
Abstract
Analyses of species traits have increased our understanding of how environmental drivers such as disturbances affect the composition of arthropod communities and related processes. There are, however, few studies on which traits in the arthropod community are affected by environmental changes and which traits affect ecosystem functioning. The assembly of arthropod traits of several taxa is difficult because of the large number of species, limited availability of trait databases and differences in available traits. We sampled arthropod species data from a total of 150 managed grassland plots in three regions of Germany. These plots represent the spectrum from extensively used pastures to mown pastures to intensively managed and fertilized meadows. In this paper, we summarize information on body size, dispersal ability, feeding guild and specialization (within herbivores), feeding mode, feeding tissue (within herbivorous suckers), plant part (within herbivorous chewers), endophagous lifestyle (within herbivores), and vertical stratum use for 1,230 species of Coleoptera, Hemiptera (Heteroptera, Auchenorrhyncha), Orthoptera (Saltatoria: Ensifera, Caelifera), and Araneae, sampled by sweep-netting between 2008 and 2012. We compiled traits from various literature sources and complemented data from reliable internet sources and the authors’ experience.
Design Type(s) | observation design • time series design • species comparison design |
Measurement Type(s) | phenotype |
Technology Type(s) | phenotype characterization |
Factor Type(s) | |
Sample Characteristic(s) | Coleoptera • Hemiptera • Orthoptera • Araneae • multicellular organism • Germany • grassland |
Machine-accessible metadata file describing the reported data (ISA-Tab format)
Similar content being viewed by others


Background & Summary
Arthropod species’ traits are increasingly used in ecology to analyze the consequences of environmental change such as land-use change and intensification and climate warming1–4. This is because species respond to environmental changes depending on the traits they possess, and because species’ functions in ecosystems are mediated by their traits5. In a review on freshwater ecosystems, the functional trait approach has been highlighted as one of the most promising tools emerging for biomonitoring6. Moreover, trait-based analyses have been shown to be useful for providing deeper insights into community assembly7–9, for enabling worldwide comparisons e.g., of community responses to environmental drivers across different species assemblages10, and for predicting biological invasion11,12.
Currently, the availability of trait databases for arthropods is still limited. As a consequence, it is largely unclear which traits are favored or selected against by different environmental drivers such as human land use (response traits) and which traits affect ecosystem functioning (effect traits). Thus, the currently available trait information is likely underestimating the consequences of changes in the trait distribution within an arthropod community for ecosystem functioning13. As a consequence, a greater effort is needed to collate and provide relevant trait information.
During the last years, trait databases have been established for some arthropod groups in Europe, e.g., for carabids (carabid.org;14), cavity-nesting wasps and bees as well as European bees (Scales project; www.scales-project.net/), European hoverflies (Syrph the Net;15), and stream macroinvertebrates16. For most groups publicly available trait databases are, however, still missing. More importantly, published standardized trait values across several taxonomic groups which would enable a common trait analysis over several taxa, are very rare in the literature, at least for terrestrial invertebrates (see e.g., ref. 17).
Here, we summarize information on the nine traits body size, dispersal ability, feeding guild, feeding specialization (within herbivores), feeding mode, feeding tissue (within herbivorous suckers), plant part (within herbivorous chewers), endophyllous lifestyle (within herbivores), and vertical stratum use for a total of 1,230 species of Coleoptera, Hemiptera (Heteroptera, Auchenorrhyncha), Orthoptera (Saltatoria: Ensifera, Caelifera), and Araneae. All these species were sampled by sweep-netting on a total of 150 grassland plots across three regions in Germany between 2008 and 2012. We compiled traits from various literature sources (reference books and single publications) and complemented data from internet sources such as the spiders of Europe website (http://www.araneae.unibe.ch) and the carabids.org online database, and the authors’ experiences. The data set supports future trait-based studies in central European grasslands and similar habitats and thus has a high reuse value. It further encourages the collection of the same trait data for species from other habitats (e.g., forests) and regions for which this information is readily available. It also encourages the establishment of additional multi-taxa trait data bases and hopefully provides a framework for the collection of traits in ecosystems or regions where complete taxonomic information is currently developed (e.g., in the tropics).
Methods
Site description
The arthropod trait database was established in a project within the Biodiversity Exploratories18. The Biodiversity Exploratories cover three regions of Germany: (1) the UNESCO Biosphere Reserve Schorfheide-Chorin in the North-East (52°47′25′′-53°13′26′′ N/13°23′27′′-14°08′53′′ E, about 1,300 km2 in size, 3–140 m a.s.l.), (2) the National Park Hainich and its surrounding areas in Central Germany (50°56′14′′-51°22′43′′ N/10°10′24′′-10°46′45′′ E, about 1,300 km2, 285–550 m a.s.l.), and (3) the UNESCO Biosphere Reserve Schwäbische Alb in the Swabian Jura in the South-West (48°20′28′′-48°32′02′′ N/9°10′49′′- 09°35′54′′E, about 422 km2, 460–860 m a.s.l.). Within each of those three regions, 50 grassland plots of 50 m×50 m size are used as experimental sites. The grasslands are continuously managed by farmers by mowing, and/or grazing and/or fertilization. The overall management intensity covers the typical range of management intensity of the respective region18,19.
Field method
Arthropods were sampled by sweep netting on the 150 grassland plots across the three regions in Germany between 2008 and 2012. In June and August (July and September in Schorfheide-Chorin in 2009) all plots were sampled in all years; in additional months between May to October, samples were taken on a subset of plots (nine plots per region). On each plot and during each sampling period three transects of 50 m along the plot borders were sampled with 20 double sweeps each (one double-sweep is defined as moving the net from the left to the right and back perpendicular to the walking direction). A round sweep net with 30 cm diameter was used. Arthropods were stored in 70% ethanol in the field and sorted into taxonomic groups (Coleoptera, Hemiptera, Orthoptera, Araneae and others) in the laboratory and species were determined by taxonomic experts.
Trait data were collected for all 1,230 species of the above named target taxa sampled in all months between 2008 and 2012.
Species identification method
All species were identified by taxonomic experts of the respective taxonomic group. Specialists were either one of the authors (MMG: Heteroptera, RA: Auchenorrhyncha, TB: Araneae, FK: Coleoptera) and/or additional specialists mentioned in the acknowledgements. Taxonomy follows the classification of Freude et al. incl. Supplement volumes20 for Coleoptera, World Spider Catalogue WSC21 for Araneae, and the Fauna Europaea data base (www.faunaeur.org;22) for the other orders.
Trait assignment
Trait sources
Trait data was collected from different literature sources and completed by the authors’ previously collated databases. Trait data of Coleoptera were based on information given in taxonomic literature20,23–26 and trait databases14. Trait data of Auchenorrhyncha were based on information in taxonomic literature27,28. Trait data of Heteroptera were based on information in taxonomic literature29. Trait data of Araneae were based on information in faunistic literature30–49, and on information in the online determination key http://www.araneae.unibe.ch50. Trait data of Orthoptera were based on information given in various sources2,51–59.
Trait collection
We aimed at defining important ecological and morphological traits that can be assigned comparable values across arthropod taxonomic groups. Information for the nine traits body size, dispersal ability, feeding guild, feeding specialization (within herbivores), feeding mode, feeding tissue (within herbivorous suckers), plant part (within herbivorous chewers), endophagous lifestyle (within herbivores), and vertical stratum use was selected and derived from the sources listed above.
Trait classification
The traits of the species were classified as follows (see also Figs 1 and 2):

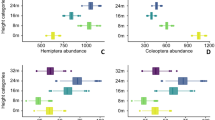
Overview on the number of species of the four orders Coleoptera, Hemiptera (Auchenorrhyncha, Heteroptera), Orthoptera (Saltatoria: Ensifera, Caelifera), and Araneae sampled over 5 years in 150 grassland plots (total: 1,230 species). For traits the number of species with each coarse classification or mm-class (body size) is given.

Overview on the number of species of the four orders Coleoptera, Hemiptera (Auchenorrhyncha, Heteroptera), Orthoptera (Saltatoria: Ensifera, Caelifera), and Araneae sampled over 5 years in 150 grassland plots (total: 1,230 species). A more detailed classification is provided for herbivorous species. The figures for feeding mode, feeding specialization, and endophagous lifestyle (note that adults of species with endophagously living larvae might also feed ectophagously) comprise all herbivores. The figure for feeding tissue comprises only sucking herbivores and the figure on feeding plant part comprises only chewing herbivores (note that species might be assigned to more than one category, e.g., many Curculionidae and Elateridae feed on roots as larvae and on leaves as adults).
(1) Body size was defined as the mean body length in mm over males and females. We standardized available information within one taxon by using information given in one standard compilation, i.e., Freude et al.20 for Coleoptera, Biedermann and Niedringhaus27 for Hemiptera: Auchenorrhyncha, Wachmann et al.29 for Hemiptera: Heteroptera, and the spiders of Europe website50 for Araneae. In Orthoptera, sizes were averaged across values given in different sources52–54.
(2) Dispersal ability, ranging from 0 to 1 by steps of 0.25, was defined differently for the groups, depending on available information. For Hemiptera and most Coleoptera, wing dimorphism between males and females was used, assigning 1 for species with fully developed wings in both sexes; 0.75 for predominantly macropterous species; 0.5 for equally brachypterous and macropterous species; 0.25 for predominantly brachypterous species and 0 for always brachypterous species. For other Coleoptera, dispersal ability was based on descriptions of flying ability20,26. Species of Araneae were assigned to the five dispersal groups taking into account activity ranges and dispersal strategies (e.g., ballooning and migration), given in different sources41–48. Species which are known to be very frequent ‘aeronauts’ and which are observed very frequently outside their main habitat were assigned 1. In contrast, class 0 would include species which never show ballooning behavior, but this did not apply to any sampled species. Species which show ballooning only over a few meter distance and were never observed outside their main habitat were assigned 0.25. When species observations outside the main habitat have been observed more frequently, species were assigned to 0.5 or 0.75 depending on the relative frequency of these observations. For Orthoptera, active dispersal ability is estimated on the basis of the size of the hind wings (alae), the occurrence of macropterous forms and studies of individual movement and colonization dynamics. We followed the classification of Reinhardt et al.59 for this trait, which takes these three criteria into account. It is largely based on the (a) wing development of the adults57, complemented by (b) results from population studies on individual movements reviewed in55,56 and long-term observations of local and regional colonization dynamics reviewed in55,56. Additionally, (c) passive dispersal behavior, e.g., by eggs attached to stipes and spread by flooding was considered51,52. All flightless species were assigned very low (0) dispersal ability and low (0.25) dispersal ability if they were using passive dispersal behavior. The other species which are able to fly were assigned to the higher dispersal abilities depending on their dispersal behavior considering criteria (b); minor individual movements and colonization dynamics was assigned medium (0.5) dispersal ability, average individual movements and colonization dynamics was assigned high (0.75) dispersal ability and high individual movements and colonization dynamics was assigned very high (1) dispersal ability.
(3) Feeding guild was classified based on the main food during the larval and during the adult stage. For example, many Heteroptera feed on plants in their larval phase but mainly on other arthropods as adults29. A coarser (herbivores, predators, fungivores, detritivores, omnivores) as well as a finer classification is given. In the finer classification, all feeding types that substantially contribute to the nutritious source during larval and adult stages of a particular species are given, with less frequent assignments given in brackets. The coarser classification was defined as follows. In Heteroptera, for example, species were classified as herbivores when (a) both nymphs and adults feed exclusively or predominantly on plants, (b) either nymphs or adults feed exclusively or predominantly on plants and plants and arthropods contribute equally to the nutrition of the other stage, or (c) either nymphs or adults feed exclusively on plants and the other stage feeds predominantly on arthropods but uses also plants as nutritious source. All species that use more than one feeding source (plants, animals, fungi, decaying plants or animals) to similar extent across larval and adult stages were classified as omnivores. Classification in Coleoptera is based on Böhme23,24, and in Orthoptera on Baur et al.53, Detzel58, and Maas et al.52 All Araneae are carnivore and all sampled Auchenorrhyncha herbivore.
(4) Feeding mode was defined as the way nutrients are ingested. We distinguished between three different modes: extraintestinal digestion, which is common in predacious spiders; chewing of plant or animal tissue by beetles and grasshoppers; sucking on plant or animal sap by Hemiptera.
(5) Feeding specialization was defined as host plant niche breadth of herbivores based on the main higher plant lineages60. We classified monophages as species feeding on one plant genus, oligophages as species feeding on one higher plant lineage (i.e., bryophytes, ferns, gymnosperms, angiosperms: monocots, angiosperms: basal eudicots, angiosperms: eurosids, angiosperms: euasterids), and polyphages as species feeding on more than one higher plant lineage. Generally, the classification is based on larval stages. Exceptions are Coleoptera species that develop in dead wood, but feed on plant pollen as adults, in which the specialization of adults was used. Classification is based on Böhme23,24, Koch61 and Rheinheimer & Hassler26 for herbivorous Coleoptera, Nickel28 for Auchenorrhyncha, Wachmann et al.29 for herbivorous Heteroptera, and Baur et al.53, Detzel58, Ingrisch & Köhler55, and Maas et al.52 for herbivorous Orthoptera.
(6) For sucking herbivores we list the feeding tissue on which the species are sucking during the larval and during the adult stage. We distinguished between xylem-, phloem-, and mesophyll-suckers and additionally assigned species that suck on reproduction organs (flowers and unripe seed on the plant), which is common in the Heteroptera families Miridae, Pentatomidae and Rhopalidae and on hard lipid-rich ripe seeds on the ground, which is common in the Heteroptera family Lygaeidae. Classification is based on Nickel28 for Auchenorrhyncha and Wachmann et al.29 for Heteroptera.
(7) For chewing herbivores we list the plant parts on which the species are feeding on during the larval and during the adult stage. We distinguished between root-feeders, stem-feeders, leaf-feeders and feeders on flowers, pollen and unripe seeds on the plant (combined to feeders of reproductive organs). Classification is based on Böhme23,24, Koch61 and Rheinheimer & Hassler26 for herbivorous Coleoptera, and Baur et al.53, Detzel58, Ingrisch & Köhler55 and Maas et al.52 for Orthoptera.
(8) Within herbivorous species we additionally classified species with an endophagous lifestyle of their larvae into gall-inducers and miners following information given in Koch61 and Rheinheimer & Hassler26 for herbivorous Coleoptera, Nickel28 and Nickel & Remane62 for Auchenorrhyncha, and Wachmann et al.29 for herbivorous Heteroptera. We additionally define the tissue were the larvae are feeding, e.g., leaf or stem miners.
(9) Vertical stratum use was selected as a trait of habitat preference and defined as the main vegetation layer (stratum) in which the species is usually observed as juvenile and adult. A coarser as well as a finer classification is given. We distinguished between soil-dwelling, ground-dwelling, herb-, shrub and/or tree-layer species, and species linked to water bodies. In the finer classification all strata which a particular species uses as habitat are given, with less frequent assignments given in brackets. In the coarser classification, all species that use more than one stratum during their life cycle were categorized as unspecific, thus ignoring different larval and adult habitat niches. For example, several Coleoptera of the family Curculionidae (e.g., genera Otiorhynchus, Phyllobius, Sitona) feed on plant roots in the soil in their larval phase but on leaves in the herb layer as adults26. Those species were hence assigned as unspecific in the coarse classification and as soil-and herb-layer species in the finer classification. Where specific information was missing (several Auchenorrhyncha & Coleoptera), the stratum was assigned from the main feeding source or the association with certain plant species. Where information on the species level was missing, information on genera or family level was used, provided that the trait value of the higher taxonomic unit was assumed to be equal for all species based on information given in literature or based on authors’ experience. Classification is based on Platen49 and Platen et al.32,36, and adapted with information in Maurer & Hänggi31, Malten33,34, Malten & Blick35, Blick38,39 and the expert’s experience for Araneae; on Böhme24, Freude et al.20, Homburg et al.14, and Rheinheimer and Hassler26 for Coleoptera; on Nickel28 for Auchenorrhyncha; on Wachmann et al.29 for Heteroptera, and on Köhler51, Baur et al.53, and Maas et al.52 for Orthoptera.
Data Records
Data set descriptors
Data set identity
Species-level data set for body size, dispersal ability, feeding guild, feeding specialization (within herbivores), feeding mode, feeding tissue (within herbivorous suckers), plant part (within herbivorous chewers), endophagous lifestyle (within herbivores), and vertical stratum use for 1,230 species of Coleoptera, Hemiptera (Heteroptera, Auchenorrhyncha), Orthoptera (Saltatoria: Ensifera, Caelifera), and Araneae.
Data set identification code
Our compiled species trait dataset (Table 1) can be found in ArthropodSpeciesTraits.txt (Data Citation 1).
Data set description
The data set comprises literature trait data of species that were sampled and measured in a project within the Biodiversity Exploratories18 which focuses on the effect of land use on arthropod community composition and related processes (e.g., species interactions such as herbivory or predation) in three regions of Germany63–65. For details on the data set see Table 2 (available online only).
Research origin descriptors
Identity
Understanding the consequences of land use in grasslands on trait composition of arthropods.
Originators
Martin M. Gossner, Nadja K. Simons, Wolfgang W. Weisser; Terrestrial Ecology Research Group, Department for Ecology and Ecosystem Management, Center for Life and Food Sciences Weihenstephan, Technische Universität München, Hans-Carl-von-Carlowitz-Platz 2, D-85354 Freising, Germany
Period of study
2008–2012.
Objectives
1. To identify important traits across taxonomic groups, 2. To detect traits that respond to land-use intensification. 3. To analyze the consequences of these trait changes for ecosystem functioning.
Technical Validation
Field methods followed a standardized protocol. A total of 60×2 sweeps×2 months×5 years=1,200 sweeps per plot is an adequate sampling effort in grasslands66. Species identification was cross-checked by different taxonomic experts and at random by barcoding67. Trait data was sent out to other experts of species ecology for cross-checks. We compiled traits from various literature sources and complemented data from reliable internet sources and the authors’ experience. If information on traits from different sources varied, experts were asked for their opinion and information of the majority is given. Body-sizes were averaged across indications given in different sources.
Additional information
Table 2 is only available in the online version of this paper.
How to cite this article: Gossner, M. M. et al. A summary of eight traits of Coleoptera, Hemiptera, Orthoptera and Araneae, occurring in grasslands in Germany. Sci. Data. 2:150013 doi: 10.1038/sdata.2015.13 (2015).
References
References
Börschig, C., Klein, A. M., von Wehrden, H. & Krauss, J. Traits of butterfly communities change from specialist to generalist characteristics with increasing land-use intensity. Basic Appl. Ecol. 14, 547–554 (2013).
Dziock, F. et al. Reproducing or dispersing? Using trait based habitat templet models to analyse Orthoptera response to flooding and land use. Agric., Ecosyst. Environ. 145, 85–94 (2011).
Moretti, M. et al. Linking traits between plants and invertebrate herbivores to track functional effects of land-use changes. J. Veg. Sci. 24, 949–962 (2013).
Vandewalle, M. et al. Functional traits as indicators of biodiversity response to land use changes across ecosystems and organisms. Biodivers. Conserv. 19, 2921–2947 (2010).
Díaz, S. et al. Functional traits, the phylogeny of function, and ecosystem service vulnerability. Ecol. Evol 3, 2958–2975 (2013).
Menezes, S., Baird, D. J. & Soares, A. M. V. M. Beyond taxonomy: a review of macroinvertebrate trait-based community descriptors as tools for freshwater biomonitoring. J. Appl. Ecol. 47, 711–719 (2010).
Van der Plas, F., Anderson, T. M. & Olff, H. Trait similarity patterns within grass and grasshopper communities: multitrophic community assembly at work. Ecology 93, 836–846 (2011).
Frenette-Dussault, C., Shipley, B. & Hingrat, Y. Linking plant and insect traits to understand multitrophic community structure in arid steppes. Funct. Ecol. 27, 786–792 (2013).
Hoiss, B., Krauss, J., Potts, S. G., Roberts, S. & Steffan-Dewenter, I. Altitude acts as an environmental filter on phylogenetic composition, traits and diversity in bee communities. Proc. R. Soc. B 279, 4447–4456 (2012).
Fountain-Jones, N. M., Baker, S. C. & Jordan, G. J. Moving beyond the guild concept: developing a practical functional trait framework for terrestrial beetles. Ecol. Entomol. 40, 1–13 (2014).
Kenis, M., Rabitsch, W., Auger-Rozenberg, M.-A. & Roques, A. How can alien species inventories and interception data help us prevent insect invasions? Bull. Entomol. Res. 97, 489–502 (2007).
Kolar, C. S. & Lodge, D. M. Progress in invasion biology: predicting invaders. Trends Ecol. Evol. 16, 199–204 (2001).
Wright, J. P. et al. Conventional functional classification schemes underestimate the relationship with ecosystem functioning. Ecol. Lett. 9, 111–120 (2006).
Homburg, K., Homburg, N., Schäfer, F., Schuldt, A. & Assmann, T. Carabids.org—a dynamic online database of ground beetle species traits (Coleoptera, Carabidae). Insect Conserv. Divers 7, 195–205 (2013).
Speight, M. C. D., Castella, E. & Sarthou, J.-P. in Syrph the Net on CD, Issue 9. The database of European Syrphidae (eds Speight M. C. D., Castella E., Sarthou J.-P., Vanappelghem C.) (Syrph the Net Publications, 2013).
Statzner, B., Bonada, N. & Dolédec, S. Conservation of taxonomic and biological trait diversity of European stream macroinvertebrate communities: a case for a collective public database. in Biodiversity and Conservation in Europe 367–390 (Springer, 2008).
Moretti, M. & Legg, C. Combining plant and animal traits to assess community functional responses to disturbance. Ecography 32, 299–309 (2009).
Fischer, M. et al. Implementing large-scale and long-term functional biodiversity research: The Biodiversity Exploratories. Basic Appl. Ecol. 11, 473–485 (2010).
Blüthgen, N. et al. A quantitative index of land-use intensity in grasslands: Integrating mowing, grazing and fertilization. Basic Appl. Ecol. 13, 207–220 (2012).
Freude, H., Harde, K. W. & Lohse, G. A. Die Käfer Mitteleuropas. Vol. 1-11 (1964–1983).
WSC. World Spider Catalog, version 15.5, http://wsc.nmbe.ch (Natural History Museum Bern, 2014).
de Jong, Y. S. D. M. Fauna Europaea version 2.6, http://www.faunaeur.org (2013).
Böhme, J . Phytophage Käfer und ihre Wirtspflanzen in Mitteleuropa—Ein Kompendium (Bioform, 2001).
Böhme, J . Die Käfer Mitteleuropas, Band K-Katalog: Faunistische Übersicht (Elsevier GmbH, Spektrum Akademischer Verlag, 2005).
Reitter, E . Die Käfer des Deutschen Reiches, Vol. Band 1-5 (K. G. Lutz Verlag, 1908–1911).
Rheinheimer, J. & Hassler, M . Die Rüsselkäfer Baden-Württembergs (Verlag Regionalkultur, 2010).
Biedermann, R. & Niedringhaus, R . The Planthoppers and Leafhoppers of Germany—Identification Key to all Species (WABV-Fründ, 2009).
Nickel, H . The leafhoppers and planthoppers of Germany (Hemiptera, Auchenorrhyncha): patterns and strategies in a highly diverse group of phytophagous insects (Pensoft, Goecke & Evers, 2003).
Wachmann, E., Melber, A. & Deckert, J . Wanzen Band 1-5 (Goecke & Evers, 2004-2012).
Růžička, V. The size groups in the spiders (Araneae) and carabids (Col. Carabidae). Acta Univ. Carol. Biol. 1982-1985, 77–107 (1985).
Maurer, R. & Hänggi, A . Katalog der schweizerischen Spinnen (Documenta Faunistica Helvetiae 12, 1990).
Platen, R., Moritz, M. & von Broen, B. Liste der Webspinnen- und Weberknechtarten (Arach.: Araneida, Opilionida) des Berliner Raumes und ihre Auswertung für Naturschutzzwecke (Rote Liste). in Rote Listen der gefährdeten Pflanzen und Tiere in Berlin, (eds Auhagen A., Platen R, Sukopp H.) Landschaftsentw. u. Umweltfr. S6, 196–205 (1991).
Malten, A. Araneae (Spinnen). in Naturwaldreservate in Hessen 5/2.1. Niddahänge östlich Rudingshain. Zoologische Untersuchungen 1990–1992. Teil 1., (eds G. Flechtner G., Dorow W. H. O., Kopelke J.-P.) Mitt. Hess. Landesforstverw. 32/1, 85–197 (1999).
Malten, A. Araneae (Spinnen). in Naturwaldreservate in Hessen 6/2.1. Schönbuche. Zoologische Untersuchungen. 1990–1992 (eds Dorow W. H. O., Flechtner G., Kopelke J.-P.). Hessen-Forst–FIV Ergeb.- u. Forschungsber. 28/1, 35–132 (2001).
Malten, A. & Blick, T. Araneae (Spinnen). in Naturwaldreservate in Hessen. Band 7/2.2. Hohestein. Zoologische Untersuchungen 1994-1996 (eds Dorow W. H. O., Kopelke J.-P.). Mitt. Hess. Landesforstverw. 42, 7–93 (2007).
Platen, R., Broen, B., Herrmann, A., Ratschker, U. M. & Sacher, P . Gesamtartenliste und Rote Liste der Webspinnen, Weberknechte und Pseudoskorpione (Araneae, Opiliones, Pseudoscorpiones) mit Angaben zur Häufigkeit und Ökologie. Natursch. Landschaftspfl. Brandenburg 8(Suppl.) 1–79 (1999).
Uetz, G. W., Halaj, J. & Cady, A. B. Guild structure of spiders in major crops. J. Arachnol. 27, 270–280 (1999).
Blick, T. Die Spinnen (Araneae) des Naturwaldreservats Goldbachs- und Ziebachsrück (Hessen). Untersuchungszeitraum 1994–1996. Mitt. Hess. Landesforstverw. 45, 57–138 (2009).
Blick, T . Die Spinnen (Araneae) des Naturwaldreservats Kinzigaue (Hessen). Untersuchungszeitraum 1999–2001. Naturwaldreserv. Hessen 12, 53–124 (2012).
Buchar, J., Růžička, V. & Merrett, P . Catalogue of Spiders of the Czech Republic (Peres Publ., 2002).
Blandenier, G. Ballooning of spiders (Araneae) in Switzerland: general results from an eleven-year survey. Bull. Br. arachnol. Soc. 14, 308–316 (2009).
Bonte, D., Baert, L., Lens, L. & Maelfait, J. P. Effects of aerial dispersal, habitat specialisation, and landscape structure on spider distribution across fragmented grey dunes. Ecography 27, 343–349 (2004).
Duffey, E. Aerial dispersal in a known spider population. J. Anim. Ecol. 25, 85–111 (1956).
Entling, M. H., Stämpfli, K. & Ovaskainen, O. Increased propensity for aerial dispersal in disturbed habitats due to intraspecific variation and species turnover. Oikos 120, 1099–1109 (2011).
Rensch, M., Volkmar, C. & Spilke, J. Aerial dispersal of spiders in middle-east Germany. Mitt. dt. Ges. allg. angew. Ent 17, 41–44 (2009).
Richter, C. J. Aerial dispersal in relation to habitat in eight wolf spider species (Pardosa, Araneae, Lycosidae). Oecologia 5, 200–214 (1970).
Thomas, C. & Jepson, P. Differential aerial dispersal of linyphiid spiders from a grass and a cereal field. J. Arachnol. 27, 294–300 (1999).
Vanden Borre, J., Bonte, D. & Maelfait, J.-P. Aeronautisch gedrag ('ballooning') bij spinnen (Araneae). Nwsbr. Belg. Arachnol. Ver. 19, 1–24 (2004).
Platen, R. Ökologie, Faunistik und Gefährdungssituation der Spinnen (Araneae) und Weberknechte (Opiliones) in Berlin (West) mit dem Vorschlag einer roten Liste. Zool. Beitr. N.F 28, 445–487 (1984).
Nentwig, W., Blick, T., Gloor, D., Hänggi, A. & Kropf, C. Araneae—Spiders of Europe, http://www.araneae.unibe.ch/ (2014).
Köhler, G . Fauna der Heuschrecken (Ensifera et Caelifera) des Freistaates Thüringen (Thüringer Landesanstalt für Umwelt und Geologie, 2001).
Maas, S., Detzel, P. & Staudt, A . Gefährdungsanalyse der Heuschrecken Deutschlands: Verbreitungsatlas, Gefährdungseinstufung und Schutzkonzepte (Bundesamt für Naturschutz, 2002).
Baur, B., Baur, H., Roesti, C. & Roesti, D . Die Heuschrecken der Schweiz (Haupt Verlag, 2006).
Bellmann, H . Heuschrecken: beobachten, bestimmen Vol. 2 (Naturbuch Verlag, 1993).
Ingrisch, S. & Köhler, G . Die Heuschrecken Mitteleuropas (Westarp Wissenschaften, 1998).
Köhler, G . Ökologische Grundlagen von Aussterbeprozessen: Fallstudien an Heuschrecken (Caelifera et Ensifera) (Laurenti, 1999).
Bruckhaus, A. & Detzel, P. Erfassung und Bewertung von Heuschrecken-Populationen. Ein Beitrag zur Objektivierung des Instruments der Roten Listen. Nat.schutz Landsch.plan 29, 19–33 (1997).
Detzel, P . Die Heuschrecken Baden-Württembergs (Verlag Eugen Ulmer, 1998).
Reinhardt, K., Köhler, G., Maas, S. & Detzel, P. Low dispersal ability and habitat specificity promote extinctions in rare but not in widespread species: the Orthoptera of Germany. Ecography 28, 593–602 (2005).
The Angiosperm Phylogeny G. An update of the Angiosperm Phylogeny Group classification for the orders and families of flowering plants: APG III. Bot. J. Linn. Soc. 161, 105–121 (2009).
Koch, K . Die Käfer Mitteleuropas. Ökologie, Band 1-3 (Goecke & Evers, 1989–1992).
Nickel, H. & Remane, R. Artenliste der Zikaden Deutschlands, mit Angaben zu Nährpflanzen, Lebenszyklen und Verbreitung (Hemiptera, Fulgoromorpha et Cicadomorpha). Beitr. Zikadenkunde 5, 27–64 (2002).
Gossner, M. M., Weisser, W. W. & Meyer, S. T. Invertebrate herbivory decreases along a gradient of increasing land-use intensity in German grasslands. Basic Appl. Ecol. 15, 347–352 (2014).
Simons, N. K. et al. Resource-mediated indirect effects of grassland management on arthropod diversity. PLoS ONE 9, e107033 (2014).
Simons, N. K. et al. Effects of land-use intensity on arthropod species abundance distributions in grasslands. J. Anim. Ecol. 84, 143–154 (2015).
Southwood, T. R. E. & Henderson, P. A . Ecological methods. 3rd edn (Blackwell Science Ltd, 2000).
Raupach, M. J. et al. Building-up of a DNA barcode library for true bugs (Insecta: Hemiptera: Heteroptera) of Germany reveals taxonomic uncertainties and surprises. PLoS ONE 9, e106940 (2014).
Data Citations
Gossner, M. M. Dryad (2015) http://dx.doi.org/10.5061/dryad.53ds2
Acknowledgements
We thank Markus Lange, Manfred Türke, Esther Pašalić, Ellen Sperr, Louis Sikora, Steffen Both and Norbert Leber for their help with sweep-netting and Boris Büche, Michael-Andreas Fritze and Thomas Wagner (Coleoptera), Anne Kästner and Oliver Wiche (Auchenorrhyncha), Günter Köhler (Orthoptera), Franz Schmolke (Heteroptera), for identifying the arthropods and fruitful discussions. We thank the managers of the three Exploratories, Kirsten Reichel-Jung, Swen Renner, Katrin Hartwich, Sonja Gockel, Kerstin Wiesner, and Martin Gorke for their work in maintaining the plot and project infrastructure; Christiane Fischer and Simone Pfeiffer for giving support through the central office, Michael Owonibi for managing the central data base, and Markus Fischer, Eduard Linsenmair, Dominik Hessenmöller, Jens Nieschulze, Daniel Prati, Ingo Schöning, François Buscot, Ernst-Detlef Schulze and the late Elisabeth Kalko for their role in setting up the Biodiversity Exploratories project. The work has been funded by the DFG Priority Program 1374 ‘Infrastructure-Biodiversity-Exploratories’ (DFG-WE 3081/21-1.). Field work permits were issued by the responsible state environmental offices of Baden-Württemberg, Thüringen, and Brandenburg (according to § 72 BbgNatSchG).
Author information
Authors and Affiliations
Contributions
M.M.G. conceived the idea for the manuscript, defined the final outline of the manuscript, wrote the first manuscript draft, collected trait information, and provided a trait database of Heteroptera. N.K.S. conceived the idea for the manuscript, collected trait information, and commented on all manuscript versions. R.A. provided a trait database of Auchenorrhyncha and commented on the manuscript. T.B. provided a trait database of Araneae and commented on the manuscript W.H.O.D. provided a trait database of Heteroptera and commented on the manuscript. F.D. provided a trait database of Orthoptera and commented on the manuscript. F.K. provided a trait database of Coleoptera. W.R. provided a trait database of Heteroptera and commented on the manuscript. W.W.W. conceived the idea for the manuscript and commented on all manuscript versions.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing financial interests.
ISA-Tab metadata
Rights and permissions
This work is licensed under a Creative Commons Attribution 4.0 International License. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in the credit line; if the material is not included under the Creative Commons license, users will need to obtain permission from the license holder to reproduce the material. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0 Metadata associated with this Data Descriptor is available at http://www.nature.com/sdata/ and is released under the CC0 waiver to maximize reuse.
About this article

Cite this article
Gossner, M., Simons, N., Achtziger, R. et al. A summary of eight traits of Coleoptera, Hemiptera, Orthoptera and Araneae, occurring in grasslands in Germany. Sci Data 2, 150013 (2015). https://doi.org/10.1038/sdata.2015.13
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/sdata.2015.13
This article is cited by
-
Insect decline in forests depends on species’ traits and may be mitigated by management
Communications Biology (2023)
-
Forest gaps increase true bug diversity by recruiting open land species
Oecologia (2023)
-
Increased arthropod biomass, abundance and species richness in an agricultural landscape after 32 years
Journal of Insect Conservation (2023)
-
Present and historical landscape structure shapes current species richness in Central European grasslands
Landscape Ecology (2022)
-
Urbanization alters the abundance and composition of predator communities and leads to aphid outbreaks on urban trees
Urban Ecosystems (2021)
