
Abstract
Some small animals only use water transport mechanisms passively driven by surface energies. However, little is known about passive water transport mechanisms because it is difficult to measure the wettability of microstructures in small areas and determine the chemistry of biological surfaces. Herein, we developed to directly analyse the structural effects of wettability of chemically modified biological surfaces by using a nanoliter volume water droplet and a hi-speed video system. The wharf roach Ligia exotica transports water only by using open capillaries in its legs containing hair- and paddle-like microstructures. The structural effects of legs chemically modified with a self-assembled monolayer were analysed, so that the wharf roach has a smart water transport system passively driven by differences of wettability between the microstructures. We anticipate that this passive water transport mechanism may inspire novel biomimetic fluid manipulations with or without a gravitational field.
Similar content being viewed by others
Introduction
Water is important for maintaining life and activity. Land animals have evolved diverse water transport mechanisms depending on the habitat and body structure of animals that are actively driven by mechanical energies or passively driven by surface energies, including the extraction of moisture from food1, absorption through the skin2 or by oral/anal intake in most cases3,4,5. Most animals use suction or drinking, which requires mechanical energy to draw water into their mouths. Smart water transport systems based on a combination of active and passive mechanisms have been reported. Adult cats acquire water by lapping, thus exploiting the inertial energy of ascending columns based on water adhesion to the dorsal side of its tongue3. Shorebirds transport water using a capillary ratchet induced by repetitive opening and closing of their beak4.
On the other hand some small animals only use passive water transport mechanisms. Insects found in the Namib Desert collect water droplets from the morning fog using hydrophilic–hydrophobic microstructures on their backs before transportation to their mouth6. Australian lizards transport water to their mouth using the capillary system of their grooved skin7,8. Desert sandgrouses transport water to their young by wetting their feathers, which are densely covered with wettable hydrophilic fibres9. The wharf roach transports water through the open capillaries of the caudal legs10. In comparison to active transport, little is known about passive transport because it is difficult to measure the wettability of microstructures in small areas and characterize the chemistry of biological surfaces.
In this article, we determined how the wharf roach transports water via open capillaries on its legs. In general, water moves up an open capillary using the interfacial free energy resulting from the surface chemical composition and microstructures. Firstly we investigated the surface chemical composition of the wharf roach using conventional surface chemical analysis such as energy dispersive X-ray, infrared and Raman spectroscopies. However, elucidation of surface chemistry was difficult because of considerable individual variability and heat damage. Secondly, we tried to prepare duplicated open capillaries by template and photolithographic methods. However, exact duplicates of the legs could not be obtained because of the complicated microstructures. Herein, to solve these big problems of investigation of smart water transport systems of the wharf roach possessing complicated biological microstructures and undefined surface chemistry, we developed a novel method including direct modifications of biological microstructures and wettability analysis of small areas.
Results
Open capillaries on the legs of wharf roach
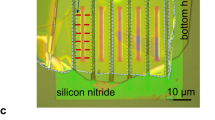
The wharf roach Ligia exotica (Crustacea, Isopoda, Fig. 1a) is a fast-moving arthropod found on rocky seashores with seven pairs of legs used to escape the surging surf and drowning. However, water is essential for its existence11, especially for gaseous exchange through its gills during respiration12. It possesses a smart water transport system10 that transports water from the wet surface of the seashore using a pair of caudal VI and VII legs (Fig. 1b). Vertical water transport was observed on the wet surface of the VI leg, from the lower end of the 2nd podite to the upper end of the 5th podite (Fig. 1c and see Supplementary Movie S1 online). Scanning electron microscope (SEM) observations revealed that open capillaries are present from the 2nd to the 5th podite, except the 6th podite on the VI leg (Fig. 1d). On the VII leg, open capillaries are found only on the 6th podite. Water is transported to the gills via these open capillaries when VI and VII legs are firmly apposed10.

Open capillaries on the legs of Ligia exotica.
(a) An digital photograph of Ligia exotica used in this study taken by Dr. Horiguchi. (b) Illustrations of Ligia exotica depicted by Dr. Horiguchi; side and ventral views. (c) Sequence of images showing water transport through open capillaries on the 5th podite of a freshly isolated VI leg. (d) Combined SEM image of the VI leg comprising the six podites: the dactylopodite (1st podite), the propodite (2nd podite), the carpopodite (3rd podite), the mieropodite (4th podite), the ischiopodite (5th podite) and the basipodite (6th podite). (e–j) SEM images of open capillaries; (e) the two different cuticular protrusions were observed in the open capillaries of the 4th podite, (f) a hair-like protrusion (HLP) located at the edge of the jointand (g) a paddle-like protrusion (PLP) located at the centre. (h) The outsides of the open capillary were covered with small curved plates. (i,j) The entire area of on the 3rd podite was covered with HLP. The outsides of open capillaries were also covered with small curved plates.
Detailed observations showed that open capillaries of the VI leg comprised two different cuticular protrusions, a hair-like protrusion (HLP) and a paddle-like protrusion (PLP). In the 4th and 5th podites, HLPs and PLPs are located at the edge and centre of open capillaries, respectively (Fig. 1e). Joints of legs contained HLPs (Fig. 1f). In the 2nd and 3rd podites, the entire area of open capillaries comprised only HLPs bending slightly towards the direction of oncoming water (Fig. 1i) and each protrusion is approximately 50-μm long (Fig. 1j). PLPs were approximately 100-μm high and 200-nm thick and were aligned parallel to the water flow (Fig. 1g). The outsides of open capillaries were covered with small curved plates (ca. 10-μm high; Fig. 1h), so that the outside roughness was lower than that of open capillaries.
Surface chemical modification of open capillaries
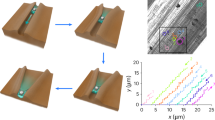
Legs of L. exotica were cut, rinsed and freeze dried13 to maintain fine structures, which were physically modified by gold sputtering (Fig. 2a). The interfacial free energies of the gold-sputtered legs were chemically modified using hydrophilic and hydrophobic compounds14,15. The same modification was performed on flat mica substrates to verify the relationship between the molar ratio and wettability. To quantify the narrow area in open capillaries (Fig. 2b), water contact angles (θ) were measured using 1.0 nanolitter droplet (Fig. 2c). We plotted θ of the chemically modified flat mica substrates (θf), HLP (θh) and PLP (θp) areas and the outside of open capillaries (θo) against the molar ratio of the hydrophobic thiol compound (r) (Fig. 2d and see Supplementary Movie S2–S4 online). θf decreased gradually from 108° (hydrophobic) to 13° (hydrophilic) as r decreased. θo also decreased gradually from 124° to 48° like θf, but it was slightly higher than θf (r ≥ 0.15). θh and θp decreased abruptly to 0° (superhydrophilic) at r = 0.50 and 0.35, respectively. According to the Cassie–Baxter wetting theory16, a rough surface containing small air pockets is more hydrophobic than a flat surface. On the hydrophobic-modified leg (r = 1.0), θ followed the order θp > θh > θo > θf. SEM images (Fig. 1e) clearly show that the roughness and amount of air pockets in PLP and HLP areas were greater than those on the outside. According to the Wenzel wetting theory17, when θf is smaller than 90°, a rough surface is more wettable than a flat surface. θh drastically decreased at r ≤ 0.6 (θf ≤ 90°). However, high hydrophobicity in the PLP area was maintained even when θf reached 72° (r = 0.40; Cassie–Baxter wetting). This wetting type was defined by roughness and the amount of air pockets, which depended on the shape of the surface microstructures in open capillaries.

Surface chemical modification of open capillaries.
(a) Schematic illustrations of surface physical/chemical modifications of the VI leg. Cut legs were physically modified by gold sputtering. The gold-sputtered legs were chemically modified using by hydrophilic and hydrophobic compounds. (b) Top view of measurement area of wettability analysis. (c) Projection view of the small volume droplet by a 1,000 fps high-speed camera in wettability analysis. (d) Water contact angles of the chemical-modified flat mica substrates (θf, yellow), the HLP area (θh, red) the PLP area (θp, blue) and the outside of open capillaries (θo, black) of the 4th podite plotted against the molar ratio of the hydrophobic thiol compound (r). Solid arrows indicate the samples shown in water transport analysis (Fig. 3).
The wettability-modified legs were classified into the following four regions according to the relationship between r (i.e. θf) and θ of the three microstructures (Fig. 2d). Region 1 indicated that all areas were modified to become superhydrophilic (θh = θp = θo = 0°). In region 2, open capillaries (θh = θp = 0°) were superhydrophilic, whereas the outside was not wettable. In region 3, only the HLP area of open capillaries was wettable (θh = 0°). Strong water repellency of all areas was observed in region 4.
Water transport through open capillaries of the wettability-modified legs
Water transport from the 4th to 5th podite of the wettability-modified legs was observed using green coloured water to make a colour contrast with the gold background. Water accumulated in region 1 of the wettability-modified legs in open capillaries and spread to the outer area of each podite. However, water transport was not observed in region 4 because of its strong water repellency. Two types of water flow through open capillaries were observed in region 2, (i) water leaked from the joint between each podite and spread to the outside (Fig. 3a and see Supplementary Movie S5 online; r = 0.30, θf = 62°) and (ii) water flowed only through open capillaries (Fig. 3b and see Supplementary Movie S6 online; r = 0.35, θf = 71°). Water leaked readily to the outside when θo was low in region 2. However, water transport via open capillaries was only achieved efficiently when θo was high. In region 3, water was not transported to the 5th podite (Fig. 3c and see Supplementary Movie S7 online; r = 0.4, θf = 78°). These results indicate that water only climbed upward via open capillaries when they were superhydrophilic compared with the outside (θp = 0° and θp ≪ θo). Hydrophobicity of the outside had an important role in preventing water leakage from open capillaries.

Water transport by open capillaries of the wettability-modified legs.
(a–c) Image sequence of water transport in open capillaries of the wettability-modified legs at (a) r = 0.30, (b) r = 0.35 and (c) r = 0.40.
Discussion
In the 2nd and 3rd podites, water flowed homogeneously along HLP, while in the 4th and 5th podites, water flowed along the HLP area initially, before filling the PLP area (Fig. 1c). To evaluate the wetting dynamics of open capillaries, penetration times of a 1-nL water droplet in HLP (th) and PLP (tp) areas were measured using a high-speed camera (Fig. 4a, 4b). th and tp of the wettability-modified legs at r = 0.35 were 17 ms and 66 ms, respectively. In region 2, th was always smaller than tp, suggesting that the HLP area was more wettable than the PLP area. Water flow in the 4th and 5th podites comprised two steps (Fig. 4c), an initial slow infiltration into the HLP area and its spread into the PLP area in the centre of open capillaries without any leakage. The features of water flow in the wettability-modified leg at r = 0.35 (θf = 71°) resembled those of freshly autotomized legs (Fig. 1c) whose θh (th), θp (tp) and θo were 0° (20 ms), 0° (150 ms) and 68 ± 19°, respectively. The PLP area in the centre of open capillaries on the 4th and 5th podites played an important role as a water reservoir. The results showed that the water roach adsorbed water using a passive mechanism produced by three different wettability areas comprising surface microstructures, i.e. the HLP, PLP and outside.

Mechanism of water transport by open capillaries.
(a,b) Sequence of images showing the penetration of a 1-nL water droplet in (a) HLP and (b) PLP areas of the wettability-modified legs at r = 0.35 captured using a high-speed camera (1,000 frames per second). (c) Schematic illustration of the water transport mechanism by open capillaries on the 4th and 5th podites. Water initially rapidly infiltrated into the HLP area, before spreading to the PLP area in the centre of open capillaries without any leakage. The PLP area in the centre of open capillaries played an important role as a water reservoir.
In this study, we determined the mechanism of a water transport system through open capillaries against gravity on the legs of an arthropod using a novel method that was readily applicable to biological surfaces. We conclude that the wharf roach transports water from a wet surface to the gills using a combination of two different micro-protrusions and the outside of open capillaries. The method of wettability modification of biological surfaces used in this study is applicable to other small creatures with passive water transport mechanisms. The results suggest that water can be transported through an open-capillary by means of differences in the interfacial free energy of structured surfaces even in a non-gravitational field such as cosmic space. Recently, an open air capillary system for use in a non-gravitational field was being researched at NASA's Ames Research Center18. We anticipate that this mechanism may be applied to open-air micro-fluidic devices and fluid manipulations with or without a gravitational field.
Methods
Animals
Adult specimens of the wharf roach Ligia exotica were collected from the boulders of the littoral of Lake Hamanako (34°45_N, 137°35_E), a brackish lake in Shizuoka Prefecture, on the Pacific Ocean side of Japan. The animals were maintained in the laboratory in plastic tanks containing shallow amounts of filtered seawater and a broken piece of plant pots.
Preparation of legs for scanning electron microscopy and gold spattering
The body length of ca. 2 cm was used for this experiment. The legs of animals were easily autotomized by controlling their free movements with the help of a pair of fine forceps at the proximal end of basipodite. The 6th and 7th legs were pre-fixed overnight in 2% glutaraldehyde and 2% paraformaldehyde buffered with 0.1 M sodium cacodylate buffer adjusted to pH 7.2. The specimens were then rinsed several times in 0.1 M sodium cacodylate buffer solution, post-fixed for 2 h in 1% OsO4 in phosphate buffered saline (0.13 M, pH 7.4) at room temperature, dehydrated through a graded series of ethanol solutions and rinsed in 100% t-butylalcohol three times at 37°C. Then the specimens were freeze-dried (JEOL, JFD-300) for several hours at 2°C.
For scanning microscopy observations, dried specimens were coated with OsO4 (Meiwa, Plasma multicoater PMC-5000) and observed with a scanning electron microscope (Hitachi S-3000N). For water flow observations, legs were coated with gold ion sputtering (5 mV, 225 s), in order to make them covered by same surface material. Various surface-modified legs with systematically changed interfacial free energies were made by surface treatment of hydrophilic and/or hydrophobic thiol compounds on gold-coated legs. The gold-coated legs were immersed in 1 mM ethanol solution including different molar ratios of hydrophilic thiol compound (11-mercapt-1-undecanol) and hydrophobic one (1-dodecane thiol) for 12 h. After the surface treatments, the chemical-modified legs were rinsed by ethanol and dried naturally for at least overnight. The same treatments were achieved by flat mica substrates to measure the surface wettability which is related with the interfacial free energy between the surface and water.
Observation of water flow using isolated legs
The law and wettability-modified legs were then attached to a fine plastic rod by dental wax to the autotomized proximal end. The other side of the rod was held in a manipulator (Narishige), so that the pereiopod could be stationed in its natural vertical position. The pereiopod was then immersed from the tip. The water was coloured with red/green water-soluble food dye (Tsukemoto Corporation) to facilitate observation of its flow with the help of a horizontally mounted stereomicroscope. To keep the position of pereiopod, the level of water was moved upward by a manipulator. The action was recorded on a digital camera (Olympus E-5, with video capture function).
Measurement of the interfacial free energy
The interfacial free energy was compared with surface wettability measured by the automatic microscopic contact angle meter (Kyowa MCA-3). A 1-nL microdroplet of water formed on the tip of a glass capillary having inner diameter of 5 μm was dropped on the surface with capturing of a projection view of the small volume droplet by a 1,000 fps high-speed camera. Water contact angles (θ) were measured by the captured image when the water microdroplet did not change for 10 ms. If the θ was 0°, in other wards the wettability of the surface was superhydrophilic, penetration times (t) of a 1-nL water droplet were measured using a high-speed camera.
References
Attenborough, D. [The Baking Deserts]. The Living Planet: A Portrait of the Earth [148] (Collins and British Broadcasting Corporation, London, 1984).
Bentley, P. J. & Yorio, T. Do frogs drink? J. Exp. Biol. 79, 41–46 (1970).
Reis, P. M., Jung, S., Aristoff, J. M. & Stocker, R. How cats lap: water uptake by Felis catus. Science 330, 1231–1234 (2010).
Prakash, M., Quéré, D. & Bush, J. W. M. Surface tension transport of prey by feeding shorebirds: the capillary ratchet. Science 320, 931–934 (2008).
Fox, H. M. Anal and oral intake of water by crustacean. J. Exp. Biol. 29, 583–599 (1952).
Parker, A. R. & Lawrence, C. R. Water capture by a desert beetle. Nature 414, 33–34 (2001).
Bentley, P. J. & Blumer, W. F. C. Uptake of water by the lizard, Moloch horridus. Nature 194, 699–670 (1962).
Gans, C., Merlin, R. & Blumer, W. F. C. The water-collecting mechanism of Moloch horridus re-examined. Amphibia-Reptilia 3, 57–64 (1982).
Cade, T. J. & Maclean, G. L. Transport of water by adult sandgrouse to their young. The Condor 69, 323–343 (1967).
Horiguchi, H., Hironaka, M., Meyer-Rochow, V. B. & Hariyama, T. Water uptake via two pairs of specialized legs in Ligia exotica (Crustacea, Isopoda). Biol. Bul. 213, 196–203 (2007).
Edney, E. B. The temperature of woodlice in the sun. J. Exp. Biol. 30, 331–349 (1953).
Edney, E. B. [Water Loss–Respiratory]. Water Balance in Land Arthropods [66–86] (Springer-Verlag, Berlin, 1977).
Osatake, H. & Inoue, T. Drying method of biological specimens for scanning method microscopy: the t-butyl alcohol. Arch. Histol. Cytol. 51, 53–59 (1988).
Porter, M. D., Bright, T. B., Allara, D. L. & Chidsey, C. E. D. Spontaneously organized molecular assemblies. 4. Structural characterization of n-alkyl thiol monolayers on gold by optical ellipsometry, infrared spectroscopy and electrochemistry. J. Am. Chem. Soc. 109, 3559–3568 (1987).
Ulman, A., Evans, S. D., Shnidman, Y., Sharma, R., Eilers, J. E. & Chang, J. C. A. Concentration-driven surface transition in the wetting of mixed alkanethiol monolayers on gold. J. Am. Chem. Soc. 113, 1499–1506 (1991).
Cassie, A. B. D. & Baxter, S. Wettability of porous surfaces. Trans. Faraday Soc. 40, 546–551 (1944).
Wenzel, R. N. Resistance of solid surfaces to wetting by water. Ind. Eng. Chem. 28, 988–994 (1936).
Thomas, E. A., Weislogel, M. M. & Klaus, D. M. Design considerations for sustainable spacecraft water management systems. Adv. Space Res. 46, 761–767 (2010).
Acknowledgements
This research was partly supported by Grant-in-Aid for Young Scientists (A) (No. 24686076) of The Japan Society for the Promotion of Science (JSPS) and Grant-in-Aid for Scientific Research on Innovative Areas “Engineering Neo-Biomimetics” (No. 24120004) of The Ministry of Education, Culture, Sports, Science and Technology (MEXT), Japan.
Author information
Authors and Affiliations
Contributions
H.H. collected animals and prepared dried samples; Y.H. performed surface modifications; D.I. collected SEM images and data and performed wettability analysis; Y.M., H.Y. and K.I. performed conventional surface chemical analyses; K.T. and T.S. discussed wettability; D.I., T.H. and M.S. designed the study and wrote the paper. All authors discussed the results and commented on the manuscript.
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Electronic supplementary material
Supplementary Information
Supplementary Information
Supplementary Information
Movie S1
Supplementary Information
Movie S2
Supplementary Information
Movie S3
Supplementary Information
Movie S4
Supplementary Information
Movie S5
Supplementary Information
Movie S6
Supplementary Information
Movie S7
Rights and permissions
This work is licensed under a Creative Commons Attribution-NonCommercial-NoDerivs 3.0 Unported License. To view a copy of this license, visit http://creativecommons.org/licenses/by-nc-nd/3.0/
About this article
Cite this article
Ishii, D., Horiguchi, H., Hirai, Y. et al. Water transport mechanism through open capillaries analyzed by direct surface modifications on biological surfaces. Sci Rep 3, 3024 (2013). https://doi.org/10.1038/srep03024
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/srep03024
This article is cited by
-
Route of pesticide spread on the body surface of Blattella germanica (Linnaeus): a NanoSuit–energy dispersive X-ray spectroscopy analysis
Scientific Reports (2023)
-
A comprehensive evaluation of the potential of semiterrestrial isopods, Ligia exotica, as a new animal food
Scientific Reports (2021)
-
Hydrophobic-hydrophilic crown-like structure enables aquatic insects to reside effectively beneath the water surface
Communications Biology (2021)
-
Hierarchical structures hydrogel evaporator and superhydrophilic water collect device for efficient solar steam evaporation
Nano Research (2021)
-
Bioinspired Unidirectional Liquid Transport Micro-nano Structures: A Review
Journal of Bionic Engineering (2021)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
