shows a sharp decline when the redox potential falls below −50 mv21 and the sulphide concentration can reach about 320 mg kg−1 in the sediment water interface of Spartina alterniflora39. Simply, abiotic factors including the sediment electric conductivity, sediment oxygenation and salinity affect seed germination and subsequent plant growth41. These physical and chemical sediment traits may greatly influence the growth and adaption of plant species30,41,42, such as Spartina and Scirpus, thus changing their competition.
The intensities of interspecific and intraspecific competition for both species were significantly different with sediment types. For Spartina, interspecific competition from Scirpus was larger than intraspecific competition and competition intensity was greatest in clay (Figure 1A). On the other hand, for Scirpus, overall there was no significant difference between interspecific and intraspecific competition and intraspecific competition tended to be higher than interspecific competition in clay (Figure 1B). These results suggest that, when the two species grow together, Scirpus had a stronger competitive effect on Spartina in clay.
Competition between plant species can be changed by environmental factors4,8,11,17,43,44 and our previous study has also shown that nitrogen level could change competition between Spartina and Scirpus38. In this study, we found that competition between Spartina and Scirpus became stronger when the sediment was clay than when they were sand or a clay-sand mixture. Similarly, sediment type has been found to significantly alter competition between Puccinellia maritima and Spartina1 and between S. alterniflora and P. australis43. Changes in competition between Spartina and Scirpus under different sediment type suggest that in the upper zone salt marsh where Spartina is currently distributed and where Scirpus is abundant replacement of Spartina by Scirpus may happen. Therefore, we predict that further declines in Spartina are likely to take place in the upper zone salt marsh. Because S. alterniflora invasion drives up45,46, there are some differences in sediment types among low and high zone salt marsh47,48,49,50. However, for a more accurate prediction, effects of other environmental factors such as tide action and salinity on competition between two species should be taken into consideration. Our findings might facilitate the development of schemes to control the Spartina invasion that is occurring in some countries in the world.
Methods
The species
Spartina is a rhizomatous perennial grass that is highly invasive in German, Australian, Irish estuarine mudflats, sand flats and salt marshes35,51,52. The plant may reach a height of 50–100 cm and its leaf blades are flat or in-rolled and 5–12 mm wide. Spartina is mainly wind pollinated and produces abundant flowers and viable seeds in Europe36. However, on the coastal areas of China, Spartina rarely produces viable seeds due to its poor pollen quality and abnormal pollen tube; it spreads mainly by clonal growth36,53. The growth form of Spartina varies in different habitats35.
Scirpus is a rhizomatous perennial sedge native to China37. The plant may reach a height of about 20–100 cm. Its stem is trigonous, with leaves 1.5–5.5 cm in length and 1.5–2.5 mm in width. Scirpus flowers and produces seeds from June to September. It occurs in different habitats in tidal wetlands, ranging from brackish to freshwater along the coast37.
Experimental design
Plants of both Spartina and Scirpus were collected from the mash zone of Xinyang Harbor in Yancheng Wetland National Nature Reserve in Jiangsu Province, China. The plant collection was authorized by the management of Yancheng Wetland National Nature Reserve because the field studies did not involve endangered or protected species. Similar-sized plants (ramets), each consisting of a single tiller with attached roots, were selected and used for this experiment. All the ramets of both species were collected within about 500 m, which should reduce the chance of different genotypes. All selected ramets were about 6 cm tall and were cultivated in pots (28 cm in diameter and 20 cm in height) containing a 1:1 (v:v) mixture of sand and clay. For the experiment, we used similar-sized ramets with a height of 12.6 ± 0.5 cm (mean ± SE) for Spartina and 15.2 ± 0.5 cm for Scirpus. The ramets were selected at random from the cultivated stock population to reduce the possible influence of clonal variation and plant history.
The experiment had three types of sediment and five species combinations in a factorial design, resulting in 15 treatments. The three sediment types were (1) sand, (2) clay and (3) a mixture of sand and clay at a volume ratio of 1:1. The amounts of available nitrogen and phosphorus were measured before the experiment. The amount of available nitrogen was 43.65 ± 2.49 (mean ± SE, n = 3) mg kg−1 in sand, 76.34 ± 1.23 mg kg−1 in clay and 52.5 ± 3.59 mg kg−1 in the sand-clay mixture. The amount of available phosphorus was 1.42 ± 0.25 mg kg−1 in sand, 8.31 ± 0.76 mg kg−1 in clay and 5.09 ± 0.18 mg kg−1 in the sand-clay mixture. There were five species combination treatments, i.e. each pot was planted with (1) two ramets of Spartina (coded as SA2), (2) two ramets of Scirpus (ST2), (3) four ramets of Spartina (SA4), (4) four ramets of Scirpus (ST4) and (5) two ramets of Spartina and two ramets of Scirpus (SA2 + ST2).
We sampled the salt water in the natural habit of Spartina and Scirpus to measure the salinity, which was about 1.48 ± 0.02% (mean ± SE, n = 3). To simulate the marsh conditions that these two species commonly experienced, salt water containing 1.5% NaCl was added to the pots and water was maintained 2 cm above the soil surface level. The salt water was produced by dissolving crude salt into tap water. The crude salt had been directly extracted from seawater in Jiangsu Province. The salinity content in the water in the pots was monitored weekly and adjusted to the initial conditions (1.5% NaCl) when it was below 1.4% or above 1.6%. The experiments lasted 26 weeks (from 20 May to 4 December) and were carried out in a greenhouse at the Pukou campus of Nanjing University. There were six replicates for each treatment.
Data collection
At harvest, we measured total biomass, the number of ramets and rhizome length of each plant for the two plant species. Biomass is a typical measure of plant growth, the number of ramets a measure of asexual (clonal) reproduction and rhizome length a measure of clonal expansion (the distance of lateral clonal spreading). We counted the number of ramets and measured the rhizome length of Spartina and Scirpus separately. Plants of Spartina and Scirpus were dried at 80°C for 72 h and weighed.
No plants produced flowers or set seeds during the experiment. One plant of Spartina in SA2 + ST2 grown in the sand-clay mixture died during the experiment and this replicate was excluded from harvest and subsequent analysis for the two species.
Data analysis
Before analysis, we normalized the data from each plant. For each replicate, we calculated biomass, number of ramets and rhizome length per initial ramet. For instance, for SA2 the final biomass was divided by two and for SA4 it was divided by four. Thus all these measures were standardized to per initial plant level. The subsequent analyses were based these derived data.
We used two-way ANOVA to test the effects of sediment type and species competition on all growth measures of each species. Sediment type and species competition were treated as two fixed factors. Sediment type had three levels (sand, sand-clay mixture and clay) and species competition had also three levels (no competition, with intraspecific competition and with interspecific competition). For species competition of Sparina, SA2, SA4 and SA2 + ST2 were used, treating SA2 as no competition, SA4 as with intraspecific competition and SA2 + ST2 as with interspecific competition. Similarly, for competition of Scirpus, ST2, ST4 and SA2 + ST2 were used, treating ST2 as no competition, ST4 as with intraspecific competition and SA2 + ST2 as with interspecific competition. We used the Student-Newman-Keuls (SNK) test to compare the overall means among the three sediment treatments and among the three competition treatments.
To measure the intensity of interspecific competition, we calculated the log response ratio (LogRR) as LogRR = log (B+/B0), where B+ is biomass of plants in the presence of the interspecific neighbors and B0 is the mean biomass of plants in the absence of neighbors across the six replicates55,56. Similarly, we also measured the intensity of intraspecific competition by calculating LogRR, where B+ is biomass of plants in the presence of the intraspecific neighbors and B0 is the mean biomass of plants in the absence of neighbors across the six replicates54,55. A positive value of LogRR indicates facilitation and a negative value indicates competition54,55,56.
In this study, the absence of neighbors (i.e. no competition) meant the treatments in which one pot was planted with two ramets (either Spartina or Scirpus), the presence of intraspecific neighbors (with intraspecific competition) referred to the treatments in which one pot was planted with four ramets of the same species and the presence of interspecific neighbors (with interspecific competition) referred to the mixture treatment in which each pot was planted with two ramets of both Spartina and Scirpus. In other words, intraspecific competition encompassed competition mainly from the same species, while interspecific competition encompassed competition mainly from other species. We used two-way ANOVA to test the effects of sediment type (three levels: sand vs. sand-clay mixture vs. clay) and interaction type (two levels: intraspecific competition vs. interspecific competition) on LogRR.
Statistical analyses were conducted with SPSS 18.0 for Windows (SPSS Inc., USA). The effects were considered significant if P < 0.05. Measures of biomass were log transformed to improve homogeneity of variance prior to ANOVA.
References
Huckle, J. M., Potter, J. A. & Marrs, R. H. Influence of environmental factors on the growth and interactions between salt marsh plants: effects of salinity, sediment and waterlogging. J. Ecol. 88, 492–505 (2000).
Silvestri, S., Defina, A. & Marani, M. Tidal regime, salinity and salt marsh plant zonation. Estuar. Coast. Shelf Sci. 62, 119–130 (2005).
Cui, B., He, Q., Zhang, K. & Chen, X. Determinants of annual–perennial plant zonation across a salt–fresh marsh interface: a multistage assessment. Oecologia 166, 1067–1075 (2011).
Yuan, Y. et al. Interspecific interactions between Phragmites australis and Spartina alterniflora along a tidal gradient in the Dongtan Wetland, Eastern China. PLoS ONE 8, e53843 (2013).
Holmer, M., Gribsholt, B. & Kristensen, E. Effects of sea level rise on growth of Spartina anglica and oxygen dynamics in rhizosphere and salt marsh sediments. Mar. Ecol. Prog. Ser. 225, 197–204 (2002).
Suchrow, S. & Jensen, K. Plant species responses to an elevational gradient in German North Sea salt marshes. Wetlands 30, 735–746 (2010).
Greenwood, M. E. & MacFarlane, G. R. Effects of salinity on competitive interactions between two Juncus species. Aquat. Bot. 90, 23–29 (2009).
Weigelt, A., Röttgermann, M., Steinlein, T. & Beyschlag, W. Influence of water availability on competitive interactions between plant species on sandy soils. Folia Geobot. 35, 169–178 (2000).
Emery, N. C., Ewanchuk, P. J. & Bertness, M. D. Competition and salt-marsh plant zonation: stress tolerators may be dominant competitors. Ecology 82, 2471–2485 (2001).
Davis, M. A. & Pelsor, M. Experimental support for a resource-based mechanistic model of invasibility. Ecol. Lett. 4, 421–428 (2001).
Fetene, M. Intra- and inter-specific competition between seedlings of Acacia etbaica and a perennial grass (Hyparrenia hirta). J. Arid Environ. 55, 441–451 (2003).
Montemayor, D., Canepuccia, A., Pascual, J. & Iribarne, O. Aboveground biomass patterns of dominant Spartina species and their relationship with selected abiotic variables in Argentinean SW Atlantic Marshes. Estuar. Coast. 1–10 (2013).
Zhang, J., Cheng, G., Yu, F., Kräuchi, N. & Li, M.-H. Intensity and importance of competition for a grass (Festuca rubra) and a legume (Trifolium pratense) vary with environmental changes. J. Integr. Plant Biol. 50, 1570–1579 (2008).
Swales, A., Ovenden, R., MacDonald, I. T., Lohrer, A. M. & Burt, K. L. Sediment remobilisation from decomposing cordgrass (Spartina anglica) patches on a wave-exposed intertidal flat. N. Z. J. Mar. Freshwat. Res. 39, 1305–1319 (2005).
Kim, D., Cairns, D. M. & Bartholdy, J. Tidal creek morphology and sediment type influence spatial trends in salt marsh vegetation. Professional Geographer 65, 544–560 (2013).
Medeiros, D. L., White, D. S. & Howes, B. L. Replacement of Phragmites australis by Spartina alterniflora: the role of competition and salinity. Wetlands 33, 421–430 (2013).
Sharma, G., Muhl, S., Esler, K. & Milton, S. Competitive interactions between the alien invasive annual grass Avena fatua and indigenous herbaceous plants in South African Renosterveld: the role of nitrogen enrichment. Biol. Invasions 12, 3371–3378 (2010).
Badgery, W. B., Kemp, D. R., Michalk, D. L. & King, W. M. C. G. Competition for nitrogen between Australian native grasses and the introduced weed Nassella trichotoma. Ann. Bot. 96, 799 (2005).
Levine, C. M. & Stromberg, J. C. Effects of flooding on native and exotic plant seedlings: implications for restoring south-western riparian forests by manipulating water and sediment flows. J. Arid Environ. 49, 111–131 (2001).
Manea, A. & Leishman, M. Competitive interactions between native and invasive exotic plant species are altered under elevated carbon dioxide. Oecologia 165, 735–744 (2011).
Anastasiou, C. J. & Brooks, J. R. Effects of soil ph, redox potential and elevation on survival of Spartina patens planted at a west central Florida salt marsh restoration site. Wetlands 23, 845–859 (2003).
Proffitt, C. E., Travis, S. E. & Edwards, K. R. Genotype and elevation influence Spartina alterniflora colonization and growth in a created salt marsh. Ecol. Appl. 13, 180–192 (2003).
Sundby, B., Vale, C., Caetano, M. & Luther, G. W. Redox chemistry in the root zone of a salt marsh sediment in the Tagus Estuary, Portugal. Aquat. Geochem. 9, 257–271 (2003).
Swales, A., MacDonald, I. T. & Green, M. O. Influence of wave and sediment dynamics on cordgrass (Spartina anglica) growth and sediment accumulation on an exposed intertidal flat. Estuaries 27, 225–243 (2004).
Adam Langley, J., Mozdzer, T. J., Shepard, K. A., Hagerty, S. B. & Patrick Megonigal, J. Tidal marsh plant responses to elevated CO2, nitrogen fertilization and sea level rise. Global Change Biol. 19, 1495–1503 (2013).
Boorman, L. Salt marshes – present functioning and future change. Mangroves Salt Marshes 3, 227–241 (1999).
Meier, C. I., Reid, B. L. & Sandoval, O. Effects of the invasive plant Lupinus polyphyllus on vertical accretion of fine sediment and nutrient availability in bars of the gravel-bed Paloma river. Limnologica 43, 381–387 (2013).
Jin, C.-W. et al. Aquatic plant debris improve phosphorus sorption into sediment under anoxic condition. Environ. Sci. Pollut. R. 1–8 (2013).
Lee, R. W. Physiological adaptations of the invasive cordgrass Spartina anglica to reducing sediments: rhizome metabolic gas fluxes and enhanced O2 and H2S transport. Mar. Biol. 143, 9–15 (2003).
Slocum, M., Mendelssohn, I. & Kuhn, N. Effects of sediment slurry enrichment on salt marsh rehabilitation: plant and soil responses over seven years. Estuaries 28, 519–528 (2005).
Tong, C., Baustian, J. J., Graham, S. A. & Mendelssohn, I. A. Salt marsh restoration with sediment-slurry application: effects on benthic macroinvertebrates and associated soil-plant variables. Ecol. Eng. 51, 151–160 (2013).
Mendelssohn, I. A. & Kuhn, N. L. Sediment subsidy: effects on soil–plant responses in a rapidly submerging coastal salt marsh. Ecol. Eng. 21, 115–128 (2003).
Koop-Jakobsen, K. The dynamics of sediment oxygenation in Spartina anglica rhizospheres-a planar optode study. Geophys. Res. Abstracts 15, EGU2013-12516 (2013).
Li, H. et al. Density-dependent effects on the dieback of exotic species Spartina anglica in coastal China. Ecol. Eng. 35, 544–552 (2009).
Chelaifa, H., Monnier, A. & Ainouche, M. Transcriptomic changes following recent natural hybridization and allopolyploidy in the salt marsh species Spartina × townsendii and Spartina anglica (Poaceae). New Phytol. 186, 161–174 (2010).
An, S. Q. et al. Spartina invasion in China: implications for invasive species management and future research. Weed Res. 47, 183–191 (2007).
Zhang, X. Y. et al. Response characteristics of Scirpus Triqueter and its rhizosphere to pyrene contaminated soils at different growth stages. Int. J. Phytoremediation 14, 691–702 (2011).
Li, H. L. et al. Nitrogen level changes the interactions between a native Scirpus triqueter and an exotic species Spartina anglica in coastal China. PLoS ONE 6, e25629 (2011).
Widdows, J., Pope, N. D. & Brinsley, M. D. Effect of Spartina anglica stems on near-bed hydrodynamics, sediment erodability and morphological changes on an intertidal mudflat. Mar. Ecol. Prog. Ser. 362, 45–57 (2008).
Bartholdy, A. T., Bartholdy, J. & Kroon, A. Salt marsh stability and patterns of sedimentation across a backbarrier platform. Mar. Geol. 278, 31–42 (2010).
Shaw, G. A., Adams, J. B. & Bornman, T. G. Sediment characteristics and vegetation dynamics as indicators for the potential rehabilitation of an estuary salt marsh on the arid west coast of South Africa. J. Arid Environ. 72, 1097–1109 (2008).
Lowe, B. J., Watts, R. J., Roberts, J. & Robertson, A. The effect of experimental inundation and sediment deposition on the survival and growth of two herbaceous riverbank plant species. Plant Ecol. 209, 57–69 (2010).
Wang, Q. et al. Effects of growing conditions on the growth of and interactions between salt marsh plants: implications for invasibility of habitats. Biol. Invasions 8, 1547–1560 (2006).
Zhi, Y. et al. Inter-specific competition: Spartina alterniflora is replacing Spartina anglica in coastal China. Estuar. Coast. Shelf Sci. 74, 437–448 (2007).
Chung, C. H., Zhuo, R. Z. & Xu, G. W. Creation of Spartina plantations for reclaiming Dongtai, China, tidal flats and offshore sands. Ecol. Eng. 23, 135–150 (2004).
Curado, G., Figueroa, E. & Castillo, J. M. Vertical sediment dynamics in Spartina maritima restored, non-restored and preserved marshes. Ecol. Eng. 47, 30–35 (2012).
Peng, R., Fang, C., Li, B. & Chen, J. Spartina alterniflora invasion increases soil inorganic nitrogen pools through interactions with tidal subsidies in the Yangtze Estuary, China. Oecologia 165, 797–807 (2011).
Wang, Y. J., Zhou, L. M., Zheng, X. M., Qian, P. & Wu, Y. H. Influence of Spartina alterniflora on the mobility of heavy metals in salt marsh sediments of the Yangtze River Estuary, China. Environ. Sci. Pollut. R. 20, 1675–1685 (2013).
Christiansen, T., Wiberg, P. L. & Milligan, T. G. Flow and sediment transport on a tidal salt marsh surface. Estuar. Coast. Shelf Sci. 50, 315–331 (2000).
Moskalski, S. M. & Sommerfield, C. K. Suspended sediment deposition and trapping efficiency in a Delaware salt marsh. Geomorphology 139–140, 195–204 (2012).
Loebl, M., van Beusekom, J. & Reise, K. Is spread of the neophyte Spartina anglica recently enhanced by increasing temperatures? Aquat. Ecol. 40, 315–324 (2006).
Cutajar, J., Shimeta, J. & Nugegoda, D. Impacts of the invasive grass Spartina anglica on benthic macrofaunal assemblages in a temperate Australian saltmarsh. Mar. Ecol. Prog. Ser. 464, 107–120 (2012).
Li, H. et al. Protogynous, pollen limitation and low seed production reasoned for the dieback of Spartina anglica in coastal China. Plant Sci. 174, 299–309 (2008).
Goldberg, D. E., Rajaniemi, T., Gurevitch, J. & Stewart-Oaten, A. Empirical approaches to quantifying interaction intensity: competition and facilitation along productivitty granients. Ecology 80, 1118–1131 (1999).
Wang, P., Lei, J. P., Li, M. H. & Yu, F. H. Spatial heterogeneity in light supply affects intraspecific competition of a stoloniferous clonal plant. PLoS ONE 7, e39105 (2012).
Wang, P., Xu, Y. S., Dong, B. C., Xue, W. & Yu, F. H. Effects of clonal fragmentation on intraspecific competition of a stoloniferous floating plant. Plant Biol., 10.1111/plb.12170 (2014).
Acknowledgements
This research was supported by the Fundamental Research Funds for the Central Universities (TD-JC-2013-1) and the National Natural Science Foundation of China (No. 31200313 and 31470475). We thank Dr. Shane Richards and a native English-speaking science editor (International Science Editing) and Peter Bridgewater for English edit.
Author information
Authors and Affiliations
Contributions
H.L.L. and S.Q.A. designed the experiment, H.L.L. and Y.B.Z. executed the experiment. H.L.L. and Y.Y.W. contributed to analyzing the data and making the figures. H.L.L., G.C.L. and M.X.Z. contributed to writing and editing the manuscript.
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Rights and permissions
This work is licensed under a Creative Commons Attribution-NonCommercial-ShareAlike 4.0 International License. The images or other third party material in this article are included in the article's Creative Commons license, unless indicated otherwise in the credit line; if the material is not included under the Creative Commons license, users will need to obtain permission from the license holder in order to reproduce the material. To view a copy of this license, visit http://creativecommons.org/licenses/by-nc-sa/4.0/
About this article

Cite this article
Li, HL., Wang, YY., An, SQ. et al. Sediment Type Affects Competition between a Native and an Exotic Species in Coastal China. Sci Rep 4, 6748 (2014). https://doi.org/10.1038/srep06748
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/srep06748
This article is cited by
-
Sand supplementation favors tropical seagrass Thalassia hemprichii in eutrophic bay: implications for seagrass restoration and management
BMC Plant Biology (2022)
-
Stoichiometric nutrient balance of Suaeda salsa wetlands in different supratidal habitats of Tianjin, China
Hydrobiologia (2019)
-
Integrating multiple vegetation indices via an artificial neural network model for estimating the leaf chlorophyll content of Spartina alterniflora under interspecies competition
Environmental Monitoring and Assessment (2017)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
