
Abstract
Early neuroimaging studies using Cyberball suggested that social rejection activated the pain matrix, as identified in studies of physical pain. However, these early studies were characterized by small sample sizes. Our statistical multi-level kernel density analysis (MKDA) of Cyberball neuroimaging studies with 244 participants fails to support the claim that social rejection operates on the same pain matrix as nociceptive stimuli, questioning whether social pain is more figurative or literal. We also performed an MKDA of the neuroimaging studies of reliving a romantic rejection to test whether the pain matrix was activated if the rejection were more meaningful. Results again failed to support the notion that rejection activates the neural matrix identified in studies of physical pain. Reliving an unwanted rejection by a romantic partner was significantly characterized by activation within and beyond the “Cyberball” brain network, suggesting that the neural correlates of social pain are more complex than previously thought.
Similar content being viewed by others

Introduction
The functional neuroanatomy of physical pain has been investigated in animal models and human studies for decades1,2,3. By the turn of the century, the first meta-analysis of functional neuroimaging studies of pain in humans was performed to determine the neural marker of pain experience3. A brain pain matrix was identified that included the second somatic region, insular region, dorsal anterior cingulate cortex (dACC), contralateral thalamus, primary somatic area and periaqueductal grey. Three years later, Eisenberger et al. published the first neuroimaging study of social rejection in a sample of 13 participants, showing that rejection led to increased activity in the dACC, insula and the right ventral prefrontal cortex (vPFC) regions4. These results were interpreted as evidence that social rejection operates on the pain matrix to produce social pain5.
Various cultures and languages use terms and phrases for physical pain (e.g., hurt, searing, broken heart) to describe social rejection, but is the similarity between a nociceptive physical stimulus and social rejection literal or figurative? There is a surprisingly large behavioral literature suggesting that the similarity may be literal6,7. The addition of the neuroimaging evidence showing that social rejection activates the pain matrix seemed to make an overwhelming case for social rejection coopting the physical pain system in humans.
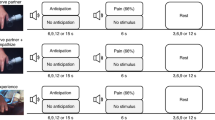
Contrarian views have been espoused, however. For instance, Somerville, Heatherton and Kelley contrasted expectancy violation with social rejection and found that the dACC was affected by expectancy violation, whereas a more ventral ACC was affected by social rejection8. However, Somerville et al. used negative feedback from a confederate to manipulate social rejection, whereas Eisenberger et al. and most of the subsequent neuroimaging studies of social rejection have used the computerized version of Williams “ball-tossing” paradigm, termed “Cyberball”. The participant in Cyberball is represented by an avatar, who is playing catch with two other avatars (who, putatively, represent two other human participants). The game begins with each avatar catching and throwing the ball about a third of the time. In the “inclusion” condition, the participant continues to catch and throw the ball about a third of the time, whereas in the “exclusion” (rejection) condition, the other two avatars throw the ball back and forth and neither looks at or throws the ball to the participant. Williams suggested that: “The ball-tossing paradigm, despite having no verbal interaction and a minimal context, seems to extract the essence of the drama of ostracism. Within a short period of time (less than 5 minutes), ostracism has powerful and fairly consistent effects” (p. 140)7.
There is now a sizeable neuroimaging literature using the Cyberball paradigm that includes whole brain fMRI data from more than 240 participants. Several narrative reviews of this literature have also appeared, with all concluding that social rejection and nociceptive stimuli both activate the physical pain matrix5,9,10,11. Narrative reviews are subject to various biases, however, including the influence of prior expectations, the difficulty in identifying effect sizes, weighting the effect sizes by sample sizes and combining these weighted effect sizes in a rigorous, quantitative fashion.
To quantitatively test for consistency and specificity of regional activation of the studies on social rejection and to examine the notion that social rejection produces social pain in a literal rather than figurative sense (as indexed by the activation of the physical pain matrix), we performed a statistical Multi-level Kernel Density Analysis (MKDA; see Online Method for details)12 using fMRI studies of Cyberball (Supplementary Table 1). This statistical approach quantifies the distribution of significant peak brain coordinates related to social rejection as reported in the Cyberball. As demonstrated by several groups12,13, this approach minimizes biases such as having one study that reports many activation foci from dominating the meta-analysis and it accounts for the smoothness of reported data, false-positive rates and statistical power.
To explore whether the pain matrix might be activated if the social rejection were more pertinent or powerful, we also performed a MKDA of the neuroimaging studies of reliving an unwanted rejection by a romantic partner. With more than 60 participants (Supplementary Table 2), this paradigm is the second most common in neuroimaging studies of social rejection. Participants who have experienced a recent (within the past eight months), unwanted breakup are exposed to photographs of a former romantic partner and are instructed to relive the experience of the unwanted romantic relationship breakup from that significant other. Although there are important differences in these two paradigms, such as the task (a somatosensory task vs. an emotional memory task) and the participant's familiarity and relationship with the person(s) who rejects them (two strangers vs. a beloved significant other), a meta-analysis of these studies bears on the question of whether the similarity between a nociceptive physical stimulus and social rejection is literal or figurative.
Results
The first meta-analysis revealed three main areas were reliably recruited when a participant was rejected by strangers in the Cyberball task: the anterior insula (bilaterally), the left ACC and the left inferior orbito-frontal cortex (Video 1; Supplementary Table 3). Surprisingly, the dACC subdivision, which has been identified as a core region in both theoretical analyses and narrative reviews of this literature9,11, did not emerge in this analysis. To investigate further, we eliminated the minimum cluster size (15) for a region to be considered. Results indicated that 12 voxels were activated in the medial frontal gyrus and one voxel was activated in the left dACC (Supplementary Table 3).
Results from the second meta-analysis also failed to provide unambiguous evidence that rejection activates the neural matrix identified in studies of physical pain 3. Reliving rejection by a significant other revealed four main brain areas that were reliably recruited: the right anterior insula, the right ACC, the left inferior orbito-frontal cortex and the right caudate nucleus (Supplementary Table 4).
Discussion
Button et al.14 recently cautioned that the low statistical power characteristic of contemporary neuroimaging studies not only reduces the chance of detecting true effect but reduces the likelihood that a statistically significant result reflects a true effect. By including 244 participants in the first meta-analysis and 64 participants in the second, the present meta-analyses should provide a better indication of the brain regions activated by social rejection than those provided in any single empirical investigation. Contrary to claims in the extant narrative reviews, our quantitative analysis of participants in the Cyberball paradigm fails to support the claim that social rejection operates on the same neural pain matrix as nociceptive stimuli, raising the possibility that social pain is figurative rather than literal.
The identity of the rejecter may play a role in the modulation of the social pain matrix. Although the most common paradigm used in neuroimaging research to study social rejection is Cyberball, rejection in Cyberball is perpetrated by strangers (actually, a computer program) rather than by a person who is significant in the participant's life. It is possible that rejection by strangers in the Cyberball paradigm is insufficient to activate real social pain. This point was emphasized by Kross and colleagues (2011) in their study15. However, our meta-analysis of studies of reliving an unwanted rejection by a romantic partner also failed to provide clear evidence for the activation of the dACC or the pain matrix more generally. One putative distinction made between social and physical pain is that memories can strongly trigger social but not physical pain7,16. It is possible that reliving a rejection by a significant other arouses emotions but is no more likely to activate the pain matrix than is reliving a nociceptive stimulus. The Cyberball paradigm does involve a real-time social rejection, however, so the possible effects of reliving social pain cannot account for the results of the first meta-analysis.
If not social pain, what might the neural correlates identified in these meta-analyses reflect? The anterior insula (bilaterally), the left ACC and the left inferior orbito-frontal cortex were reliably activated in the Cyberball paradigm, whereas the right anterior insula, the right ACC, the left inferior orbito-frontal cortex and the right caudate nucleus were reliably activated in the reliving paradigm. Evidence exists showing that the joint activation of the anterior insula and anterior cingulate is involved in attention and emotion and, together, may play a role in transforming sensory inputs into subjective feelings and behavior17. These regions and the caudate nucleus have also been observed in studies of uncertainty18,19,20, rumination and emotional craving21 and in the abstract mental representation of significant others and interpersonal relationship13,22. The neural network elicited in these studies may therefore reflect social uncertainty, rumination, distress and craving rather than social pain per se.
In sum, our results suggest that there is presently insufficient neuroimaging evidence to conclude that social rejection operates on the putative pain matrix defined in the narrative reviews of the social rejection literature5,9,10,11 and they raise the possibility that the neural correlates of social pain are different and more complex than previously thought. Moreover, our results emphasize the value of quantitative over narrative reviews of the neuroimaging literature and underscore the importance of the recommendation by Button and colleagues14 to perform neuroimaging studies with sufficient statistical power to identify reliable effects.
Methods
Literature search
We performed a systematic review of functional magnetic resonance imaging (fMRI) studies of social rejection/isolation in healthy individuals. All papers in the literature published through March 2013 (inclusive) were considered for this meta-analysis, subject to two general limitations: the scientific publication had to be a peer-reviewed manuscript; and the title and abstract had to be available in English. Materials were identified through computer-based search and through relevant review papers, as described below.
We used MEDLINE library through PubMed database. The search for fMRI studies on social rejection/pain had no restrictions on the date of the study. We used the keywords "fMRI", "social rejection", “social isolation”, “ostracism”, "social pain", "social exclusion", “cyberball”, “rejection”, “love” and “romantic break-up”. In addition, we searched the reference lists of articles and reviews identified by this search strategy.
Selection Criteria
The set of scientific publications identified was then subjected to the following narrower criteria: 1) participants had to undergo "social pain" through experimental manipulation: either through the standard “cyberball tossing task” behavioral paradigm, or through the second most common paradigm in neuroimaging studies of social rejection i.e., the romantic breakup paradigm, which consists of viewing photographs of a former romantic partner and reliving the experience of the unwanted romantic relationship breakup with that romantic partner; 2) participants must experience a rejection by someone, who no longer wants to be/interact with the participant (This means studies on bereavement cannot be included, as the deceased person probably did not die with the aim to reject the participant); 3) participants must have undergone a functional magnetic resonance imaging (fMRI) scan; 4) participants must have been free of psychiatric or neurological disease and drug or alcohol abuse as ascertained by the authors; 5) participants must have been 18 years old or more; and 6) all studies concerning social pain have been conducted in accordance with ethical standards and under the supervision of the responsible local human subject's committees.
In addition, the following methodological inclusion criteria were included: 1) the fMRI study must report a whole brain analysis of a social pain against a non-social pain control condition (rather than a resting state condition); and 2) the reported activations cannot be conflated with other factors, such as drug effect correlations, correlations with inflammatory variables, or correlations with psychological variables.
Meta-Analysis methods
We calculated the number of statistical contrast maps that activated each voxel in the brain using 10 mm kernel and used Monte Carlo simulations (10,000 iterations) to obtain a threshold and establish statistical significance against a null hypothesis that activated regions in the resulting pair-wise contrast maps are not spatially consistent (i.e. that they are randomly distributed throughout the brain). The reported peak coordinates within a study's contrast maps are weighted in the meta-analysis by study quality and sample size.
The use of the distribution of maximum values provides a strong control of family-wise error rate and is an established method for multiple comparisons correction. All voxels (which constitute the various brain region volumes) whose density exceeded the 99.9th percentile value under the null hypothesis were considered significant (i.e., Family-Wise Error Rate corrected for spatial extent at P < .05). Brain coordinates obtained using the MKDA method above were entered into the SPM8 software program (Wellcome Trust Centre for Neuroimaging, London; http://www.fil.ion.ucl.ac.uk/spm/) using Anatomy Toolbox version 1823 in order to facilitate identification and labeling of each activation peak. The MNI coordinates and volumes of significantly overlapping clusters (brain foci) were extracted using the AFNI software plug-in 3dcluster24. Displayed localizations were further validated by visual inspection relative to the Tzourio-Mazoyer et al.’s automated anatomical parcellation of the spatially normalized single-subject high-resolution T1 volume provided by the Montreal Neurological Institute25 in SPM8. Brain video of the results was created with the Amira scientific visualization software package version 5.4 (www.vsg3d.com) where it was volume rendered with a linearly varying color/alpha map. The volume rendered brain model upon which the fMRI activation rendering is overlaid was generated from the Colin head model obtained at 1 mm spatial resolution that was manually segmented to extract the brain voxels and then down sampled to 2 mm spatial resolution to match the resolution of the fMRI activation imagery. The down sampled brain model was volume rendered in a low alpha range to provide transparency and allow for visualization of fMRI activation sites inside the brain volume.
References
Wager, T. D. & Atlas, L. Y. How Is Pain Influenced by Cognition? Neuroimaging Weighs In. Perspectives on Psychological Science 8, 91–97 (2013).
Atlas, L. Y., Bolger, N., Lindquist, M. a. & Wager, T. D. Brain Mediators of Predictive Cue Effects on Perceived Pain. Journal of Neuroscience 30, 12964–12977 (2010).
Peyron, R., Laurent, B. & García-Larrea, L. Functional imaging of brain responses to pain. A review and meta-analysis (2000). Neurophysiologie clinique Clinical neurophysiology 30, 263–288 (2000).
Eisenberger, N. I., Lieberman, M. D. & Williams, K. D. Does rejection hurt? An FMRI study of social exclusion. Science 302, 290–2 (2003).
Eisenberger, N. I. The pain of social disconnection: examining the shared neural underpinnings of physical and social pain. Nature reviews. Neuroscience 13, 421–34 (2012).
Macdonald, G. & Leary, M. R. Why does social exclusion hurt? The relationship between social and physical pain. Psychological Bulletin 131, 202–223 (2005).
Williams, K. D. Ostracism: The power of silence. New York 20, 141–142 (Guilford Press: 2001).
Somerville, L. H., Heatherton, T. F. & Kelley, W. M. Anterior cingulate cortex responds differentially to expectancy violation and social rejection. Nature Neuroscience 9, 1007–8 (2006).
Eisenberger, N. I. & Cole, S. W. Social neuroscience and health: neurophysiological mechanisms linking social ties with physical health. Nature neuroscience 15, 669–74 (2012).
Eisenberger, N. I. & Lieberman, M. D. Why rejection hurts: a common neural alarm system for physical and social pain. Trends in Cognitive Sciences 8, 294–300 (2004).
Lieberman, M. D. & Eisenberger, N. I. Neuroscience. Pains and pleasures of social life. Science 323, 890–891 (2009).
Wager, T. D., Lindquist, M. A., Nichols, T. E., Kober, H. & Van Snellenberg, J. X. Evaluating the consistency and specificity of neuroimaging data using meta-analysis. NeuroImage 45, S210–21 (2009).
Cacioppo, S., Bianchi-Demicheli, F., Frum, C., Pfaus, J. G. & Lewis, J. W. The Common Neural Bases Between Sexual Desire and Love: A Multilevel Kernel Density fMRI Analysis. The Journal of Sexual Medicine 9, 1048–54 (2012).
Button, K. S. et al. Power failure: why small sample size undermines the reliability of neuroscience. Nature Revews Neurosci 14, 365–376 (2013).
Kross, E., Berman, M. G., Mischel, W., Smith, E. E. & Wager, T. D. Social rejection shares somatosensory representations with physical pain. Proceedings of the National Academy of Sciences of the United States of America 108, 6270–5 (2011).
Chen, Z., Williams, K. D., Fitness, J. & Newton, N. C. When hurt will not heal: exploring the capacity to relive social and physical pain. Psychological Science 19, 789–795 (2008).
Medford, N. & Critchley, H. D. Conjoint activity of anterior insular and anterior cingulate cortex: awareness and response. Brain structure function 214, 535–549 (2010).
Feinstein, J. S., Stein, M. B. & Paulus, M. P. Anterior insula reactivity during certain decisions is associated with neuroticism. Social cognitive and affective neuroscience 1, 136–142 (2006).
Grinband, J., Hirsch, J. & Ferrera, V. P. A neural representation of categorization uncertainty in the human brain. Neuron 49, 757–763 (2006).
Singer, T., Critchley, H. D. & Preuschoff, K. A common role of insula in feelings, empathy and uncertainty. Trends in Cognitive Sciences 13, 334–340 (2009).
Fisher, H. E., Brown, L. L., Aron, A., Strong, G. & Mashek, D. Reward, addiction and emotion regulation systems associated with rejection in love. Journal of neurophysiology 104, 51–60 (2010).
Wager, T. D. & Barrett, L. F. From affect to control: Functional specialization of the insula in motivation and regulation. Emotion 129, 2865 (2004).
Eickhoff, S. B. et al. A new SPM toolbox for combining probabilistic cytoarchitectonic maps and functional imaging data. NeuroImage 25, 1325–1335 (2005).
Cox, R. W. AFNI: software for analysis and visualization of functional magnetic resonance neuroimages. Computers and biomedical research an international journal 29, 162–173 (1996).
Tzourio-Mazoyer, N. et al. Automated anatomical labeling of activations in SPM using a macroscopic anatomical parcellation of the MNI MRI single-subject brain. NeuroImage 15, 273–289 (2002).
Acknowledgements
The authors are supported by the University of Chicago, the Center for Cognitive and Social Neuroscience (CCSN). This work was supported by grants from the Swiss National Science Foundation (Grant #PP00_1_128599/1 to SC), Department of the Army, Defense Medical Research and Development Program, (Agency Award #W81XWH-11-2-0114 to JTC) and NCRR NIH COBRE Grant E15524 (Grant #E15524 to the Sensory Neuroscience Research Center of West Virginia University to JWL). The authors are grateful for the support of the University of Chicago CCSN and the Research Computing Center for assistance with the brain videos produced in this work.
Author information
Authors and Affiliations
Contributions
S.C. and J.T.C. wrote the main manuscript text; C.F. and J.W.L. performed the M.K.D.A. analyses, E.A. help with the literature search and R.W. prepared the video. All authors reviewed the manuscript.
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Electronic supplementary material
Supplementary Information
Video 1
Supplementary Information
Supplementary Information
Rights and permissions
This work is licensed under a Creative Commons Attribution-NonCommercial-NoDerivs 3.0 Unported License. To view a copy of this license, visit http://creativecommons.org/licenses/by-nc-nd/3.0/
About this article
Cite this article
Cacioppo, S., Frum, C., Asp, E. et al. A Quantitative Meta-Analysis of Functional Imaging Studies of Social Rejection. Sci Rep 3, 2027 (2013). https://doi.org/10.1038/srep02027
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/srep02027
This article is cited by
-
Compulsive Sexual Behavior Disorder and Problematic Pornography Use in the Context of Social Ties
Current Addiction Reports (2024)
-
The multi-level outcome study of psychoanalysis for chronically depressed patients with early trauma (MODE): rationale and design of an international multicenter randomized controlled trial
BMC Psychiatry (2023)
-
Neural correlates of social exclusion and overinclusion in patients with borderline personality disorder: an fMRI study
Borderline Personality Disorder and Emotion Dysregulation (2023)
-
Effects of acute psychosocial stress on source level EEG power and functional connectivity measures
Scientific Reports (2023)
-
Script-driven imagery of socially salient autobiographical memories in major depressive disorder
Scientific Reports (2023)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
