
Abstract
TSPYL2 is an X-linked gene encoding a nucleosome assembly protein. TSPYL2 interacts with calmodulin-associated serine/threonine kinase, which is implicated in X-linked mental retardation. As nucleosome assembly and chromatin remodeling are important in transcriptional regulation and neuronal function, we addressed the importance of TSPYL2 through analyzing Tspyl2 loss-of-function mice. We detected down-regulation of N-methyl-D-aspartate receptor subunits 2A and 2B (GluN2A and GluN2B) in the mutant hippocampus. Evidence from luciferase reporter assays and chromatin immunoprecipitation supported that TSPYL2 regulated the expression of Grin2a and Grin2b, the genes encoding GluN2A and GluN2B. We also detected an interaction between TSPYL2 and CBP, indicating that TSPYL2 may activate gene expression through binding CBP. In terms of functional outcome, Tspyl2 loss-of-function impaired long-term potentiation at hippocampal Schaffer collateral-CA1 synapses. Moreover, mutant mice showed a deficit in fear learning and memory. We conclude that TSPYL2 contributes to cognitive variability through regulating the expression of Grin2a and Grin2b.
Similar content being viewed by others

Introduction
Neurodevelopmental disorders, such as autism, schizophrenia and idiopathic learning disabilities, are more common or severe in males, where one likely cause is the involvement of X-linked genetic factors1,2. Given the heterogeneous nature of these disorders, functional studies using mutant mouse models are important for understanding the role of individual genes. TSPYL2, an X-linked gene that encodes a nucleosome assembly protein (NAP) in neurons, is a good candidate for neurodevelopmental disorders. TSPYL2 is also called CINAP (CASK-interacting NAP) as it binds calmodulin-associated serine/threonine kinase (CASK)3, which when mutated is clearly associated with X-linked mental retardation1,4.
The NAP domain binds histone for nucleosome remodeling, an important process in the regulation of gene expression5. Furthermore, NAPs have been proposed to regulate gene expression through bridging the transcriptional co-activator complex and chromatin6. The NAP superfamily is divided into NAP1-like (NAP1L), suppressor of variegation-enhancer of zeste-trithorax (SET) and Testis-specific protein, Y-encoded-like (TSPYL) families according to sequence homology of the NAP domain5. NAP1L1, NAP1L4 and SET (previously called NAP1, NAP2 and TAF, respectively) interact with histone acetyltransferase p300, a co-activator of gene expression6,7. p300 and the closely related CREB binding protein (CBP) cooperate with NAP1 to promote nucleosome eviction at the HTLV-1 promoter and transcriptional activation8. In differentiating neurons, NAP1L2 controls the expression of Cdkn1c by promoting histone H3K9/14 acetylation9. By contrast, SET is a subunit of the inhibitory complex of histone acetyltransferases10. SET negatively regulates the transcription of a subset of neuronal markers in neuroblastoma cells11. These studies demonstrate a role of NAPs in histone acetylation and transcriptional regulation.
Tspyl2 is expressed in neurons in multiple brain regions12. Hsueh and colleagues (2004) showed that TSPYL2 forms a complex with CASK and T-box brain gene 1 (Tbr-1), a transcription factor essential for cerebral cortex development. In primary hippocampal neurons, TSPYL2 activates the transcription of the Tbr-1 target gene Grin2b3. N-methyl-D-aspartate (NMDA) receptors are involved in memory performance13,14,15 and neurodevelopmental disorders16, therefore mutations affecting their expression might be expected to result in defects in these processes. However, Hsueh's group subsequently reported that mice homozygous for a targeted mutation of Tspyl2 exhibit normal levels of GluN2B in various brain regions; and no learning and memory defects as expected for a reduction in NMDA receptor function was detected17. We have generated an independent null mutant allele of Tspyl2 (Tspyl2tm1.Sich, synonym Tspyl2m) on a different genetic background18. In contrast to the earlier study, here we show that TSPYL2 is an important transcriptional regulator of both Grin2a and Grin2b. We also found that Tspyl2 mutant mice indeed exhibit deficits in both long-term potentiation (LTP) and fear-associative learning.
Results
TSPYL2 regulates the levels of GluN2A and GluN2B in hippocampus
We have previously reported that Tspyl2 is expressed in the cortex and hippocampus of adult mice and that both the size and gross morphology of the Tspyl2 mutant brain are normal18. Nissl staining on adult forebrain slices showed normal neuroanatomy in the mutant brain (Fig. 1A). To determine whether the expression of specific glutamate receptors is affected by the Tspyl2 mutation, we examined the protein levels of the key glutamate receptor subunits, including NMDA receptor subunits GluN1, GluN2A, GluN2B, α-amino-3- hydroxy-5-methylisoxazole -4-propionic acid (AMPA) receptor subunit GluA1 and metabotropic glutamate receptor subtype mGluR5 in the mutant hippocampus. Western blot analysis and densitometry scans revealed that the levels of GluN2A and GluN2B were reduced significantly in the mutant hippocampus (p < 0.05), whereas the levels of the other glutamate receptors were unaffected (Fig. 1B). To test whether this is due to reduced transcript levels, quantitative RT-PCR was performed and the results indicated that the levels of GluN2A (Grin2a) and GluN2B (Grin2b) mRNA were reduced in the mutant hippocampus (p < 0.05). As expected, the mRNA levels of the other glutamate receptors were similar between the wild-type and mutant. Interestingly, the transcript level of Reln, a Tbr-1 target gene, was also unaffected (Fig. 1C). Since the transcript levels of Grin2a and Grin2b were reduced in the mutants, we wondered whether it was due to reduced transcription or reduced mRNA stability. RNA stability assays were performed by adding actinomycin D to block transcription in primary neuron cultures derived from wild-type and mutant hippocampi. From quantitative RT-PCR, the degradation rates of Grin2a and Grin2b transcripts in the mutant hippocampal neurons were similar to that of the wild-type (Fig. 1D). Together, these data suggest that TSPYL2 is important for Grin2a and Grin2b transcription.

Reduced expression of Grin2a and Grin2b in Tspyl2 mutant hippocampus.
(A), Nissl staining of coronal brain sections from 2 month-old mice. Cellular composition of hippocampal substructures and layer development of cortex were indistinguishable between wild-type and mutant. Scale bars: 100 μm. (B), The protein level of key glutamate receptors in 2-month old hippocampi was detected by western blot and quantitated by densitometry. Cropped gel images are shown and the gels were run under the same experimental conditions. The protein level was normalized to actin and the wild-type level was set as 1. The protein levels of GluN2A and GluN2B were significantly reduced (n = 4 mice per genotype). (C), Transcript levels of the above genes in 2- month old hippocampi were detected by quantitative RT-PCR. Relative mRNA level to Hprt in the wild-type was set as 1. Expressions of Grin2a and Grin2b were reduced significantly in the mutant (n = 4 mice per genotype). Expression of Reln was unchanged. (D), RNA stability of Grin2a and Grin2b was determined by adding 10 μg/ml actinomycin D to hippocampal cultures at 7 days in vitro. Quantitative RT-PCR was done on samples collected at 0, 4, 8, 16 and 24 hr after treatment. No difference between wild-type and mutant neurons was observed (n = 3 samples per genotype). Error bars represent SEM. *P < 0.05, Student's t-test. Abbreviations: +/Y, wild-type male; m/Y, mutant male; WT, wild-type.
TSPYL2 activates transcription of Grin2a and Grin2b
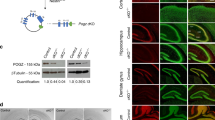
TSPYL2 is a NAP3, which is expected to function in the nucleus to regulate the expression of multiple genes. To confirm the nuclear localization of TSPYL2, we transfected cells with plasmids expressing TSPYL2 tagged either with HA at the N-terminus or GFP at the C-terminus. The staining patterns of both forms of tagged-TSPYL2 were the same and the result for HA-TSPYL2 is shown in Fig. 2A. Tagged-TSPYL2 was localized in the nucleus in both the neuroblastoma-glioma fusion cell line NG108-15 and primary hippocampal neurons. TSPYL2 could also be observed in the cytoplasm in primary hippocampal neurons, but not NG108-15, upon prolonged exposure. The data show that TSPYL2 is mainly localized in the nucleus.

TSPYL2 activates the transcription of Grin2a and Grin2b.
(A), Immunocytochemistry of HA-tagged TSPYL2 in NG108-15 cells and primary hippocampal neurons. Exogenous TSPYL2 was localized in the nucleus. Low expression was observed in the cytoplasm in neurons. Scale bar: 10 μm. (B), Luciferase reporter assay of Grin2a promoter in NG108-15 cells and primary hippocampal neurons. Reln promoter served as a negative control. HA-TSPYL2 repressed the promoter activity of Grin2a in NG108-15 but activated it in hippocampal neurons (n = 3 independent experiments). (C), Chromatin immunoprecipitation (ChIP) of Grin2a and Grin2b promoters in transfected NG-108-15 cells. The signal was normalized to signal from input DNA and the ChIP IgG level was set as 1. Both promoters were pulled down by anti-HA antibody in HA-TSPYL2 transfected cells. p21 promoter was used as a negative control. HA-SUN2 transfected cells were used to show the specificity of HA-TSPYL2 binding (n = 3 independent transfections). The sizes of sonicated DNA fragments were concentrated between 200–600 bp (Right). (D), Co-localization between HA-TSPYL2 and endogenous p300 or CBP in primary hippocampal neurons by confocal microscopy. Scale bar: 10 μm. (E), Interaction between TSPYL2 and CBP in mammalian-two-hybrid assay. Primary hippocampal neurons were transfected with various plasmids with GAL4 DNA binding domain (BD) or VP16- activation domain (AD) (n = 3 independent experiments). (F), ChIP of Grin2a and Grin2b promoters in primary hippocampal neurons cultured for 14 days (n = 3 independent samples). Error bars represent SEM. *P < 0.05, ** P < 0.01, ***P < 0.001, Student's t-test.
From luciferase reporter assays, TSPYL2 was found to activate the Grin2b promoter in primary hippocampal neurons, but to inhibit it in Neuro-2A neuroblastoma cells3. To investigate whether TSPYL2 also regulates the expression of Grin2a, we performed luciferase reporter assays using the Grin2a promoter. In NG-108-15, the activity of the Grin2a promoter was 10 fold that of the pGL3-basic vector. Co-transfection with an HA-TSPYL2 expression plasmid inhibited the activity of the Grin2a promoter by about 0.3 fold compared to the control with co-transfection of pcDNA3 (p < 0.05). In contrast, with primary neurons, the activity of the Grin2a promoter was 3 fold that of the pGL3-basic vector, while co-transfection with the HA-TSPYL2 expression plasmid enhanced the promoter activity of Grin2a by about 8.6 fold when compared to the control with co-transfection of pcDNA3 (p < 0.05). As a negative control, the activity of the Reln promoter was 200 and 5 fold that of pGL3-basic vector in NG108-15 and primary hippocampal neurons, respectively. Transfection of HA-TSPYL2 had no significant effect on the activity of the Reln promoter (Fig. 2B).
Next, we tested whether TSPYL2 was tethered to the native Grin2a and Grin2b promoters by using chromatin immunoprecipitation (ChIP) with cross-linking. We transfected NG108-15 cells with HA-tagged TSPYL2 and performed ChIP using IP-grade antibodies against HA. Both Grin2a and Grin2b promoters, but not the negative control p21 promoter, were pulled down by the HA antibody (Fig. 2C). As a negative control, cells were transfected with HA-tagged SUN2, a nuclear envelope protein, which should not bind any promoter. In this case the Grin2a and Grin2b promoters could not be detected by ChIP using the HA antibody. These data show that the binding of TSPYL2 to the Grin2a and Grin2b promoters is specific.
NAP1L1, NAP1L4 and SET have been reported to interact with CBP or p3006,7,19. Therefore we wondered whether TSPYL2 was recruited to the Grin2a and Grin2b promoters through CBP or p300. We found colocalization of immunofluorescence signals for HA-tagged TSPYL2 with endogenous CBP and p300 in primary hippocampal neurons (Fig. 2D). Next, we detected functional interaction between TSPYL2 and CBP in a mammalian two-hybrid assay with primary hippocampal neurons (Fig. 2E). As expected from the histone acetyltransferase activities of p300 and CBP, both GAL4-binding domain (BD)-p300 and BD-CBP activated the reporter activity. Together with the expression plasmid for activation domain (AD)-TSPYL2, the reporter activity doubled for BD-CBP (p < 0.01) but no change was observed for BD-p300 (p = 0.69). Lastly, the importance of TSPYL2 in the assembly of the transcriptional complex in neurons was tested by comparing the binding of p300 and CBP to promoter regions in wild-type and mutant hippocampal neurons. In mutant primary neurons, ChIP analysis revealed significantly reduced binding of p300 (p < 0.05) and CBP (p < 0.001) in the Grin2a and Grin2b promoter, respectively (Fig. 2F). Possibly, TSPYL2 activates transcription by interacting with p300 or CBP and bridges the transcriptional complex and histones as shown for several other NAPs6,7,19.
Long-term potentiation is impaired in Tspyl2 mutant neurons
NMDA receptors are important in synaptic function and memory. Using whole-cell patch clamp electrophysiological techniques, the basal excitability of CA1 neurons in both mutant and wild-type mice was examined. The ability of the neurons to fire action potentials in response to current injection as well as the input resistance of the neurons were normal in the mutant (Fig. 3A & B). Furthermore, in hippocampal slices prepared from wild-type and mutant mice, field excitatory postsynaptic potentials (fEPSPs) in the CA1 region were evoked by stimulating the Schaffer collaterals. The basal synaptic transmission, as assessed by the output-input relationship (fEPSP vs stimulation intensity) and the fEPSP to fiber volley, was not affected by the mutation (Fig. 3C). The paired-pulse ratio of the fEPSP also did not change, indicating a lack of effect on short-term synaptic plasticity (Fig. 3D). These results were consistent with the finding that the expression of the AMPA receptor subunit GluA1 was not affected. To examine potential defects in long term plasticity in Tspyl2 mutant mice, fEPSPs at the hippocampal Schaffer collateral-CA1 synapses were assessed using the conventional high frequency stimulation induction protocol consisting of a 1 second train of 100 Hz stimulation. When compared with wild-type, both male and female mutant mice showed a significant reduction in LTP. The impairment was 18% in male and 30% in female (p < 0.001; Figs. 3E & F). The normal membrane excitability and basal synaptic transmission in the mutant hippocampus implicated that the impairment in LTP was not due to defects in transmitter release machinery or fast AMPA receptors, but were in line with postsynaptic problems such as NMDA receptor malfunction.

Tspyl2 mutation impairs long term potentiation (LTP).
(A), Membrane excitability of hippocampal CA1 neurons was measured by whole-cell patch-clamp recording in 2-month old mice. Series of current steps between −160 pA and +160 pA (−55 mV) in 20 pA increments were applied. No difference was observed between wild-type and mutant in action potential (n = 4 each). Three representative recorded action potentials (40, 100, 160 pA) were shown on the right. (B), Input resistance of hippocampal CA1 neurons was measured by whole-cell patch-clamp recording in 2- month old mice at a current of −60 mV. No significant difference was observed between wild-type and mutant (n = 43 for +/Y, n = 34 for m/Y). (C), Basal synaptic transmission was compared by the field excitatory post-synaptic potential (fEPSP)-stimulation relationship. No significant differences were observed between the two groups throughout the range of stimulation intensities. There was also no difference between the fEPSP to fiber volley ratio between the two groups (10 slices from 3 +/Y mice, 12 slices from 3 m/Y mice). (D), The paired-pulse ratio of the evoked fEPSP was not affected by the mutation (10 slices from 3 +/Y mice, 12 slices from 3 m/Y mice). (E), LTP of 2-month old male and female mice. FEPSP was measured from the dendritic layer of CA1 neurons in the Schaffer collateral pathway. Period 1 was the baseline fEPSP while period 2 was the fEPSP after induction. Arrow indicated the electrical stimulation (1 train, 100 Hz, 1 s). Representative recorded potentials were shown on the top. (F), Quantitative change in the average amplitude of the fEPSP taken from 50 to 60 min after induction. LTP was impaired in the mutant (n = 12 for male, n = 9 for female). Error bars represent SEM. ***P < 0.001, Student's t-test. Abbreviations: AP, Action potential; +/Y, wild-type male; m/Y, mutant male; +/+, wild-type female; +/m, heterozygous female; m/m, mutant female.
TSPYL2 is important in fear-associative learning
As the molecular defect in the Tspyl2 mutant neuron leads to impaired LTP, we searched for learning deficits in the mutant mouse. Our mutant mice are on a 129/SvEv background, which are good learners for learning and memory tests20. We therefore performed Morris water maze tests to assess spatial memory and contextual fear conditioning tests to assess fear-associative memory. Our analysis demonstrated that Tspyl2 mutant mice have normal vision and swim speed during visual tests (Fig. 4A) and behaved similarly to wild-type in our Morris water maze settings for reference memory and probe tests (Fig. 4B, C). Both genotypes learned with the escape latency decreased significantly across trials in the reference memory test (Days: F2,24 = 38.46, p < 0.001; genotype: F1,24 = 1.06, p = 0.323 by repeated measures ANOVA) and increased time spent in the target quadrant in the probe test. To be eligible for fear conditioning tests, mutant mice were first examined and shown to have normal pain sensation (Fig. 4D). In addition, exploratory activity during habituation was not significantly different between genotypes (Genotype: F1,242 = 1.95, p = 0.176; time: F11,242 = 3.76, p < 0001; interaction: F11,242 = 2.42, p < 0.05 by repeated measures ANOVA, Fig. 4E, left). Noticeably, Tspyl2 mutant mice exhibited significantly impaired contextual fear conditioning (p < 0.05, Student's t-test, Fig. 4E, right), indicating a deficit in fear-associative memory.

Normal spatial memory but impaired fear-associative memory in Tspyl2 mutant mice.
(A–C), Tspyl2 mutant mice perform normally in Morris water maze (n = 7 per genotype). (A), Visual test was performed in clear water. There was no significant difference between genotypes in swim speed (p = 0.160) and latency (p = 0.613) to find the platform. (B), Spatial reference memory was tested with a fixed, hidden platform. Mutant mice learned normally. (C), Probe test was done by removing the platform after the spatial reference memory test. Wild-type and mutant mice spend similar time in the target quadrant Q2. (D), Normal pain sensation in hot plate test (n: +/Y = 15, m/Y = 5) as one of the prerequisites for fear conditioning test. (E), Results of fear conditioning tests. Mutant and wild-type mice did not differ significantly in exploratory activities during the 6 min habituation on day 1 (left). The freezing rate during habituation was shown as basal. After conditioned stimulus-unconditioned stimulus training (shock) on day 1, the mice were tested in a novel chamber with the same tone presented (cue) on day 2 and in the training chamber without tone stimulus (context) on day 3. As reflected by freezing behavior, mutant mice showed significantly impaired contextual fear conditioning (Error bars represent SEM, *p < 0.05, Student's t-test, n = 12 per group). Abbreviations: +/Y, wild-type male; m/Y, mutant male.
Discussion
TSPYL2 is within the chromosome region linked to neurodevelopmental syndromes21,22,23. In this study, we investigated whether TSPYL2 plays a role in cognitive function through transcriptional regulation of neuron-specific genes. We examined the expression of glutamate receptors in the hippocampus, a key brain area involved in learning and memory24, in Tspyl2 mutant mice and found a reduction in the expression of genes encoding the GluN2A and GluN2B subunits in the mutant hippocampus. Furthermore, ChIP analysis indicated that Grin2a and Grin2b promoters are indeed targets of TSPYL2. Our data illustrate the importance of TSPYL2 in the transcriptional regulation of both GluN2A and GluN2B subunits and suggest a role for TSPYL2 in learning and memory.
NMDA receptors play a critical role in some forms of synaptic plasticity and learning13. Mice with loss of GluN2A25 or GluN2B26 show impaired hippocampal LTP and impaired spatial memory in the Morris water maze. In our Tspyl2 mutant mice, which show reduced expression of both GluN2A and GluN2B instead of total loss of either one receptor subunit, we did not identify spatial memory defects in a conventional water maze paradigm, but we did observe disrupted contextual fear conditioning. Other tests of spatial learning with different sensorimotor and motivational demands, such as appetitive motivated maze tasks, may reveal deficits in mutant mice having normal water maze performance27,28. For the fear conditioning task, it is dependent on amygdala-hippocampal function29,30. We attribute the defects in our mice at least partly to disrupted NMDA signaling in the hippocampus because this is important in the process of fear conditioning31. Recently, de novo mutations in GRIN2A and GRIN2B affecting protein functions were identified in individuals with mental retardation16 and autism spectrum disorders32. Reduction or loss of function of GluN2A and GluN2B, together with other genetic factors, is likely to cause variable neurodevelopmental phenotypes.
How does TSPYL2 activate the transcription of both Grin2a and Grin2b in the hippocampus? Previously it has been shown that TSPYL2 forms a complex with Tbr-1 through interaction with CASK and activates the Grin2b promoter3. Our finding of TSPYL2 regulating the transcription of Grin2a is novel. As there are multiple reports of direct interactions between NAPs and CBP or p3006,7,8,33, recruitment of TSPYL2 to the various promoters by transcription regulators such as CASK, p300 and CBP may be a general mechanism. In return, TSPYL2 will help to anchor the transcriptional complex to chromatin through its binding to histone. In future, the availability of antibodies for specific immunoprecipitation of TSPYL2 will allow us to further confirm the binding of TSPYL2 to the promoters of Grin2a and Grin2b in specific brain regions, as well as to identify other in vivo gene targets by ChIP-sequencing. In addition, insights into how TSPYL2 regulates the transcription of the Grin2a promoter can be gained by identifying other interacting proteins of TSPYL2, including transcription factors and transcription regulators.
CBP interacts with over 400 transcription factors34 and its importance for memory formation is well established35,36,37,38. A recent study shows that p300 is required for recognition memory and contextual fear memory39. Interestingly, we found co-localization of HA-TSPYL2 with CBP and p300 in primary hippocampal neurons and we have confirmed this result in various cell lines. Our data from mammalian-two-hybrid assays further suggested functional interaction between TSPYL2 and CBP. However, we cannot find evidence for an interaction between p300 and TSPYL2 even though they co-localized. One possibility is the transient or weak nature of their interaction. To address this we investigated the datasets from a proteomic study designed to detect weak protein interactions by utilizing high levels of reciprocity in 3290 immunoprecipitations with transcriptional coregulators in cell lines. TSPYL1 is found to form a stable complex with TSPYL2 and this complex interacts with p300. On the other hand, multiple immunoprecipitations with CBP or p300 antibodies did not reveal a steady-state stoichiometric partner40. Taken together, we propose that TSPYL2 interacts with CBP and p300 to regulate transcription in neurons.
The phenotypic outcome of a mutation can be modified by other genes and varies between genetic backgrounds41,42. With another Tspyl2 targeted mutation in mice on a C57BL/6 background, both the level of GluN2B in various brain regions and fear-conditioning were reported to be normal while locomotor activities are increased17. We only observed a marginal increase in locomotion in terms of swim speed (p = 0.160) and exploration (p = 0.176) in our mutant mice. The targeted allele in both studies is likely to be a null allele. The full length TSPYL2 protein was eliminated and there was no abnormal protein being detected in theirs17 or in our mutant mouse brain (Supplementary Fig. 1). Differences in phenotype among inbred strains are not unexpected due to the polymorphisms in genetic modifiers and these may contribute to the different patterns of results across studies. For example, all 129 inbred substrains have a deletion polymorphism in exon 6 of Disc1 (Disrupted in schizophrenia 1) gene43, which results in removal of one of the isoforms of DISC144. These mice do not have working memory defects which were observed when their Disc1 allele was transferred to the C57BL/6J inbred background45. Besides behavior, the genetic background can also have a major affect on the phenotypes seen after mutating specific genes important in development, for example the gene for epidermal growth factor receptor46.
In summary, our data highlights the role of TSPYL2 in regulating the expression level of multiple genes in the brain, thereby playing an important role in shaping learning and behavior. Further molecular and electrophysiological studies of TSPYL2 function in the amygdala and hypothalamus, two brain areas besides the hippocampus with highest Tspyl2 expression17, may reveal more robust target genes and functions of TSPYL2. In summary, the demonstration here of reduced GluN2A and GluN2B expression, impaired LTP and fear-conditioning in our TSPYL2 mutant mice prove that TSPYL2 contributes to cognitive functions. Our mutant mice will be useful for further understanding of the role of TSPYL2 in neuronal function.
Methods
Animals
Tspyl2 mutant mice were maintained in a pure 129Sv/Ev background18. Nissl staining was performed on coronal brain sections collected from perfusion fixed animals. Mouse experiments were approved by the Committee on the Use of Live Animals in Teaching and Research at the University of Hong Kong (Approval no. 1643-08 and 2612-11). All experiments were performed in accordance with the relevant guidelines and regulation of the Laboratory Animal Unit at the University of Hong Kong, which has full accreditation with Association for Assessment and Accreditation of Laboratory Animal Care (AAALAC).
Cell culture
Primary hippocampal neurons were isolated from 16.5 days embryos and seeded in Neurobasal Medium (Invitrogen) supplemented with B27 (Invitrogen), 1% FBS and penicillin/streptomycin on poly-L-lysine coated culture dishes. The seeding cell density was 600 cells/mm2 for immunocytochemistry and 750 cells/mm2 for luciferase assay. NG108-15 cells were cultured in DMEM supplemented with 10% FBS.
Primers
The sequences of forward and reverse primers (5′ to 3′) were as follows:
Grin1: GGAAGAACCTGCAGGATAGA; CTCGTGTCTTTGGAGGACCT;
Grin2a: CATCAGCAGGGGCATCTACA; GGGTTGGACTCATTGAGAGT;
Grin2b: CCATCAGCAGAGGTATCTAC; CAGTCTGAATGCGTGAAGCT;
Gria1: ACATTTCCCCCAGGTCCCTGTCTG; AACCGCTAGGTTTACGGGACCTCT;
Grm5: GACGACTTGACAGGTTGTGA; GATCTTCTTCTTGCTGCCAG;
Reln: GAAAGCTTCCAAGGTGACGA; AGTGCTTACTAGGACGACCT;
Hprt: AACTGGAAAGAATGTCTTGATTG; TCAAATCCAACAAAGTCTGGC.
Primer for promoters:
Grin2a: CGGAGAGCGTGGTTTCAGCA; GGAACAAGGCCGACCTAGGT;
Grin2b: CACACCCTGCCTCTTGGGTTTC; GTCCTGGTCTTGATTTGGGTCT;
p21: AGTCCTGGGTGGGGACTAGCT; CCACCCTGCACTGAAGCAGCC.
Antibodies
The sources of antibodies were: actin (A5060), GluN1 (G8913), GluN2A (M264), HA (H3663 for immunocytochemistry) from Sigma; GluA1(L844), mGluR5 (K56) from bioworld; GluN2B (06-600) from Upstate; CBP (sc-369), HA (sc-805 for ChIP) and p300 (sc-585) from Santa Cruz.
Plasmids
A cDNA clone containing full length human TSPYL2 was obtained by library screening using a partial fragment isolated previously47. The entire coding region of TSPYL2 (693 amino acids) was PCR-amplified using forward and reverse primers, 5′-ACGGAATTCATGGACCGCCCAGATGAGGG-3′ and 5′-ACGGTCGACAATCCGGTTTTCCCCCTCTTCC- 3′. The purified PCR product was cloned into pT-Adv (Clontech), then subcloned into EcoRI/SalI sites of pEGFP-N1 (Clontech) and a modified pcDNA3 vector (Invitrogen) with the HA tag inserted. All plasmids were verified by sequencing. Plasmid of HA-SUN2 was cloned as described previously48. Firefly reporter plasmids Grin2a-1253 containing 1253 bp upstream of translation start was kindly provided by Dr. Andres Buonanno49; and Reln-514 containing 514 bp upstream of transcription start site by Dr. Dennis R. Grayson50. Plasmids for mammalian two-hybrid assay (pG4-p300, pG4-CBP) were gifts from Dr. Neil D Perkins51; pG5-luc containing GAL4 DNA binging sites, pACT containing the AD of VP16, pRL-TK were from Promega. AD-TSPYL2 was constructed by subcloning full length TSPYL2 cDNAs into the EcoRI/SalI sites of pACT.
Immunoblotting
Hippocampi were collected from 2-month old male littermate mice in ice-cold HBSS (Sigma). Tissue was lysed with a dounce homogenizer in RIPA buffer supplemented with complete protease inhibitors (Roche) and 100 μM MG132 (Sigma). Proteins (50 μg) were resolved and detected with standard immunoblotting procedures and ECL reagents (Millipore). Quantitation of the protein bands was done with software from GeneTools (Syngene).
Quantitative RT-PCR
RNA was extracted using Trizol solution (Invitrogen). Two μg of total RNA was reverse transcribed in 20 μl with Oligo(dT). Quantitative PCR was done with 0.5 μl of cDNA by using QuantiFast SYBR green PCR kit (Qiagen) in 7900 HT Fast Real-Time PCR System (Applied Biosystems). For RNA stability assay, hippocampal neurons collected from littermate embryos were cultured for 7 days. Neurons were then incubated with 10 μg/ml actinomycin D (Sigma) to inhibit transcription. RNA samples were collected at 0, 4, 8, 16 and 24 hr after treatment.
Immunocytochemistry
Cells grown on coverslips coated with gelatin or poly-L-lysine were washed with PBS and fixed in 4% paraformaldehyde for 15 min. Cells were permeabilized with 0.1% Triton X-100 in PBS for 10 min. Standard staining procedures with Alexa Flour 488 or 594 labeled-goat antibodies (Invitrogen) were followed. The images were acquired with epifluorescence microscope (Carl Zeiss, Axioplan 2 imaging) or LSM710 Meta laser scanning confocal microscope (Carl Zeiss). The setting of the confocal microscope was 40X oil objective lens; numerical aperture 1.4.
Luciferase assay and mammalian two-hybrid assay
Expression plasmids were co-transfected with the firefly reporter plasmid (Grin2a-1253 or Reln-514) and TK-Renilla reporter pRL-TK into NG108-15 cells (in 24-well plates) or hippocampal neurons at 6 days in culture (in 12-well plates) using Lipofectamine 2000 (Invitrogen). Lysates were prepared 24–48 hr after transfection and measured with Dual-Glo Luciferase assay system (Promega). Firefly luciferase activities were standardised to the corresponding Renilla luciferase activities. For mammalian two-hybrid assay, plasmid concentrations were optimised to give ~ 1:1 molar ratio of AD-TSPYL2 and interacting proteins to be tested. Primary hippocampal neurons were cultured for 6 days in 12-well plates and transfected by calcium phosphate precipitation with a total of 2 μg of plasmids containing 200 ng pRL-TK, 800 ng pG5-luc, 300 ng of AD-TSPYL2 together with 700 ng of pG4-p300 or pG4-CBP. Assays were carried out 24 hours after transfection using Dual Luciferase assay system (Promega). Experiments were performed in triplicate and n = the number of independent experiments.
Chromatin immunoprecipitation
Cells were grown on 10 cm dishes. For primary neurons, they were collected from the whole litter of wild-type or mutant embryos obtained on the same day and cultured for 14 days in parallel. Cells were cross-linked by 1.3% formaldehyde for 10 min and harvested in lysis buffers with protease inhibitors and sonicated (Soniprep 150, MSE). An aliquot of chromatin from 1 million cells was used for each IP, washed five times with RIPA buffer and once with TE buffer. Standard protocols were then followed for DNA extraction and qPCR52. Promoter regions −1002 to −917 of Grin2a and −291 to −132 of Grin2b were amplified.
Electrophysiology
Two-month old littermate animals were employed and the detailed procedures were described previously53. Briefly, 250 μm thick parasagittal sections were maintained at 34°C for whole cell patch-clamp recordings. Pipettes with low KCl internal solution was used to record the electrophysiological properties of hippocampal CA1 neurons. The holding current was adjusted until the membrane potential was held at −70 mV or −50 mV. For characterization of the membrane excitability and properties of the action potentials, a series of current steps between −160 pA and +160 pA in 20 pA increments were applied. To study the basal synaptic transmission and E-LTP, 300 μm hippocampal slices were prepared. To increase the efficiency and to minimize variations in the results arising from differences in incubation times, a maximum of four slices were studied simultaneously. The slices were placed on probes fabricated with 8 × 8 electrode arrays. FEPSPs were recorded from the dendritic layer of CA1 neurons by choosing an electrode in the Schaffer collateral pathway as the stimulating electrode. Since there was no difference in the size of fiber volley evoked by various stimulation currents between the wild-type and the mutant, the basal synaptic transmission was compared by assessing the fEPSP-stimulus current relationship and also the fEPSP to fiber volley ratio. Furthermore, paired-pulse ratio with a chosen stimulus interval of 50 ms was compared. For the LTP, based on the stimulus-response curve, we chose a stimulation intensity that evoked the fEPSP with a magnitude of 30–40% of the maximum response (around 1 mV in most cases). After allowing a stable baseline of 30 min, 1 train of 100 Hz stimulus that lasted for 1 s was applied and the field potential response for 1 hr after the tetanus was recorded. LTP was quantified as % change in the average amplitude of fEPSPs taken from 50 to 60 min interval after the induction, compared with that of baseline.
Morris water maze
Two to three months old male littermate mice were subject to reverse light-dark cycle 1 week prior to tests which were performed in the dark cycle. Mutant mice were confirmed to be of normal vision and swimming ability in the visual test, where a platform was put in clear water in standard water maze settings. For the reference memory test, a hidden platform was placed in a fixed location in milk water and visual aids were provided on the walls. The training was carried out in blocks of four trials per day for 3 days. Probe trials were performed on day 4 with the hidden platform removed. The moving path and escape latency for reaching the platform were recorded by camera. Data collection and analysis were performed using the video-tracking system EthoVision XT 7.1.
Fear conditioning
Mice were checked for normal pain sensation by the hot plate test. The surface of a hot plate was warmed to 55°C and the latencies for the mice to lick their hind paws were recorded. For fear conditioning tests, two-month old male littermate mice were tested in the dark cycle. On day 1, mice were placed in the conditioning chamber (25 cm × 25 cm × 25 cm) for 6 min habituation before training with conditioned stimulus-unconditioned stimulus (CS-US) pairing protocol as described previously54. Briefly, the mice were presented with three tone-foot shock pairing trials (CS: 75 dB for 30 s, US: 0.5 mA for 2 s) with inter-trial interval of 2 min. On day 2, cued fear memory was tested. Mice were placed in a novel chamber for 6 min (pre-CS) and then presented with three CS (75 dB for 30 s). On day 3, contextual fear memory was tested by placing the mice in the training chamber without stimulus for 5 min. The freezing behavior was recorded and analysed with EthoVision XT 7.1.
Data analysis
Data were analyzed by repeated-measure ANOVA or two-sided unpaired Student's t-test. P < 0.05 was considered statistically significant.
References
Tarpey, P. S. et al. A systematic, large-scale resequencing screen of X-chromosome coding exons in mental retardation. Nat Genet 41, 535–543 (2009).
Piton, A. et al. Systematic resequencing of X-chromosome synaptic genes in autism spectrum disorder and schizophrenia. Mol Psychiatry 16, 867–880 (2011).
Wang, G. S. et al. Transcriptional modification by a CASK-interacting nucleosome assembly protein. Neuron 42, 113–128 (2004).
Hackett, A. et al. CASK mutations are frequent in males and cause X-linked nystagmus and variable XLMR phenotypes. Eur J Hum Genet 18, 544–552 (2010).
Park, Y. J. & Luger, K. Structure and function of nucleosome assembly proteins. Biochem Cell Biol 84, 549–558 (2006).
Shikama, N. et al. Functional interaction between nucleosome assembly proteins and p300/CREB-binding protein family coactivators. Mol Cell Biol 20, 8933–8943 (2000).
Asahara, H. et al. Dual roles of p300 in chromatin assembly and transcriptional activation in cooperation with nucleosome assembly protein 1 in vitro. Mol Cell Biol 22, 2974–2983 (2002).
Sharma, N. & Nyborg, J. K. The coactivators CBP/p300 and the histone chaperone NAP1 promote transcription-independent nucleosome eviction at the HTLV-1 promoter. Proc Natl Acad Sci U S A 105, 7959–7963 (2008).
Attia, M., Rachez, C., De Pauw, A., Avner, P. & Rogner, U. C. Nap1l2 promotes histone acetylation activity during neuronal differentiation. Mol Cell Biol 27, 6093–6102 (2007).
Seo, S. B. et al. Regulation of histone acetylation and transcription by INHAT, a human cellular complex containing the set oncoprotein. Cell 104, 119–130 (2001).
Kim, D. W., Kim, K. B., Kim, J. Y., Lee, K. S. & Seo, S. B. Negative regulation of neuronal cell differentiation by INHAT subunit SET/TAF-Ibeta. Biochem Biophys Res Commun 400, 419–425 (2010).
Lin, C. W. et al. Neural activity- and development-dependent expression and distribution of CASK interacting nucleosome assembly protein in mouse brain. J Comp Neurol 494, 606–619 (2006).
Bannerman, D. M., Rawlins, J. N. & Good, M. A. The drugs don't work-or do they? Pharmacological and transgenic studies of the contribution of NMDA and GluR-A-containing AMPA receptors to hippocampal-dependent memory. Psychopharmacology (Berl) 188, 552–566 (2006).
Bliss, T. V. & Collingridge, G. L. A synaptic model of memory: long-term potentiation in the hippocampus. Nature 361, 31–39 (1993).
de Quervain, D. J. & Papassotiropoulos, A. Identification of a genetic cluster influencing memory performance and hippocampal activity in humans. Proc Natl Acad Sci U S A 103, 4270–4274 (2006).
Endele, S. et al. Mutations in GRIN2A and GRIN2B encoding regulatory subunits of NMDA receptors cause variable neurodevelopmental phenotypes. Nat Genet 42, 1021–1026 (2010).
Chung, W. C., Huang, T. N. & Hsueh, Y. P. Targeted deletion of CASK-interacting nucleosome assembly protein causes higher locomotor and exploratory activities. Neurosignals 19, 128–141 (2011).
Tao, K. P. et al. TSPYL2 is important for G1 checkpoint maintenance upon DNA damage. PLoS One 6, e21602 (2011).
Karetsou, Z., Martic, G., Sflomos, G. & Papamarcaki, T. The histone chaperone SET/TAF-Ibeta interacts functionally with the CREB-binding protein. Biochem Biophys Res Commun 335, 322–327 (2005).
Wehner, J. M. & Silva, A. Importance of strain differences in evaluations of learning and memory processes in null mutants. Ment Retard Dev Disabil Res Rev 2, 243–248 (1996).
Dann, J. et al. A linkage study of schizophrenia to markers within Xp11 near the MAOB gene. Psychiatry Res 70, 131–143 (1997).
Annunziata, I. et al. Mapping of MRX81 in Xp11.2-Xq12 suggests the presence of a new gene involved in nonspecific X-linked mental retardation. Am J Med Genet A 118A, 217–222 (2003).
Thiselton, D. L. et al. An integrated, functionally annotated gene map of the DXS8026-ELK1 interval on human Xp11.3-Xp11.23: potential hotspot for neurogenetic disorders. Genomics 79, 560–572 (2002).
Neves, G., Cooke, S. F. & Bliss, T. V. Synaptic plasticity, memory and the hippocampus: a neural network approach to causality. Nat Rev Neurosci 9, 65–75 (2008).
Sakimura, K. et al. Reduced hippocampal LTP and spatial learning in mice lacking NMDA receptor epsilon 1 subunit. Nature 373, 151–155 (1995).
Brigman, J. L. et al. Loss of GluN2B-containing NMDA receptors in CA1 hippocampus and cortex impairs long-term depression, reduces dendritic spine density and disrupts learning. J Neurosci 30, 4590–4600 (2010).
Reisel, D. et al. Spatial memory dissociations in mice lacking GluR1. Nat Neurosci 5, 868–873 (2002).
Schmitt, W. B., Deacon, R. M., Seeburg, P. H., Rawlins, J. N. & Bannerman, D. M. A within-subjects, within-task demonstration of intact spatial reference memory and impaired spatial working memory in glutamate receptor-A-deficient mice. J Neurosci 23, 3953–3959 (2003).
Phillips, R. G. & LeDoux, J. E. Differential contribution of amygdala and hippocampus to cued and contextual fear conditioning. Behav Neurosci 106, 274–285 (1992).
Tayler, K. K., Tanaka, K. Z., Reijmers, L. G. & Wiltgen, B. J. Reactivation of neural ensembles during the retrieval of recent and remote memory. Curr Biol 23, 99–106 (2013).
Rodrigues, S. M., Schafe, G. E. & LeDoux, J. E. Intra-amygdala blockade of the NR2B subunit of the NMDA receptor disrupts the acquisition but not the expression of fear conditioning. J Neurosci 21, 6889–6896 (2001).
O'Roak, B. J. et al. Exome sequencing in sporadic autism spectrum disorders identifies severe de novo mutations. Nat Genet 43, 585–589 (2011).
Rehtanz, M., Schmidt, H. M., Warthorst, U. & Steger, G. Direct interaction between nucleosome assembly protein 1 and the papillomavirus E2 proteins involved in activation of transcription. Mol Cell Biol 24, 2153–2168 (2004).
Bedford, D. C., Kasper, L. H., Fukuyama, T. & Brindle, P. K. Target gene context influences the transcriptional requirement for the KAT3 family of CBP and p300 histone acetyltransferases. Epigenetics 5, 9–15 (2010).
Korzus, E., Rosenfeld, M. G. & Mayford, M. CBP histone acetyltransferase activity is a critical component of memory consolidation. Neuron 42, 961–972 (2004).
Alarcon, J. M. et al. Chromatin acetylation, memory and LTP are impaired in CBP+/− mice: a model for the cognitive deficit in Rubinstein-Taybi syndrome and its amelioration. Neuron 42, 947–959 (2004).
Petrij, F. et al. Rubinstein-Taybi syndrome caused by mutations in the transcriptional co-activator CBP. Nature 376, 348–351 (1995).
Wood, M. A., Attner, M. A., Oliveira, A. M., Brindle, P. K. & Abel, T. A transcription factor-binding domain of the coactivator CBP is essential for long-term memory and the expression of specific target genes. Learn Mem 13, 609–617 (2006).
Oliveira, A. M. et al. Subregion-specific p300 conditional knock-out mice exhibit long-term memory impairments. Learn Mem 18, 161–169 (2011).
Malovannaya, A. et al. Analysis of the human endogenous coregulator complexome. Cell 145, 787–799 (2011).
Silva, A. J. et al. Mutant mice and neuroscience: recommendations concerning genetic background. Banbury Conference on genetic background in mice. Neuron 19, 755–759 (1997).
Bucan, M. & Abel, T. The mouse: genetics meets behaviour. Nat Rev Genet 3, 114–123 (2002).
Clapcote, S. J. & Roder, J. C. Deletion polymorphism of Disc1 is common to all 129 mouse substrains: implications for gene-targeting studies of brain function. Genetics 173, 2407–2410 (2006).
Ishizuka, K. et al. Evidence that many of the DISC1 isoforms in C57BL/6J mice are also expressed in 129S6/SvEv mice. Mol Psychiatry 12, 897–899 (2007).
Koike, H., Arguello, P. A., Kvajo, M., Karayiorgou, M. & Gogos, J. A. Disc1 is mutated in the 129S6/SvEv strain and modulates working memory in mice. Proc Natl Acad Sci U S A 103, 3693–3697 (2006).
Strunk, K. E., Amann, V. & Threadgill, D. W. Phenotypic variation resulting from a deficiency of epidermal growth factor receptor in mice is caused by extensive genetic heterogeneity that can be genetically and molecularly partitioned. Genetics 167, 1821–1832 (2004).
Sun, G. et al. Isolation of differentially expressed genes in human heart tissues. Biochim Biophys Acta 1588, 241–246 (2002).
Liang, Y., Chiu, P. H., Yip, K. Y. & Chan, S. Y. Subcellular localization of SUN2 is regulated by lamin A and Rab5. PLoS One 6, e20507 (2011).
Desai, A., Turetsky, D., Vasudevan, K. & Buonanno, A. Analysis of transcriptional regulatory sequences of the N-methyl-D-aspartate receptor 2A subunit gene in cultured cortical neurons and transgenic mice. J Biol Chem 277, 46374–46384 (2002).
Grayson, D. R. et al. Reelin promoter hypermethylation in schizophrenia. Proc Natl Acad Sci U S A 102, 9341–9346 (2005).
Fu, M. et al. Cyclin D1 represses p300 transactivation through a cyclin-dependent kinase-independent mechanism. J Biol Chem 280, 29728–29742 (2005).
Im, H. et al. Measurement of Protein-DNA Interactions In Vivo by Chromatin Immunoprecipitation. in Signal Transduction Protocols Vol. 284, (eds. Dickson, R. & Mendenhall, M.) 129–146 (Humana Press, 2004).
Xie, H. et al. Brain-derived neurotrophic factor rescues and prevents chronic intermittent hypoxia-induced impairment of hippocampal long-term synaptic plasticity. Neurobiol Dis 40, 155–162 (2010).
Zhou, I. Y., Ding, A. Y., Li, Q., McAlonan, G. M. & Wu, E. X. Magnetic resonance spectroscopy reveals N-acetylaspartate reduction in hippocampus and cingulate cortex after fear conditioning. Psychiatry Res 204, 178–183 (2012).
Acknowledgements
This work was supported by HKU and donation to ‘Development and Disease’ programme. L Hang is supported by Edward Sai Kim Hotung Paediatric Education and Research Fund. We thank the University of Hong Kong Li Ka Shing Faculty of Medicine Core Facility for support on confocal microscopy and Shun Tak District Min Yuen Tong of Hong Kong for donating the Fast Real-time PCR system. We are grateful to Drs. Neil D Perkins (Institute for Cell and Molecular Biosciences, Newcastle University, UK), Andres Buonanno (National Institute of Child Health & Human Development, USA) and Dennis R. Grayson (University of Illinois at Chicago, USA) for gifts of reagents; Kimmy F.L. Tsang (HKU) for technical support; Dr. Soojka Chung (HKU) for the Morris water maze setup. We also thank Profs. Robin Lovell-Badge (MRC National Institute for Medical Research, UK) and Chi Chung Hui (University of Toronto, Canada) for suggestions on the manuscript.
Author information
Authors and Affiliations
Contributions
S.Y.C., W.H.Y., G.M., Y.S.C. designed the experiments and provided essential reagents and materials, K.H.T., S.K.L., Q.L., H.L., P.H.S.M., C.C.P.N. prepared samples, performed the experiments and prepared figures, S.Y.C., K.H.T., W.H.Y., G.M. wrote the main manuscript text. All authors reviewed the manuscript.
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Electronic supplementary material
Supplementary Information
Supplementary Figure S1. Absence of TSPYL2 protein in Tspyl2 mutant mice.
Rights and permissions
This work is licensed under a Creative Commons Attribution-NonCommercial-ShareALike 3.0 Unported License. To view a copy of this license, visit http://creativecommons.org/licenses/by-nc-sa/3.0/
About this article
Cite this article
Tsang, K., Lai, S., Li, Q. et al. The Nucleosome Assembly Protein TSPYL2 Regulates the Expression of NMDA Receptor Subunits GluN2A and GluN2B. Sci Rep 4, 3654 (2014). https://doi.org/10.1038/srep03654
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/srep03654
This article is cited by
-
CRL2APPBP2-mediated TSPYL2 degradation counteracts human mesenchymal stem cell senescence
Science China Life Sciences (2024)
-
TSPYL2 Regulates the Expression of EZH2 Target Genes in Neurons
Molecular Neurobiology (2019)
-
Xp11.2 microduplications including IQSEC2, TSPYL2 and KDM5C genes in patients with neurodevelopmental disorders
European Journal of Human Genetics (2016)
-
Tspyl2 Loss-of-Function Causes Neurodevelopmental Brain and Behavior Abnormalities in Mice
Behavior Genetics (2016)
-
Hydrogen Sulfide Prevents Synaptic Plasticity from VD-Induced Damage via Akt/GSK-3β Pathway and Notch Signaling Pathway in Rats
Molecular Neurobiology (2016)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
