
Abstract
Expression of chitinase is developmentally regulated in insects in consonance with their molting process. During the larval-larval metamorphosis in Helicoverpa armigera, chitinase gene expression varies from high to negligible. In the five-day metamorphic course of fifth-instar larvae, chitinase transcript is least abundant on third day and maximal on fifth day. MicroRNA library prepared from these highest and lowest chitinase-expressing larval stages resulted in isolation of several miRNAs. In silico analysis of sequenced miRNAs revealed three miRNAs having sequence similarity to 3′UTR of chitinase. Gene-targeted specific action of these miRNAs, was investigated by luciferase reporter having 3′UTR of chitinase. Only one of three miRNAs, miR-24, inhibited luciferase expression. Further, a day-wise in vivo quantification of miR-24 in fifth-instar larvae revealed a negative correlation with corresponding chitinase transcript abundance. The force-feeding of synthetic miR-24 induced significant morphological aberrations accompanied with arrest of molting. These miR-24 force-fed larvae revealed significantly reduced chitinase transcript abundance.
Similar content being viewed by others


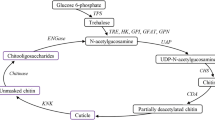
Introduction
MicroRNAs (miRNAs) are small non-coding endogenous RNAs (ca. 22 nucleotides) derived from much longer pre-primary transcripts1. The miRNAs represent a major class of post-transcriptional regulatory molecules, which play a key role in a wide diversity of biological processes such as development, metabolism and apoptosis in eukaryotic organisms2,3,4. They control the expression of target genes through binding to complementary target sites in mRNA targets5,6,1. In animals, these sites typically have imperfect complementarity to the miRNA. The ability to target imperfectly complementary target sites, along with other miRNAs in the genome, has led to the prediction of thousands of genes which are miRNA regulated7,8. However, despite the apparent abundance of potential miRNA/3′UTR target interactions, only few animal miRNA/target pairs have been validated till date in a physiologically relevant context9,10.
An array of diverse miRNAs are constitutively expressed whereas a few others show temporal and tissue-specific expression patterns11. A number of miRNAs have been isolated and annotated from several eukaryotic organisms including insects. Nearly 21,264 miRNAs were deposited in the latest miRBase database release 19.0 in August 2012. Nearly 3457 insect miRNAs have been identified and these mainly restrict to Diptera (Drosophila sp., Anopheles gambiae, Aedes aegypti, Culex quinquefasciatus), Hymenoptera (Nasonia sp., Apis mellifera), Coleoptera (Tribolium castaneum), Orthoptera (Locusta migratoria), Lepidoptera (Bombyx mori, Manduca sexta, Spodoptera litura, Helicoverpa armigera, Heliothis virescens) and Homoptera (Acyrthosiphon pisum) (http://www.mirbase.org/). However, the targets of most of these annotated miRNAs have not been identified yet.
In S. litura, 58 putative miRNAs have been identified using computational methods. The temporal and spatial expression profiles of these miRNAs have been examined in different tissues and stages of development12. By sequencing small RNA libraries of M. sexta embryos, larvae, pupae and adults, 163 conserved and 13 novel miRNAs have been identified so far13. Homology-based predictions of lepidopteran miRNAs have been conducted in H. armigera and H. virescens14,15. Nevertheless, a few miRNAs that control metamorphic transition from larvae to pupae have been functionally identified in B. mori16.
Extensive softwares have been developed to tentatively identify the targets of miRNAs but the identification of precise biological function targeted by miRNAs still remains a challenge. It has been estimated that the false positive rate of in silico predicted miRNAs is 24–70%17. This inability to predict and correlate miRNA controlled metabolic events, emphasizes the need for experimental validation of the predicted miRNAs, in order to identify their specific targets. Several different approaches are being pursued to identify miRNA targets and have been recently described18.
We have been working on the profiling of chitinase (EC 3.2.1.14) at various larval transition stages in Helicoverpa armigera19. In insects, chitin forms a major component of the cuticle and the peritrophic membrane of the midgut20. Chitinase catalyzes the degradation of chitin, a linear homopolymer of N-acetylglucosamine in a β-(1,4) linkage and plays a major role in insect metamorphosis21. The expression of chitinase coincides with the molting of larvae and is regulated by ecdysone19,21.
To profile relative expression abundance of various microRNAs at various stages of chitinase expression, an miRNA library was prepared from different larval stages. These microRNAs were enriched by their ability to hybridize with 3′ untranslated region (UTR) of chitinase gene. Using solution hybridization, we identified a few putative miRNAs targeting the chitinase gene in H. armigera (hachi). The correlation between putative microRNAs interacting with 3′UTR of hachi was examined by cloning it downstream of 3′end of firefly luciferase reporter gene (Fluc). The direct involvement of miR-24 in the molting process was examined by force-feeding synthetic miR-24 to H. armigera larvae. The in vivo levels of this miRNA at various stages of larval development corresponding to chitinase expression were quantified. These force-fed larvae showed arrested molting process with corresponding reduction of chitinase transcript abundance.
Results
Temporal expression of H. armigera chitinase (hachi)
The molting stage of 5th instar larvae of H. armigera is spread over 5 days followed by pupation. The relative levels of hachi transcript in different days of 5th instar larvae were analyzed by Real-time qPCR (Figure 1).

Transcript abundance of H. armigera chitinase (hachi).
The relative abundance of hachi transcript in 5th instar larvae of H. armigera (1–5 day old) was determined by Real-time qPCR. Total RNA isolated from the midgut tissue was used as template and normalised to internal control, β-actin. Expression levels of hachi were measured using Comparative CT method. A non-template control was run with every experiment. Representative data (mean ±S.D.) from three independent experiments is shown.
A varying pattern of hachi expression was observed in 1–5 day old, 5th instar larvae (Figure 1). High chitinase level is required for dissolution of exoskeleton and permits expansion of larvae mass. Chitinase transcript was highly abundant on the first and fifth day of 5th instar i.e. immediately upon molting and close to pre-pupa stage. However, detectable hachi transcript was present during intermolting stages on second, third and fourth days. Nearly, 2-folds increase in hachi transcript was observed on fifth day (last day) as compared to fourth day (Figure 1). The observed pattern of chitinase expression coincides with reported levels of chitinase enzyme19. Such pattern reflects the developmental necessity of chitinase for insect to molt.
Isolation and functional validation of chitinase-specific miRNAs
Chitinase-specific miRNAs were isolated from H. armigera midgut tissue using Solution hybridization technique. The hybridized small RNAs were cloned into pGEM-Te vector and the recombinant colonies were initially screened using vector-specific primers. Approximately 50 insert positive colonies were sequenced and out of these 50, three colonies revealed sequences that displayed varying degree of complementarity to 3′UTR of chitinase. The three putative miRNAs viz. miR-24, miR-2 and miR-131 were detected which displayed >65% sequence similarity to the 3′UTR of hachi (Figure 2; Table 1).

Nucleotide sequence of 3′UTR of H. armigera chitinase.
A 1057 bp fragment encoding the 3′UTR of hachi has been cloned in pGEM-Te vector and sequenced. The nucleotide sequence of 3′UTR corresponding to miR-2, miR-24 and miR-131 (miRBase) is shown. The bases in black are completely identical while those in red are mismatches.
The potential miRNA/3′UTR target interactions were studied by conducting luciferase assays. The 3′UTR of hachi was cloned downstream to 3′end of firefly luciferase gene (Fluc) in pIB/V5-His Topo vector to generate firefly luciferase reporter construct. Co-transfection of this luciferase reporter construct along with pIB-Rluc vector and synthetic miR-24 into Sf21 cells resulted in inhibition of luciferase activity (Figure 3). Dose-dependent reduction in luciferase activity was observed with nearly five-folds reduction in luminescence upon co-transfection with 200 nM of synthetic miR-24 as compared to controls. In contrast, other miRNAs that displayed sequence similarity to 3′UTR of hachi viz. miR-2 and miR-131 did not inhibit luciferase activity (Figure 3).

Functional validation of chitinase-specific miRNAs using luciferase-based reporter assay in Sf21 cells.
The 3′UTR of hachi was cloned downstream to firefly luciferase (Fluc) in pIB vector. Sf21 cells were co-transfected with recombinant pIB-Fluc-3′UTRhachi, pIB-RLuc control vector and miR-2 or miR-131 or miR-24 (10–250 nM). Dual-luciferase reporter assays were performed 48 hrs after transfection. Firefly/Renilla luciferase ratios were calculated. Histogram depicts the normalised luciferase activity from three independent experiments. The following controls were set up: Control 1: Only Sf21 cells; Control 2: (pIB): Sf21cells transfected with pIB vector alone; Control 3: (FLuc+RLuc): Sf21 cells were co-transfected with pIB-Fluc+pIB-Rluc. The control set of experiments were also repeated thrice in triplicates.
In vivo levels of miR-24 in H. armigera larvae
Having established the pattern of chitinase gene expression at different days during molting, we estimated levels of miR-24 at corresponding stages. The quantitative analysis of miR-24 in different days of 5th instar larvae of H. armigera was analyzed by Real-time qPCR (Figure 4). QuantiMir cDNA was prepared from midgut total RNA and normalised to β-actin which served as internal control. The levels of miR-24 increased steadily from day1, peaking at day 3 and reduced subsequently upto day 5. The transcript level of miR-24 was nearly 3-folds less on the first and fifth day of 5th instar as compared to third day. Nearly, 5-folds increase in miR-24 transcript was observed on the third day as compared to fifth day (Figure 4).

Quantitation of miR-24 in H. armigera larvae.
The relative abundance of miR-24 transcript was estimated in 5th instar larvae of H. armigera (1–5 day old) by Real-time qPCR. Total RNA was extracted from the midgut tissue and used as template. Transcripts were normalised to β-actin control. Expression levels of miR-24 were measured using Comparative CT method. A non-template control was run with every experiment. Representative data (mean ±S.D.) from three independent experiments is shown.
An overall inverse pattern of miR-24 expression was observed in relation to chitinase transcript levels. Thus, the observed inverse relationship of miR-24 and chitinase together with sequence similarity of miR-24 to 3′UTR of hachi suggested that miR-24 indeed is involved in the regulation of chitinase.
Phenotypic effect of synthetic miR-24 on H. armigera larvae
Comparative data on the relative abundance of miR-24 and chitinase clearly revealed inverse relationship suggesting that high miR-24 levels would block molting process. To investigate further, we administered synthetic miR-24 to the 4th instar two-day old larvae. Upon ingestion of synthetic miR-24 (0.02 nmoles), the larvae ceased to feed and could not molt further (Figure 5; Table 2). The relative expression level of hachi was monitored in both miR-24 fed and control larvae by Real-Time qPCR (Figure 6). Nearly, 2-folds reduction in hachi transcript was observed in miR-24 fed larvae after 24 hrs as compared to controls (Figure 6). After 48 hrs, the larvae did not molt, were shrunken and eventually died (Figure 5; Table 2).

Phenotypic effect of miR-24 in H. armigera larvae.
The H. armigera larvae (4th instar, two-day old) were force-fed with synthetic miR-24 (0.02 nmoles) using a microinjector. Larvae fed with Ringer's buffer and scrambled synthetic miR-24 served as controls. The larvae were monitored upto 48 hrs and morphological changes were observed.

Transcript abundance of hachi in miR-24 fed H. armigera larvae.
The H. armigera larvae (4th instar, two-day old) were force-fed with synthetic miR-24 (0.02 nmoles) and midgut tissue was dissected 24 hrs post-feeding. Total RNA was isolated from the midgut tissue of miR-24 fed (treated), scrambled miR-24 fed (non-specific control) and buffer-fed (control) larvae. Chitinase transcript was examined by Real-time qPCR using β-actin as internal control. Expression levels of hachi were measured using Comparative CT method. A non-template control was run with every experiment. Representative data (mean ±S.D.) from three independent experiments is shown.
Discussion
The temporally regulated expression of chitinase gene in 5th instar larvae of H. armigera focussed our investigation towards its regulation and prompted to explore the role of miRNAs. Instead of the standard protocol of in silico miRNA prediction followed by their experimental validation for target identification, a reverse in vitro analysis was carried out to identify the miRNAs targeting chitinase mRNA. The strategy employed in the present investigation facilitated the identification of microRNAs that displayed sequence similarity to 3′UTR of hachi. As in animal microRNA targets, the sequence similarity was imperfect that varied from 69–85%22.
A comparative analysis of all these tentatively identified miRNAs revealed varying degree of sequence similarity with 3′UTR of chitinase. The miR-2, miR-131 and miR-24 displayed 69, 89 and 75% sequence similarity respectively at different regions of 3′UTR. Of these, miR-24 displayed perfect match at 4-12 position and may be representing the seed region. The percentage sequence similarity did not correlate with the observed down regulation of chitinase indicating that mere sequence identity cannot be the prime criteria for target identification23.
Several studies have shown that only a few in silico predicted miRNAs eventually are validated by in vivo experimentation24,25,18. To identify the functional miRNAs, all three miRNAs, miR-2, miR-131 and miR-24 were examined further for their functional interaction with 3′UTR of hachi by employing dual luciferase assay system. The assay employed pIB/V5-His Topo vector with 3′UTR of hachi cloned after the 3′end of Fluc. This construct was used for assessing the effect of miRNAs individually. Co-transfection of firefly luciferase reporter plasmid with miR-24 showed inhibition of luciferase activity in a concentration-dependent manner whereas miR-2 and miR-131 did not show any effect on luciferase activity. The participation of miR-24 in regulating expression of chitinase was further substantiated by the observed low levels of miR-24 at developmental stage which displayed highest level of chitinase transcript. In the present study, miR-24 levels were highest at day 3 of 5th instar larva, at this stage chitinase transcript abundance was minimal. The inverse relationship in the temporal expression pattern of miR-24 and chitinase provides clear evidence of negative regulation of chitinase gene expression by miR-24. The microRNAs are known to mediate their effect through target miRNA degradation or through translational repression22. At present it is difficult to speculate on the mechanism of action of miR-24 on chitinase transcript abundance22.
A direct correlation of miR-24 regulation of chitinase expression was revealed in experiments where larvae were force-fed with miR-24. The miR-24 force-fed larvae were shrunk, could not moult and also showed drastic reduction in the chitinase transcript levels depicting morphological and physiological changes in vivo. This suggests that miR-24 acts as a genetic switch in the regulatory cascade of chitinase gene function22. Taken together, our results reveal that modulation of relative levels of miR-24 have a direct bearing on important morphogenetic developmental process. Also, most importantly our approach of identifying microRNAs that control diverse developmental events could be applied to other tightly regulated biochemical processes.
Methods
Insect rearing, midgut dissections and RNA isolation
The culture of H. armigera was maintained in our insectary under controlled conditions of temperature 25°C, 70% relative humidity and a photoperiod of 14 hrs light: 12 hrs dark. The larvae were reared on a semi-synthetic artificial diet. Fifth-instar larvae (1-5 day old) were chilled on ice for 15 min and dissected to isolate the midgut tissue. The midgut was slit longitudinally, the peritrophic membrane was removed and the residual midgut contents were rinsed away with diethyl pyrocarbonate (DEPC)-treated water. The midgut tissue was either used immediately or snap frozen in liquid nitrogen and then stored at −80°C until further use.
Total RNA was isolated from the midgut tissue using Trizol reagent (Invitrogen, USA) as directed by the manufacturer.
Real-Time qPCR analysis
The relative transcript abundance of chitinase was determined by Real-Time qPCR analysis using GENE-REAMix™ SYBR One Step kit (Puregene, Genetix Biotech, India) and StepOne™ Real-Time PCR System (Applied Biosystems, USA). Gene-specific primers were designed from cDNA sequence of H. armigera chitinase (NCBI accession no. AY325496). The chitinase transcript was amplified using primers (HelioN3 For: 5'-AGGAACTTCACAGCTCTTCG-3' and HelioC2 Rev: 5'-CTCATAAGCCCACTGATCATG-3'). The amount of total RNA was normalised to β-actin transcript using primers (β-actin For: 5'-CAGATCATGTTTGAGACCTTCAAC-3' and β-actin Rev: 5'-GA/C/TCCATCTCC/TTGCTCGAAA/GTC-3'). Each 25 μl reaction mixture contained 2 μl of template RNA (100–500 ng/μl), 2X Gene-REAMix™ SYBR One Step, 0.25 μl of forward and reverse primer (25 μM), 0.5 μl of RiboSafe RNase Inhibitor (10 U/μl) and RNase-free water. Real-time cycling conditions included a preliminary reverse transcription at 42°C for 10 min, an initial activation step at 95°C for 10 min followed by 40 cycles each of 95°C, 15 sec; 52°C, 15 sec and 72°C, 15 sec respectively. The final step included gradual temperature increase from 60°C to 95°C at the rate of 0.3°C/10 sec to enable melt-curve data collection. A non-template control (NTC) was run with every assay. Reactions were set up in triplicates.
The threshold cycles (CT) were recorded for each amplicon (hachi and β-actin) and the difference between the CT (i.e ΔCT) was determined. The relative abundance was calculated using Comparative CT method using the formula 2−ΔΔCT26.
Isolation and synthesis of chitinase-specific miRNAs
Isolation of small RNAs from midgut tissue
Total RNA isolation and small RNA enrichment procedure were performed using mirVana™ miRNA Isolation Kit (Ambion, USA) as directed by the manufacturer. Briefly, the midgut tissue was homogenized in lysis buffer. The lysate was then extracted with acid-phenol:chloroform, added ethanol to bring up the sample to 25% ethanol and fractionated to isolate the large and small RNAs using glass-fiber filter. RNA species (<200 nt) were visualized on a 15% denaturing polyacrylamide gel.
Cloning of 3′UTR of H. armigera chitinase
The full-length H. armigera chitinase (hachi) has been cloned and characterized in our laboratory using RACE technology19. Using plasmid DNA of 3′RACE product of hachi as template and gene-specific primers (HaUTR For: 5′-TGGTCATTGCACAGATACCGT-3′) and (HaUTR Rev: 5′-GGATCCACCATAATTTATTGTCTTA-3′), a 1057 bp fragment encoding 3′UTR of hachi was amplified. This fragment was cloned into the pGEM-Te vector (Promega, USA) and sequenced (Macrogen, Korea). The RNA transcript of the 3′UTR of hachi was synthesized by in vitro transcription using T7 RNA polymerase (MBI Fermentas, USA).
Solution Hybridization assay
The miRNA expression levels in small RNA samples were measured by solution hybridization using mirVana™ miRNA Detection Kit (Ambion, USA) as directed by the manufacturer. Briefly, small RNA species and RNA transcript of 3′UTR of hachi were hybridized in solution at 42°C for 12 hrs. Single-stranded RNAs were digested using RNase A/T1 solution and RNA-RNA hybrids were protected and precipitated. To clone the protected RNAs, an oligoribonucleotide was ligated using T4 RNA ligase (MBI Fermentas, USA) and cDNA was synthesized using SuperScript III (Invitrogen, USA). The cDNA was polyG-tailed at 3′end using terminal deoxynucleotidyl transferase (Invitrogen, USA). Using primers complementary to the adaptors, the cDNA was amplified and cloned into pGEM-Te vector and sequenced (Macrogen, Korea).
miRNA synthesis
Based on the sequence of the isolated miRNAs and their sequence identity to the 3′UTR of hachi, double-stranded miRNAs were synthesized commercially (Dharmacon, USA). These synthetic miRNAs were used for luciferase reporter assay and force-feeding experiments.
Cell culture, transfections and luciferase assay
Full-length Firefly luciferase (Fluc) and Renilla luciferase (Rluc) cDNAs were amplified and cloned in an insect expression vector, pIB/V5-His TOPO (Invitrogen, USA) and named ‘pIB-Fluc control vector’ and ‘pIB-Rluc control vector’ respectively. The 3′UTR of hachi was cloned downstream to pIB-Fluc after the 3′end of Fluc in EcoRV/SacII sites. This construct was referred as ‘recombinant firefly luciferase reporter vector’ or ‘pIB-Fluc-3′UTR hachi’.
For luciferase assays, Sf21 cells were grown and maintained at 27°C in BD Baculogold TNM-FH insect medium (BD Biosciences, USA) containing 10% fetal bovine serum and gentamicin. Cells were grown as a monolayer upto 70–80% confluence in T-75 flasks (Nunc, USA). Two hours before transfection, Sf21 cells (60–70% confluent) were scraped and approximately 4 × 105 cells/well were seeded into 6-well plates. Cells were co-transfected with pIB-Fluc plasmid (1 μg), pIB-Rluc plasmid (1 μg) and miR-2 or miR-131 or miR-24 at varying concentrations (10–200 nM) using cellfectin reagent (Invitrogen, USA) in a final volume of 0.6 ml. Four hours post-transfection, serum plus medium was added to the cells and the culture plate was held stationary at 27°C for 48 hrs.
After 48 hrs, Firefly and Renilla luciferase activity was measured sequentially using Dual Luciferase assay kit (Promega, USA) as directed by the manufacturer. Luminescence was read on a Packard LumiCount manual luminometer (Cole Parmer, USA). Firefly luciferase values were normalised to Renilla luciferase values and the luciferase levels were reported as ratio of FLuc/RLuc. This normalised luciferase activity was plotted. Each transfection was tested in three independent experiments, each performed in triplicates. Luciferase activity was measured thrice for each sample.
The following controls were set up. Only Sf21 cells served as first control while only Sf21cells transfected with pIB vector only served as second control. Both did not show luminescence. In third control, Sf21 cells were co-transfected with pIB-Fluc vector and pIB-Rluc vector without 3′ UTR of hachi. Luminescence from the test samples were normalized against the ratio of Fluc/Rluc obtained from the third control. Further, a parallel set of controls included Sf21 cells transfected individually with pIB-Fluc vector/pIB-Rluc vector (without 3′UTR of hachi) to optimize transfection regimen.
Quantitation of miR-24 in H. armigera larvae
The expression level of miR-24 in 5th instar larvae (1–5 day old) was estimated using QuantiMir RT kit Small RNA Quantitation system (SBI System Biosciences, USA). Briefly, cDNA pool of anchor-tailed miRNAs (QuantiMir cDNA) was prepared from midgut total RNA as per the manufacturer's guidelines. Real-Time qPCR was performed using Power SYBR Green PCR Master Mix (Applied Biosystems, USA) in StepOne™ Real-Time PCR System (Applied Biosystems, USA) and QuantiMir cDNA as template. The miR-24 transcript was amplified using primers (miR-24 For: 5′-TGAACTGAGGATCAACCCG-3′) and 3′ universal reverse primer (3′URP). The amount of total RNA was normalised to β-actin transcript using β-actin forward and reverse primers (mentioned earlier).
Each 30 μl reaction mixture contained 1 μl of diluted QuantiMir cDNA, 2X Power SYBR Green PCR Master Mix, 1 μl of miR-24 For (10 μM) and 0.5 μl of 3′URP (10 μM) and RNAse-free water. Cycling and data accumulation conditions included an initial heating step at 50°C for 2 min, an initial activation step at 95°C for 10 min followed by 40 cycles each of denaturation (94°C, 15 sec), annealing and extension (50°C, 30 sec). The final step included gradual temperature increase from 60°C to 95°C at the rate of 0.3°C/10 sec to enable melt-curve data collection. A non-template control (NTC) was run with every assay. The threshold cycles (CT) were recorded for each amplicon (miR-24 and β-actin) and the difference between the CT (i.e ΔCT) was determined. The relative abundance was calculated using Comparative CT method using the formula 2−ΔΔCT26.
Force-feeding of miR-24
H. armigera larvae (4th instar, two-day old) were force-fed with 0.02 nmoles of synthetic miR-24 (5′-UGAACUGAGGAUCAACCCG-3′) (Dharmacon, USA) using a microinjector (KPS210, KD scientific, USA). Control larvae were force-fed with Ringer's buffer while scrambled synthetic miR-24 (5′-UAGGCGACUAUAACCCGGA-3′) force-fed larvae served as non-specific control. Each experiment was repeated thrice with 20 larvae per treatment. The chitinase transcript level in midgut tissue of both control and treated larvae was quantified by Real-Time qPCR.
References
Bartel, D. P. MicroRNAs: genomics, biogenesis, mechanism and function. Cell 116, 281–297 (2004).
Ambros, V. The functions of animal microRNAs. Nature 431, 350–355 (2004).
Miska, E. A. How microRNAs control cell division, differentiation and death. Curr. Opin. Genet. Dev. 15, 563–568 (2005).
Wienholds, E. & Plasterk, R. H. MicroRNA function in animal development. FEBS Lett. 579, 5911–5922 (2005).
Lee, R. C., Feinbaum, R. L. & Ambros, V. The C. elegans heterochronic gene lin-4 encodes small RNAs with antisense complementarity to lin-14. Cell 75, 843–854 (1993).
Reinhart, B. J. et al. The 21-nucleotide let-7 RNA regulates developmental timing in Caenorhabditis elegans. Nature 403, 901–906 (2000).
Rajewsky, N. MicroRNA target predictions in animals. Nat. Genet. (Suppl) 38, S8–13 (2006).
Didiano, D. & Hobert, O. Molecular architecture of a miRNA-regulated 3′ UTR. RNA 14, 1297–1317 (2008).
Carthew, R. W. Gene regulation by microRNAs. Curr. Opin. Genet. Dev. 16, 203–208 (2006).
Stefani, G. & Slack, F. J. Small non-coding RNAs in animal development. Nat. Rev. Mol. Cell. Biol. 9, 219–230 (2008).
Carrington, J. C. & Ambros, V. Role of microRNAs in plant and animal development. Science 301, 336–338 (2003).
Rao, Z. et al. Identification, Expression and Target Gene Analyses of MicroRNAs in Spodoptera litura. PLoS ONE 7, e37730 (2012).
Zhang, X. et al. Identification and developmental profiling of conserved and novel microRNAs in Manduca sexta. Insect Biochem. Mol. Biol. 42, 381–95 (2012).
Ge, X. et al. Identification of MicroRNAs in Helicoverpa armigera and Spodoptera litura Based on Deep Sequencing and Homology Analysis. Int. J. Biol. Sci. 9, 1–15 (2013).
Chilana, P., Sharma, A., Arora, V., Bhati, J. & Rai, A. Computational identification and characterization of putative miRNAs in Heliothis virescens. Bioinformation 9, 79–83 (2013).
Yu, X. et al. The silk worm (Bombyx mori) microRNAs and their expression in multiple developmental stages. PLoS ONE 3, e2997 (2008).
Selbach, M. et al. Widespread changes in protein synthesis induced by microRNAs. Nature 455, 58–63 (2008).
Thomson, D. W., Bracken, C. P. & Goodall, G. J. Experimental strategies for microRNA target identification. Nucleic Acids Res. 39, 6845–53 (2011).
Ahmad, T., Rajagopal, R. & Bhatnagar, R. K. Molecular characterisation of chitinase from polyphagous pest Helicoverpa armigera. Biochem. Biophys. Res. Commun. 310, 188–195 (2003).
Kramer, K. J. & Koga, D. Insect chitin: physical state, synthesis, degradation and metabolic regulation. Insect Biochem. 16, 851–877 (1986).
Kramer, K. & Muthukrishnan, J. S. Insect chitinases: molecular biology and potential use as biopesticides. Insect Biochem. Mol. Biol. 27, 887–900 (1997).
Flynt, A. S. & Lai, E. C. Biological principles of microRNA-mediated regulation: shared themes amid diversity. Nat. Rev. Genet. 9, 831–842 (2008).
Lai, E. C. MicroRNAs are complementary to 3′ UTR sequence motifs that mediate negative post-transcriptional regulation. Nat. Genet. 30, 363–364 (2002).
Kuhn, D. E. et al. Experimental validation of miRNA targets. Methods 44, 47–54 (2008).
Orom, U. A. & Lund, A. H. Experimental identification of microRNA targets. Gene 451, 1–5 (2010).
Pfall, M. W. A new mathematical model for relative quantification in real time RT-PCR. Nucl. Acids Res. 29, 2002–2007 (2001).
Acknowledgements
The manuscript is dedicated to the memory of Dr. Neema Agrawal who initiated the work but did not live long enough to see completion of the manuscript. The work was supported from fund received from NFBSFARA (National Fund for Basic, Strategic and Frontier Application Research in Agriculture), ICAR (Indian Council for Agricultural Research), India [Grant no. RNAi-2012]. We gratefully acknowledge Dr. Sunil K. Mukherjee for critical discussion during the course of the investigation.
Author information
Authors and Affiliations
Contributions
N.A. and B.S. were involved in preparation and isolation of small microRNA. B.S. performed Real-Time qPCR. N.A. and J.R. performed luciferase assays. K.S.S. carried out midgut dissections. R.K.B. and B.S. wrote the manuscript. R.K.B. conceived this project and carried out data analysis interpretation.
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Rights and permissions
This work is licensed under a Creative Commons Attribution-NonCommercial-ShareALike 3.0 Unported License. To view a copy of this license, visit http://creativecommons.org/licenses/by-nc-sa/3.0/
About this article
Cite this article
Agrawal, N., Sachdev, B., Rodrigues, J. et al. Development associated profiling of chitinase and microRNA of Helicoverpa armigera identified chitinase repressive microRNA. Sci Rep 3, 2292 (2013). https://doi.org/10.1038/srep02292
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/srep02292
This article is cited by
-
RNA interference and crop protection against biotic stresses
Physiology and Molecular Biology of Plants (2021)
-
Identification of microRNAs and their response to the stress of plant allelochemicals in Aphis gossypii (Hemiptera: Aphididae)
BMC Molecular Biology (2017)
-
Evaluation of high-throughput isomiR identification tools: illuminating the early isomiRome of Tribolium castaneum
BMC Bioinformatics (2017)
-
Transcription Factor Forkhead Regulates Expression of Antimicrobial Peptides in the Tobacco Hornworm, Manduca sexta
Scientific Reports (2017)
-
Targeting chitinase gene of Helicoverpa armigera by host-induced RNA interference confers insect resistance in tobacco and tomato
Plant Molecular Biology (2016)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
