
Abstract
The network of roads on the landscape is vast and contributes a suite of negative ecological effects on adjacent habitats, ranging from fragmentation to contamination by runoff. In addition to the immediate consequences faced by biota living in roaded landscapes, road effects may further function as novel agents of selection, setting the stage for contemporary evolutionary changes in local populations. Though the ecological consequences of roads are well described, evolutionary outcomes remain largely unevaluated. To address these potential responses in tandem, I conducted a reciprocal transplant experiment on early life history stages of a pool-breeding salamander. My data show that despite a strong, negative effect of roadside pools on salamander performance, populations adjacent to roads are locally adapted. This suggests that the response of species to human-altered environments varies across local populations and that adaptive processes may mediate this response.
Similar content being viewed by others

Introduction
The global network of roads has expanded rapidly over the last half century and continues to increase across the earth's surface1,2. In the United States, an estimated 13.7 million lane km of roads bisect the landscape3 and cover approximately 1% of the land4. A random point placed somewhere within the coterminous United States is typically less than 1 km from the nearest road5. Traffic on this network has increased fourfold in as many decades, from 1960 – 20006. Despite the obvious benefit of roads for transport, accidents on roads result in substantial loss of human life and increased economic burdens. Globally, annual estimates of road fatalities exceed 750,000, while those of economic cost of crashes exceed US $500 billion7. The ecological consequences caused by the presence and increasing use of roads are manifold and predominantly negative in effect. For example, roadkill causes an estimated one million vertebrate deaths per day in the United States, while habitat fragmentation elicits a host of indirect effects8. Surface runoff and leaching result in the deposition of myriad chemical contaminants into adjacent habitats9. Collectively, these effects are reported to extend well beyond the immediate footprint of roads and are estimated to ecologically affect 19% of the land in the United States4. These consequences of roads are especially well described for amphibians, for which studies have revealed patterns of reduced abundance and diversity10, movement11 and gene flow12 along with increases in prevalence of malformations13. Similarly, experimental exposure to road adjacency and chemical runoff has generally negatively influenced embryonic and larval amphibian performance traits, such as growth, development and survival14,15.
These ecological studies have given us keen insights into immediate outcomes associated with roads. These insights, however, are generally based on traditional ecological frameworks, which have assumed that biological responses to negative environmental effects are fixed, ignoring the possibility that evolutionary processes act fast enough to matter. However, we now recognize that evolutionary and ecological processes interact strongly16 and that contemporary evolution influences ecological outcomes across small spatial and temporal scales17. The potential for this influence may be especially pronounced in human-altered contexts, where rates of phenotypic change are high relative to undisturbed settings18. For example, studies of the moor frog (Rana arvalis) have revealed adaptation of acid tolerance in populations that breed in low pH aquatic environments with a history of anthropogenic inputs of acidifying agents19,20,21. Thus, from an eco-evolutionary perspective, the negative influences of roads should be viewed not only as threats to persistence, but also as potential agents of natural selection, setting the stage for adaptive responses.
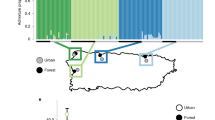
In light of this perspective, I evaluated the effects of road adjacency on a pool-breeding amphibian while accounting for the possibility that local populations might exhibit differential responses mediated by evolutionary processes. Specifically, I hypothesized 1) that roadside pools would induce negative effects on early aquatic life stages of the spotted salamander (Ambystoma maculatum) and 2) that within roadside pools, local populations would outperform foreign populations as predicted by the local adaptation hypothesis22. I evaluated these hypotheses by conducting a reciprocal transplant experiment across 10 natural temporary pools—five roadside and five woodland—comprising typical breeding sites for the spotted salamander (Fig. 1). I measured survival, growth and development of hatchling stage larvae. These phenotypic variables have been broadly categorized as performance or viability traits that have been shown in amphibians to correlate positively with survival at later life history stages23. That they have further been shown in amphibians to be heritable24 and undergo selection23 suggests they are suitable phenotypic indicators of local adaptation and population success in this system. I used mixed models to analyze these responses in relation to the interaction between population (hereafter deme) and grow-out environment (the so-called genotype by environment, or G x E, interaction). To test for local adaptation, I compared the responses of “local vs. foreign” demes in the roadside environment. This criterion is regarded as diagnostic of local adaptation when the local deme has higher fitness than the foreign deme22. Because the effect of environment may be inherited maternally yet independently of genotype, I also tested embryo size, a maternally mediated trait known to confer positive effect on offspring survival and growth in amphibians (reviewed by25). Lastly, to begin to understand the mechanism by which road proximity influences salamander performance, I characterized the environment at each pool by estimating a suite of abiotic variables (see Methods) associated with amphibian distribution and performance26. I expected that these characteristics might vary with road proximity and thus act as potential agents of natural selection in roadside pools.

Reciprocal transplant design showing locations of each of 10 pools in northeastern Connecticut, USA.
Like symbols indicate paired pools. Red symbols represent roadside pools; blue symbols represent woodland pools. The interstate highway (I-84) and on/off-ramp infrastructure is indicated in yellow. Primary roads are heavily shaded, while secondary and unpaved roads are lightly shaded. Inset indicates approximate study site location within Connecticut.
Results
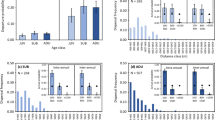
The roadside environment had a strong negative effect on embryonic salamander survival, regardless of deme (Posterior mean = −1.48; 95% CI = −2.40 – −0.42; Pmcmc = 0.005). Survival in roadside pools averaged just 56%, as compared to 87% in woodland pools (Fig. 2). Thus, an average of 36% fewer individual embryos survived to hatching in roadside versus woodland pools. I also found that survival differed with respect to the G x E interaction (Posterior mean = 1.12; 95% CI = 0.13 – 2.10; Pmcmc = 0.028. Specifically, within roadside pools, the roadside deme outsurvived the woodland deme by an average of 25% (Fig. 2). While the difference in survival is substantial within roadside pools, I did not detect evidence of a tradeoff to survival in woodland pools. Namely, average survival within woodland pools was statistically equivalent between the two demes (Posterior mean = −0.05; 95% CI = −0.81 – 0.77; Pmcmc = 0.889). With regard to developmental and growth rates, neither trait varied across the G x E interaction, nor with respect to deme. However, regardless of deme, embryos raised in roadside pools developed at a 5.5% slower rate on average than those raised in woodland pools (Posterior mean = −0.004; 95% HPD = −0.0063 – −0.0024; Pmcmc < 0.001). The inclusion of embryo size did not affect phenotype or qualitatively alter the G x E interaction in any of the models. However, embryo size itself differed between the two environments: at the time of collection, embryos from roadside pools were estimated to be 11.3% smaller on average than those from woodland pools (MCMC mean = −0.81; 95% HPD = −1.58 – −0.79; Pmcmc = 0.033). With regard to abiotic conditions, only specific conductance varied with respect to pool environment. Namely, specific conductance averaged 886 µS in roadside pools, as compared to 28 µS in woodland pools, yielding a 32-fold increase in conductivity as a result of road proximity. Accordingly, analysis of water samples revealed that chloride ions were highly elevated in roadside pools, averaging 188 mg/l, as compared to 2.8 mg/l in woodland pools.

Spotted salamander survival across the G x E interaction.
Survival (± SEM) is shown here as the mean proportion of individuals surviving to hatching across all experimental units (N = 100). The woodland deme is represented by open circles (O) while the roadside deme is represented by filled squares (▪). The environment in which the animals were grown out is on the x-axis.
Discussion
The impacts on salamander larvae born into roadside pools in this system are severe. Given the context for carry-over effects of the aquatic environment onto juvenile and adult survival in amphibians23,27, this result supports the hypothesis that roadside pools induce negative consequences for spotted salamanders. More broadly, these findings provide further evidence that roads negatively affect biota occupying road adjacent habitats. Yet critically, while the outcome described here corroborates a body of literature reporting negative road effects, it reveals that even across small spatial scales, local population responses can describe substantial variation in the capacity to tolerate human mediated environmental change28. The interaction between deme and environment demonstrates that roadside demes of the spotted salamander are adapted to the negative influence of roadside pools. Though such adaptation does not equate success, it indicates that demes inhabiting roadside pools are more capable of surviving there than those populations from just hundreds of meters away. This suggests that even though roadside environments are harsh places to live, some species—even vertebrates—may adapt. More broadly, given that the differential responses detected across populations here would remain opaque to traditional ecological approaches, eco-evolutionary approaches may redefine our current understanding of road effects.
The G x E interaction found here suggests a role for a genetic component in explaining the pattern of local adaptation. Yet, given that individuals in this experiment were wild captures, non-genetic properties such as maternal effects may also have influenced the differential responses between demes. To evaluate the role for maternal effects, I estimated embryo size, a trait found to be maternally mediated and associated with offspring performance in amphibians29. I found that embryo size did not affect phenotype or qualitatively alter the G x E interaction. Thus, there is no evidence of egg size mediated maternal effects influencing the pattern of local adaptation in this system. This conclusion is further supported by the observation that embryos from roadside pools were on average 11.3% smaller than those from woodland pools. Separately, this distinction could reflect differential allocation of resources by female spotted salamanders to reproduction in these two environments. Such trends are most often explained by a “quantity over quality” tradeoff. However, in a related study, spotted salamander egg masses in roadside pools contain 24% fewer eggs on average than those from woodland pools30, a pattern inconsistent with such a tradeoff. In this system, even though eggs from the roadside deme were smaller, they outsurvived those from the woodland deme. This pattern runs counter to typical maternal effects, in which size is positively correlated with subsequent performance (but see31). While positive maternal effects may be mediated by egg quality, rather than size, emerging literature reports that in degraded environments, maternal effects exert negative influence on offspring success as a result of contaminant transfer32. While this does not rule out the potential influence of maternal effects, it does suggest that maternal effects alone may not be adequate to explain this pattern of local adaptation. As an alternative explanation for the relative difference in survival within roadside pools, woodland populations may simply be less tolerant of translocation effects. However, in a related common garden experiment comprising identical families and populations (S.P.B., unpublished data), overall survival was high (90%) and did not differ between roadside and woodland demes (MCMC mean = −0.02; 95% HPD = −0.17 – 0.14; Pmcmc = 0.810). This suggests that any effect of translocation is minimal and equivalent between the woodland and roadside demes. Thus, the relative increase in survival shown by roadside demes is not generic to novel environments, but instead represents an adaptation to roadside pools.
Together, these findings point to the potential influence of genetic differentiation between roadside and woodland demes, suggesting that the adaptation to roadside pools is evolved. Yet, our knowledge of gene flow in this species suggests that populations are structured across relatively larger spatial and temporal scales than exist across the roadside-woodland gradient in this system33,34. However, evidence for high site fidelity in the spotted salamander35 coupled with our understanding that roads restrict and even collate, gene flow36 suggests that local genetic differentiation in this system is indeed a possibility. It is further worth noting that our current understanding of gene flow in the spotted salamander is based on neutral markers, which do not reflect variation in genes experiencing rapid selection. Thus, future studies of genetic structure in this system that interrogate non-neutral markers—capable of revealing differentiation associated with contemporary adaptive responses—hold great promise in parsing the relative contributions of environmental and genetic mechanisms of variance.
From among the suite of abiotic variables estimated across pools, only specific conductance varied with respect to road adjacency. As a measure of electrical conductivity, specific conductance of water is known to increase with the presence of ions and metals, both of which are commonly found in runoff9. Thus, runoff in this system is likely contributing a suite of contaminants into roadside pools. Among these, roadsalt is a likely constituent given the context of winter road deicing in the region. Indeed, the elevated levels of chloride ions detected in roadside pools are likely the result of the application and subsequent runoff of roadsalt, which is predominantly composed of sodium chloride in the northern U.S. and within the study region37. Together, this suggests that for aquatic stage amphibians, runoff may be the critical constituent of road proximity acting as an agent of natural selection and capable of inducing adaptive responses.
To my knowledge, this is the first phenotypic evidence of local adaptation to roadside habitats by a vertebrate. The divergent response revealed here demonstrates that even when exposed to novel, harsh conditions induced by contemporary anthropogenic disturbance, these vertebrates are capable of adaptation on contemporary timescales and across small scales of space. Indeed, aerial imagery38 suggests that four of the five roaded sites within this study were first paved 44 to 58 years prior (with the fifth at least 75 years prior) and pond pairs ranged from 880 – 6060 m apart. However, these results should be interpreted cautiously given two key assumptions: first, that performance traits reported here are positively correlated with fitness and second, that the observed patterns of differentiation are not reversed at later life-history stages. Evidence from related species and systems suggest these assumptions are valid, yet they remain untested across these demes. In addition to these assumptions, it is worth considering that embryos were exposed to roadside pool water for up to 48 hours prior to the start of the experiment. It is conceivable that this small window of early exposure improved hatchling survival via plastic responses associated with reaction norms. However, this seems unlikely given the strong negative effect of roadside pools.
Regardless of mechanism, these results provide support for an alternative perspective on the dialogue concerning species responses and the environments being reshaped by human activities39. However, it remains to be determined whether the adaptation observed in this system will facilitate population persistence given the overall negative effect of roadside pools. The long-term success of such populations will depend on the rate of environmental change in relation to factors such as functional genetic diversity, which may constrain responses.
Methods
Natural history and site selection
The spotted salamander (Ambystoma maculatum) is widely distributed throughout eastern North America, with a range extending from southern Quebec to the southeastern United States40. Within the study region, spotted salamanders breed in late March or early April, when adults migrate from upland terrestrial habitat into ephemeral wetlands to reproduce. In this study region, females oviposit egg masses containing approximately 100 embryos per mass. Embryos develop over 8–10 weeks before hatching and continue to develop as aquatic larvae throughout the summer until they metamorphose into terrestrial juveniles41.
I used National Wetland Inventory Maps42 and visual driving searches to identify roadside pools located less than 10 m from a paved road within the Yale Myers Forest region in northeastern CT (Fig. 1 inset), an area characterized by large swaths of native forest and low human population density. In spring 2009, I selected for experiment the five roadside pools believed to be most influenced by runoff by choosing those with the highest specific conductance values. I then assigned to each roadside pool a woodland pool, located at least 200 m from the nearest paved road and thereby yielding five unique pool pairs. To control for confounding variation, I selected woodland pools that minimized inter-pair distance, while at the same time maximized similarity in pool size, forest canopy cover and emergent vegetation. Reciprocal transplants were conducted within each of these five pairs (Fig. 1).
Reciprocal transplant experiment
In the days leading up to breeding (signaled by the appearance of conspecific egg masses in like pools located in central and southern Connecticut), I monitored pools daily for the arrival of new egg masses. From each pool, I collected a subset of embryos from egg masses less than 48 hours old, with a target of 10 egg masses per pool. There were no conspicuous differences in the embryos themselves between sites and selection of embryos from within each egg mass was haphazard. Thus, any potential effect of the < 48 hour exposure did not bias the subset of embryos employed in the experiment. I selected egg masses that were spatially distributed (if possible)43 and conspicuously large and distinct so as to avoid sampling masses that might have originated from the same female. From each egg mass, I carefully dissected out two clusters of 10 embryos. One cluster was stocked into one of five experimental enclosures in the origin pool while the other was relocated (suspended in a small enclosure containing pool water and incubated within an iced cooler) to the transplant pool and also assigned to one of five experimental enclosures. Each pool contained five experimental blocks, with each block containing one local enclosure and one transplanted enclosure. I targeted stocking 10 embryos from each of two egg masses per enclosure, yielding one unique pairing of egg masses (hereafter “clutch pair”) that was replicated across, but not within pool pairs. This design was chosen to maximize family level diversity while maintaining an additional level of replication at the clutch pair level, while at the same time balancing logistics of resources and spatial constraints within pools. Enclosure assignment was haphazard for each clutch pair. Because two woodland pools presented fewer than 10 oviposited clutches (n = 6, n = 8 respectively), a total of 88 enclosures each were stocked with 20 embryos (from 44 unique clutch pairs) while 12 enclosures each were stocked with 10 embryos (from 6 unique clutch pairs). Thus, across 10 pools, I stocked a total of 100 enclosures, 88 of which contained 20 embryos each, while 12 contained 10 embryos each. At the conclusion of the experiment, when all eggs had either hatched or died, I estimated hatchling survival, developmental rate and growth rate.
Each enclosure consisted of a 14 l plastic container equipped with six 7 cm diameter ventilation holes on the sides and a 25×13 cm hole in the lid. I screened over the side ventilation holes with Noseum mesh (ca. 97 holes per cm2) affixed with Gorilla Glue around the edges, thus creating a barrier to aquatic predators, yet facilitating water flow. Mesh was also affixed to the top ventilation hole using Gorilla Glue and Duct Tape. The inside of each enclosure was fitted with a piece of hardware cloth that acted as a cradle to support egg clusters off the bottom of the enclosure. Two pieces of closed cell blue foam (ca. 2.5×5×36 X cm) were secured to the long sides of the enclosures with Silicone II and stainless steel screws. This provided flotation such that embryos were submerged but suspended at a height comparable to that of naturally oviposited egg masses.
Size and developmental stage of eggs and hatchlings
Immediately after stocking eggs into field enclosures, I placed egg masses on ice and returned them to the laboratory in New Haven, CT. There, I dissected approximately five eggs from each egg mass and placed them in a glass petri dish in which photographs were captured with a digital camera attached to a dissecting stereomicroscope. Vitelline membranes surrounding embryos were left in place. I used ImageJ software44 to estimate the two-dimensional surface area of each embryo represented by a best-fit ellipse. I also estimated the developmental stage of each egg mass from this sample45. At the end of the experiment, I set aside a subset of surviving hatchlings from each enclosure for a separate experiment and analysis. From the remaining survivors, I then preserved a target of three, haphazardly selected individuals. For each of these individuals, I used a dissecting stereomicroscope to assess snout-vent length (SVL) and developmental stage46.
Characterizing roadside and woodland environments
In each pool, I measured seven environmental characteristics associated with amphibian distribution and performance26. Specific conductance, dissolved oxygen, pH and wetland depth were measured twice (20 April and 22 May 2009) during the experiment, while temperature was measured every thirty minutes using deployed temperature loggers. All water parameter measurements were taken 10 cm below the surface at the location of the deepest point in each pool. In roadside pools, specific conductance was also measured at the base of the water column because a strong, vertical halocline is present. I therefore chose to analyze and report specific conductivity as the mean value taken from the top and bottom of each roadside pool. To estimate the influence of forest canopy cover, I captured leaf-off hemispherical photographs (on 1 April 2011) at each of five locations per pool (2 m from shore at each of the four cardinal compass points plus the approximate center of pool). I used HemiView software47 to estimate global site factor (GSF)—a measure of solar radiation reaching the water surface (see48). GSF was calculated for each of two dates spanning the experiment (10 Apr; 16, May). From the overall temperature dataset, I calculated average temperature over each of two time periods, 06 – 28 April 2009 and 29 April − 20 May 2009. I also collected water samples over two periods to assay the concentration of chloride ions using liquid chromatography.
Statistical analyses
All statistical analyses were conducted in R V. 2.13.149. I composed a suite of mixed effects models to evaluate performance variables across the G x E interaction. Specifically, I analyzed survival as a bivariate response of successes and failures, while I analyzed derived growth and developmental rates separately as univariate responses. All models of performance variables were composed with and without embryo size as a covariate in order to estimate the potential influence of egg size mediated maternal effects. Each response variable was fit across a suite of models that differed in random effects structure (Table S1). For each response variable, I evaluated for inference the most parsimonious model with respect to both the significance of interacting main effects and the random effect structure as indicated by lowest AIC or DIC scores for Gaussian and binomial responses respectively (Table S2). I also used mixed effects models to evaluate the influence of pool type on both embryo size and the suite of abiotic variables. For all models, I used MCMC randomization methods to conduct inference50. Complete details regarding these analyses and their respective estimates are found in Supporting Information.
References
Doyle, M. W. & Havlick, D. G. Infrastructure and the environment. Annu. Rev. Env. Resour. 34, 349–373 (2009).
Canning, D. A database of world stocks of infrastructure, 1950–95. World Bank Econ. Rev. 12, 529 (1998).
Bureau of Transportation Statistics. ed U.S. Deptartment of Transportation. Washington, DC (2011).
Forman, R. T. T. Estimate of the area affected ecologically by the road system in the United States. Conserv. Biol. 14, 31–35 (2000).
Riitters, K. H. & Wickham, J. D. How far to the nearest road? Front. Ecol. Environ. 1, 125–129 (2003).
National Research Council. Assessing and Managing the Ecological Impacts of Paved Roads. (The National Academies Press, 2005).
Jacobs, G., Aeron-Thomas, A. & Astrop, A. Estimating global road fatalities. TRL report 445 (London, England: Transportation Research Laboratory, 2000).
Forman, R. T. T. & Alexander, L. E. Roads and their major ecological effects. Annu. Rev. Ecol. Syst. 207–231 (1998).
Trombulak, S. C. & Frissell, C. A. Review of ecological effects of roads on terrestrial and aquatic communities. Conserv. Biol. 14, 18–30 (2000).
Vos, C. C. & Chardon, J. P. Effects of habitat fragmentation and road density on the distribution pattern of the moor frog Rana arvalis. J. Appl. Ecol. 35, 44–56 (1998).
Gibbs, J. P. Amphibian movements in response to forest edges, roads and streambeds in southern New England. J. Wildl. Manage. 62, 584–589 (1998).
Marsh, D. et al. Effects of roads on patterns of genetic differentiation in red-backed salamanders, Plethodon cinereus. Conserv. Genet. 9, 603–613 (2008).
Reeves, M. K., Dolph, C. L., Zimmer, H., Tjeerdema, R. S. & Trust, K. A. Road proximity increases risk of skeletal abnormalities in wood frogs from National Wildlife Refuges in Alaska. Environ. Health Perspect. 116, 1009–1014 (2008).
Karraker, N. E., Gibbs, J. P. & Vonesh, J. R. Impacts of road deicing salt on the demography of vernal pool-breeding amphibians. Ecol. Appl. 18, 724–734 (2008).
Turtle, S. L. Embryonic survivorship of the spotted salamander (Ambystoma maculatum) in roadside and woodland vernal pools in southeastern New Hampshire. J. Herpetol. 34, 60–67 (2000).
Carroll, S. P., Hendry, A. P., Reznick, D. N. & Fox, C. W. Evolution on ecological time-scales. Funct. Ecol. 21, 387–393 (2007).
Hargeby, A., Johansson, J. & Ahnesjö, J. Habitat-specific pigmentation in a freshwater isopod: Adaptive evolution over a small spatiotemporal scale. Evolution 58, 81–94 (2004).
Hendry, A. P., Farrugia, T. J. & Kinnison, M. T. Human influences on rates of phenotypic change in wild animal populations. Mol. Ecol. 17, 20–29 (2008).
Andrén, C., Mårdén, M. & Nilson, G. Tolerance to low pH in a population of moor frogs, Rana arvalis, from an acid and a neutral environment: A possible case of rapid evolutionary response to acidification. Oikos 56, 215–223 (1989).
Merilä, J., Söderman, F., O'Hara, R., Räsänen, K. & Laurila, A. Local adaptation and genetics of acid-stress tolerance in the moor frog, Rana arvalis. Conserv. Genet. 5, 513–527 (2004).
Persson, M., Räsänen, K., Laurila, A. & Merilä, J. Maternally determined adaptation to acidity in Rana arvalis: Are laboratory and field estimates of embryonic stress tolerance congruent? Can. J. Zool. 85, 832–838 (2007).
Kawecki, T. J. & Ebert, D. Conceptual issues in local adaptation. Ecol. Lett. 7, 1225–1241 (2004).
Altwegg, R. & Reyer, H.-U. Patterns of natural selection on size at metamorphosis in water frogs. Evolution 57, 872–882 (2003).
Pakkasmaa, S., Merilä, J. & O'Hara, R. B. Genetic and maternal effect influences on viability of common frog tadpoles under different environmental conditions. Heredity 91, 117–124 (2003).
Mousseau, T. E. & Fox, C. W. in Maternal Effects, Developmental Plasticity and Life History Evolution 244–260 (Oxford University Press, New York, 1998).
Wellborn, G. A., Skelly, D. K. & Werner, E. E. Mechanisms creating community structure across a freshwater habitat gradient. Annu. Rev. Ecol. Syst. 27, 337–363 (1996).
Chelgren, N. D. Carryover aquatic effects on survival of metamorphic frogs during pond emigration. Ecol. Appl. 16, 250 (2006).
Skelly, D. K. Microgeographic countergradient variation in the wood frog, Rana sylvatica. Evolution 58, 160–165 (2004).
Laugen, A. T., Laurila, A. & Merilä, J. Maternal and genetic contributions to geographical variation in Rana temporaria larval life-history traits. Biol. J. Linn. Soc. 76, 61–70 (2002).
Karraker, N. & Gibbs, J. Contrasting road effect signals in reproduction of long- versus short-lived amphibians. Hydrobiologia 664, 213–218 (2011).
Dziminski, M. A. & Roberts, J. D. Fitness consequences of variable maternal provisioning in quacking frogs (Crinia georgiana). J. Evol. Biol. 19, 144–155 (2006).
Hopkins, W. A., DuRant, S. E., Brandon Patrick, S., Rowe, C. L. & Jackson, B. P. Reproduction, embryonic development and maternal transfer of contaminants in the amphibian Gastrophryne carolinensis. Environ. Health Perspect. 114, 661–666 (2006).
Zamudio, K. R. & Wieczorek, A. M. Fine-scale spatial genetic structure and dispersal among spotted salamander (Ambystoma maculatum) breeding populations. Mol. Ecol. 16, 257–274 (2007).
Purrenhage, J. L., Niewiarowski, P. H. & Moore, F. B. G. Population structure of spotted salamanders (Ambystoma maculatum) in a fragmented landscape. Mol. Ecol. 18, 235–247 (2009).
Vasconcelos, D. & Calhoun, A. Movement patterns of adult and juvenile Rana sylvatica (LeConte) and Ambystoma maculatum (Shaw) in three restored seasonal pools in Maine. J. Herpetol. 38, 551–561 (2004).
Holderegger, R. & Di Giulio, M. The genetic effects of roads: A review of empirical evidence. Basic Appl. Ecol. 11, 522–531 (2010).
Mullaney, J. R., Lorenz, D. L. & Arntson, A. D. Chloride in groundwater and surface water in areas underlain by the glacial aquifer system, northern United States. U.S. Geological Survey Scientific Investigations Report 2009–5086 (2009).
Connecicut State Library. (Hartford, CT, USA, 2011).
Hendry, A. P. et al. Evolutionary principles and their practical application. Evol. Appl. 4, 159–183 (2011).
Klemens, M. W. Amphibians and Reptiles of Connecticut and Adjacent Regions. (Connecticut Department of Environmental Protection, 1993).
Urban, M. C. Microgeographic adaptations of spotted salamander morphological defenses in response to a predaceous salamander and beetle. Oikos 119, 646–658 (2010).
U. S. Fish and Wildlife Service. National Wetlands Inventory website. U.S. Department of the Interior, Fish and Wildlife Service, Washington, D.C. http://www.fws.gov/wetlands/. (2008).
Urban, M. C. The evolution of prey body size reaction norms in diverse communities. J. Anim. Ecol. 77, 346–355 (2008).
ImageJ (U. S. National Institutes of Health, Bethesda, Maryland, 1997–2011).
Harrison, R. G. Stages and description of the normal development of the spotted salamander, Amblystoma punctatum (Linn.). 44–66 (Yale University Press, 1969).
Watson, S. & Russell, A. P. A posthatching developmental staging table for the long-toed salamander, Ambystoma macrodactylum krausei. Amphibia-Reptilia 21, 143–154 (2000).
HemiView (Burwell, Cambridge, UK, 1998–1999).
Halverson, M. A., Skelly, D. K., Kiesecker, J. M. & Freidenburg, L. K. Forest mediated light regime linked to amphibian distribution and performance. Oecologia 134, 360–364 (2003).
R Development Core Team. R: A language and environment for statistical computing. (2011).
Bolker, B. M. et al. Generalized linear mixed models: a practical guide for ecology and evolution. Trends Ecol. Evol. 24, 127–135 (2009).
Acknowledgements
This research was supported by funding from the Mianus River Gorge Preserve Research Assistantship Program, the Hixon Center for Urban Ecology, the Yale Institute for Biospheric Studies and the Connecticut Association of Wetland Scientists. I thank D. Skelly, S. Alonzo, P. Turner and M. Urban for project advice and A. Brady, S. Bolden and J. Burmeister for field assistance. J. Bushey provided chemical analysis. I am grateful to the Leopold Schepp Foundation for their support of my research through scholarship.
Author information
Authors and Affiliations
Contributions
SPB designed and carried out research, analyzed data and wrote the paper.
Ethics declarations
Competing interests
The author declares no competing financial interests.
Electronic supplementary material
Supplementary Information
Supplementary Methods and Tables
Rights and permissions
This work is licensed under a Creative Commons Attribution-NonCommercial-No Derivative Works 3.0 Unported License. To view a copy of this license, visit http://creativecommons.org/licenses/by-nc-nd/3.0/
About this article
Cite this article
Brady, S. Road to evolution? Local adaptation to road adjacency in an amphibian (Ambystoma maculatum). Sci Rep 2, 235 (2012). https://doi.org/10.1038/srep00235
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/srep00235
This article is cited by
-
Amphibian breeding phenology influences offspring size and response to a common wetland contaminant
Frontiers in Zoology (2021)
-
Factors driving the distribution of an amphibian community in stormwater ponds: a study case in the agricultural plain of Bas-Rhin, France
European Journal of Wildlife Research (2020)
-
Adaptive responses to salinity stress across multiple life stages in anuran amphibians
Frontiers in Zoology (2017)
-
Environmental exposure does not explain putative maladaptation in road-adjacent populations
Oecologia (2017)
-
Forest roadsides harbour less competitive habitats for a relict mountain plant (Pulsatilla vernalis) in lowlands
Scientific Reports (2016)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
