
Abstract
Oxygen is a key regulatory factor of anaerobic ammonium oxidation (anammox). Although the inhibitory effect of oxygen is evident, a wide range of oxygen sensitivities of anammox bacteria have been reported so far, which makes it difficult to model the marine nitrogen loss and design anammox-based technologies. Here, oxygen tolerance and detoxification mechanisms of four genera of anammox bacteria; one marine species (“Ca. Scalindua sp.”) and four freshwater anammox species (“Ca. Brocadia sinica”, “Ca. Brocadia sapporoensis”, “Ca. Jettenia caeni”, and “Ca. Kuenenia stuttgartiensis”) were determined and then related to the activities of anti-oxidative enzymes. Highly enriched planktonic anammox cells were exposed to various levels of oxygen, and oxygen inhibition kinetics (50% inhibitory concentration (IC50) and upper O2 limits (DOmax) of anammox activity) were quantitatively determined. A marine anammox species, “Ca. Scalindua sp.”, exhibited much higher oxygen tolerance capability (IC50 = 18.0 µM and DOmax = 51.6 µM) than freshwater species (IC50 = 2.7–4.2 µM and DOmax = 10.9–26.6 µM). The upper DO limit of “Ca. Scalindua sp.” was much higher than the values reported so far (~20 µM). Furthermore, the oxygen inhibition was reversible even after exposed to ambient air for 12–24 h. The comparative genome analysis confirmed that all anammox species commonly possess the genes considered to function for reduction of O2, superoxide anion (O2•-), and H2O2. However, the superoxide reductase (Sor)-peroxidase dependent detoxification system alone may not be sufficient for cell survival under microaerobic conditions. Despite the fact that anaerobes normally possess no or little superoxide dismutase (Sod) or catalase (Cat), only Scalindua exhibited high Sod activity of 22.6 ± 1.9 U/mg-protein with moderate Cat activity of 1.6 ± 0.7 U/mg-protein, which was consistent with the genome sequence analysis. This Sod-Cat dependent detoxification system could be responsible for the higher O2 tolerance of Scalindua than other freshwater anammox species lacking the Sod activity.
Similar content being viewed by others

Introduction
Oxygen plays a key role in regulating the marine nitrogen cycle even in oxygen minimum zones (OMZs). The OMZs constitute only 0.1% of total ocean volume (if the OMZ is defined as O2≦5 µM) but account for 20–40% of the oceanic nitrogen loss through denitrification and anaerobic ammonium oxidation (anammox) [1]. Such nitrogen loss (i.e., N2 production) intensively occurs at the oxic and anoxic interface of the OMZs [2, 3]. This indicates the importance of close microbial interactions among aerobic nitrification, denitrification, and anammox (anaerobically converts NH4+ to N2 with NO2−). The effects of oxygen levels on these processes are of central importance for understanding their contributions to the marine nitrogen cycle. Especially, the estimated contribution of anammox may vary greatly depending on the oxygen tolerance capability of marine anammox bacteria which basically determines the OMZ water volume with active anammox activity [4, 5]. Thus, their oxygen sensitivity and dynamic response to oxygen exposure need to be experimentally quantified.
A wide range of oxygen sensitivities of anammox bacteria have been reported for freshwater anammox species, whereas no data were available for marine species “Ca. Scalindua” so far [6, 7]. The previous O2 amendment studies using oceanic OMZ samples have shown that anammox activities were 50% inhibited at 0.9–16 µM [4, 8,9,10,11]. Similar or more severe oxygen sensitivity could be found for some freshwater anammox species like B. anammoxidans, Brocadia sp. and K. stuttgartiensis, demonstrating that anammox activities were completely inhibited at low dissolved oxygen (DO) levels (1.25–3.75 µM) [6, 12,13,14]. However, the anammox activities could be gradually recovered after oxygen inhibition, depending on the perceived inhibition levels [12,13,14].
A reported wide range of oxygen sensitivity could be attributed to many factors such as anammox species, degree of enrichment, form of biomass (planktonic or aggregated biomass), and measurement methods of anammox activity (e.g., NH4+ and/or NO2− consumption or 14+15N2 gas production). In order to assess the reported aerotolerant natures more prudently, the anammox biomass used in those studies need to be clearly specified in detail since oxygen could be consumed by coexisting aerobes [4, 6, 13, 15] or aerobes shield anammox consortia from oxygen exposure in microbial aggregates, biofilms, or marine snow, both of which could result in overestimation of oxygen tolerance [16]. Therefore, aggregated or flocculated biomass often exhibited higher oxygen tolerance than planktonic biomass [17, 18]. In addition, there could be truly inter-species differences in oxygen sensitivity as suggested earlier by Yan et al. [19], which, however, has not been experimentally confirmed yet. Inherent oxygen inhibition kinetics (e.g., 50% inhibitory concentration (IC50) and upper O2 limits (DOmax)) and recovery from O2 inhibition of anammox bacteria have not been directly determined using well-defined laboratory enrichment cultures to date.
Furthermore, oxygen detoxification mechanisms are largely unknown for anammox bacteria. It is generally known that when molecular oxygen (O2) diffuses into cells, reactive oxygen species (ROSs), such as superoxide anion (O2 ∙ -) and hydrogen peroxide (H2O2) are generated as oxygen reduction by-products. Furthermore, O2 ∙ - can react with H2O2 to generate hydroxyl radicals (∙OH) in cells, which are the most potent oxidants among ROSs and thus cause the damage on DNA and proteins. Thus, organisms need to detoxify these ROSs by anti-oxidative enzymes such as superoxide dismutase (Sod), catalase (Cat), and peroxidases to survive in the presence of oxygen [20]. However, it is widely known that strict anaerobes usually do not possess Sod and/or Cat because these enzymes generate O2, propagating the further production of ROS [21]. Since anammox bacteria are regarded as obligate anaerobic bacteria [22], it is vital to investigate their ROS detoxification mechanism if they possess. It was speculated that “Ca. Brocadia sp.” most likely utilize a Sod-cytochrome c peroxidase system to detoxify ROS based on metagenomic and metatranscriptomic analyses [23]. However, abundance of “Ca. Brocadia sp.” in the biomass used in their study was relatively low (< 50%), and the activities of anti-oxidative enzymes have never been experimentally determined.
In the present study, oxygen tolerance capability of one marine species (“Ca. Scalindua sp.”) and four freshwater anammox species (“Ca. Brocadia sinica”, “Ca. Brocadia sapporoensis”, “Ca. Jettenia caeni”, and “Ca. Kuenenia stuttgartiensis”) were determined and then related to the activities of anti-oxidative enzymes (Sod, Cat, and peroxides). Free-living planktonic enrichment anammox cultures were further purified (> 99.8%) by applying Percoll density gradient centrifugation [24] and then subjected to these studies. The experimental results revealed that a marine anammox species, “Ca. Scalindua sp.”, exhibited the highest oxygen tolerance, which is likely attributed to the higher Sod activity. To the best of our knowledge, this is the first time to quantitatively verify that marine anammox bacteria possess higher oxygen tolerance than freshwater species and its oxygen detoxification mechanisms.
Materials and methods
Anammox biomass
Free-living planktonic cells of “Ca. Brocadia sinica”, “Ca. Brocadia sapporoensis”, “Ca. Jettenia caeni”, “Ca. Kuenenia stuttgartiensis” and “Ca. Scalindua sp.” were obtained from respective stock membrane bioreactors (MBRs) that have been maintained in our lab [7, 25,26,27,28,29]. The MBRs of “Ca. B. sapporoensis”, “Ca. Scalindua sp.” and “Ca. K. stuttgartiensis” were maintained at 25 °C, while the MBRs of “Ca. B. sinica” and “Ca. J. caeni” were maintained at 37 °C. The average purities of these MBR anammox cultures were 95 ± 3% of total bacteria, which were determined by FISH and q-PCR analysis [29,30,31]. Based on the measurements of 16 S rRNA gene copy numbers by qPCR, “Ca. B. sinica”, “Ca. K. stuttgartiensis”, “Ca. B. sapporoensis”, “Ca. J. caeni” and “Ca. Scalindua sp.” accounted for more than 99% of the anammox population in respective anammox MBR cultures. The MBR biomass was continuously stirred with magnetic stirrer to make sure the cells are planktonic state. The growth medium was made anoxically by flushing nitrogen gas (>99.995%) for at least 3 h, and an inert gas mixture (95% Argon and 5% CO2) was continuously bubbled into the bottom of the MBRs in order to maintain anaerobic conditions. The feeding medium contained the following compositions (mg L−1): FeSO4·7H2O (9.0), EDTA·4Na (5.0), NaCl (1.0), KCl (1.4), CaCl2·2H2O (1.4), MgSO4·7H2O (1.0), NaHCO3 (84.0), KH2PO4 (54.0), and trace element solution I and II (0.5 mL each) for all five MBRs [32]. For “Ca. Scalindua sp.” and “Ca. K. stuttgartiensis” cultures, the salinity of the medium was adjusted to 2.5% and 0.5% respectively by addition of an artificial sea salt (SEALIFE, Marine Tech, Tokyo, Japan). The pH of MBR cultures were in the range 7.3 to 7.5 without any pH adjustment. The nitrogen loading rate (NLR) was carefully controlled to avoid nitrite inhibition.
Biomass preparation
One night before the start-up of batch incubation experiments, planktonic anammox biomass was collected from respective MBRs, harvested by centrifugation under anoxic conditions (20°C, ×10000 rpm, 6 min), and washed with anoxic inorganic medium without ammonium and nitrite (purged by N2 gas for 1 h) for three times in an anaerobic chamber (Coy Laboratory Products, Michigan, USA). To further purify the MBR anammox cultures, a density gradient separation was performed using Percoll (Cytiva, Tokyo, Japan) as described previously [24]. The Percoll-purified biomass was then anoxically washed twice with inorganic nutrient medium (purged by N2 gas for 1 h) without NH4+ and NO2− in the anaerobic chamber, in which anammox bacteria accounted for more than 98% of the total biomass based on FISH analysis. The Percoll-purified biomass was anoxically kept in a 37 °C incubator or room temperature (ca. 25 °C) for overnight before oxygen inhibition or reduction experiments. Percoll purification has been confirmed to have no effect on anammox activity.
Oxygen inhibition experiments
To determine the inhibitory effect of DO concentrations (µmol L−1) on specific anammox activity (SAA), 15N-labelling batch incubation experiments were performed at varying O2 concentrations (%, v/v) in 20 mL headspace of 70 mL serum vials containing 50 mL of the inorganic medium. The experimental procedure of 15N-labelling batch incubation experiments has been described in detail elsewhere [26]. Briefly, the inorganic medium containing 3 mM (14NH4)2SO4 and 3 mM Na15NO2 was firstly purged with N2 gas for at least 30 min. Each vial was then sealed with a butyl-rubber stopper and aluminum cap and then autoclaved one night before the start-up of batch incubation. The headspace gas of vials was exchanged by applying 3 cycles of vacuuming (2 min) and purging (1 min at 1.5 atm) with highly pure helium gas (99.9999% He) by a gas exchange machine (Model IP-8, SANSHIN, Yokohama, Japan) to remove the residual air (oxygen) and to enhance the detection sensitivity of 14+15N2 produced by anammox bacteria. The initial gas pressure of the headspace was set at ca. 1.5 atm. A magnetic stir bar was placed inside the vial to gently stir the medium during batch incubation.
To determine the effect of O2 concentrations on SAA, varying volumes of ambient air were injected into the headspace of sealed vials using a gas tight glass syringe (VICI, Baton Rouge, LA, USA) at least 4 h before the inoculation of biomass. The headspace O2 concentrations varied from 0 to 3.5% O2 (v/v), corresponding to theoretical saturated DO concentrations in the culture medium ranging from 0 to ca. 65 µM and 0 to ca. 53 µM at 25 °C and 37 °C (at 1.5 atm), respectively. To convert the headspace O2 concentrations to DO concentrations in the medium, the standard curves of the measured DO concentrations vs. the headspace O2 concentrations (%, v/v) were constructed in advance (Fig. S1). The DO concentrations in the medium (µM) were measured by a microsensor (Unisense oxygen needle sensor OX-N 13621) and the Winkler method and plotted against the headspace O2 concentrations (%) (Fig. S1). The measured DO concentrations were well correlated with the theoretical O2 concentrations at 1.5 atm for both temperatures (25 and 37 °C). The DO concentrations in the following O2 inhibition and reduction experiments were determined using the regression lines for microsensor’s data.
For O2 inhibition experiments, 1 mL of Percoll-purified biomass suspension was inoculated into the medium in sealed vials to obtain a final biomass concentration of 0.1–0.15 mg-protein mL−1. Immediately after biomass inoculation, the air sample (at 0 h) was collected from the headspace, and 14+15N2 concentrations were measured by a gas chromatography and mass spectrometry (GC-MS, SHIMADZU). The vials with “Ca. B. sinica” and “Ca. J. caeni” were incubated at 37 °C in an incubator, and the other anammox species were incubated at room temperature (ca. 25 °C) in dark. The 14+15N2 concentrations were measured hourly, and the maximum specific anammox activity (MSAA, expressed as µmole 14+15N2 mg-protein−1 h−1) were calculated by dividing the slope of linear regression of initial 14+15N2-production as a function of time by the amount of biomass (expressed as protein) in the vial at the beginning of the assay. The lag-phase was considered in the determination of 14+15N2-production rates. The 15N-labelling batch incubation experiments were performed at varying O2 concentrations at least triplicate for each species.
The percentages of the MASS (relative MASS) that were maintained after exposure to varying O2 concentrations were calculated with respect to the average MASS of nonexposed samples (anoxic cultures) as follows:
where MSAA and MSAAanoxic is the average of MSAA of O2-exposed samples and nonexposed samples (anoxic cultures) (µmole 14+15N2 mg-protein−1 h−1), respectively.
Oxygen inhibition model
An oxygen inhibition model with two parameters (IC50: the DO concentration that causes 50% inhibition of MSAA (µM) and DOmax: the DO concentration above which anammox activity is completely inhibited (µM)) were used to quantitatively evaluate the effect of DO concentration on anammox activity [33].
where DO is dissolved oxygen concentration in culture medium (µmole L−1). This model was used because the upper limits of DO have been previously reported for marine environments [10, 11] and engineered systems [6]. Values for IC50 and DOmax were obtained by curve fitting the data.
Oxygen reduction experiments
The selected anammox species (“Ca. B. sinica”, “Ca. K. stuttgartiensis” and “Ca. Scalindua sp.”) were transferred into 30 mL of the inorganic medium in 100 mL serum vials containing 70 mL headspace. The residual DO was removed and the headspace was exchanged with highly pure helium gas (99.9999% He) at 1.5 atm as described above. O2 gas (99%, 200 µL) was injected into the headspace with a gas tight syringe 3 h before the addition of 15NH4Cl and Na14NO2 (final concentration of 3 mM each). Thereafter, the vials were incubated at room temperature (ca. 25 °C). The air samples were taken from the headspace over the course of the incubation time, and 14+15N2 and O2 concentrations were measured by a gas chromatography and mass spectrometry (GC-MS, SHIMADZU). The O2 concentrations (%) were converted to DO concentration using the standard curves of the measured DO concentrations vs. the headspace O2 concentrations (%, v/v) (Fig. S1) as described above. The O2 reduction rates were determined by dividing the slope of linear regression of DO concentration as a function of time by the protein concentration and are expressed as µmole DO per g-protein per hour.
Recovery after oxygen exposures
Anammox biomass was exposed to varying headspace O2 concentrations (0 (anoxic), 0.7, 1.4, 2.8, and 21% (v/v)) for 12 h and 24 h in the absence of NH4+ and NO2− as described above. The 21% headspace O2 was achieved by unsealing the vials (exposure to ambient air). Unexposed vials (anoxic) were also prepared as a positive control. The oxygen recovery assays were performed in 12 ml vials with a liquid volume of 6 mL at least triplicate biological samples.
After exposure to oxygen for 12 h and 24 h, the headspace gas was exchanged by applying 3 cycles of vacuuming (2 min) and purging (1 min at 1.5 atm) with highly pure helium gas (99.9999% He) by a gas exchange machine (Model IP-8, SANSHIN, Yokohama, Japan). Thereafter, anoxically prepared (N2 gas purged) stock solution of (14NH4)2SO4 and Na15NO2 were supplemented to obtain a final concentration of 3 mM each. 14+15N2 gas production was monitored up to 8–10 h, which was defined as “immediate recovery”. The vials were also continuously incubated for 24 h under anoxic conditions, and then the same amounts of substrates were supplied again (a final concentration of (14NH4)2SO4 and Na15NO2, 3 mM each). The 14+15N2 gas production was monitored from 24 h to 32 h as described above, which was defined as “secondary recovery”.
Comparative genome analysis
Comparative genome analysis was carried out to study the types and distributions of genes encoding anti-oxidative enzymes among the anammox species studied. Presence/absence of the genes were examined by performing a blastp search (threshold e-value of blastp search 10−15) with 31 anammox bacterial genomes affiliated into 18 bacterial species in the bacterial order Brocadiales (Table S1). The 18 bacterial species cover all the bacterial species in the order Brocadiales defied in the GTDB database (Release RS95) [34]. The genome sequences include those obtained from the enrichment culture of “Ca. B. sinica”, “Ca. B. sapporoensis”, “Ca. J. caeni”, “Ca. K. stuttgartiensis” and “Ca. Scalindua sp.” used in the present study [35]. Multiple sequence alignment used the ClustalW 1.83 (Gap opening and extension penalties in a pairwise alignment; 10 and 0.05, respectively) [36], and visualized using ESPrint 3.0 [37]. A phylogenetic tree was calculated in MEGA 11.0.8 using maximum likelihood (Jones-Taylor-Thornton model) and neighbor joining methods (Poisson model) [38].
Anti-oxidative enzyme activity assays
Planktonic anammox cells were collected from respective stock MBRs and incubated in 70 mL vials after Percoll density gradient centrifugation. During incubation, anammox cells were subjected to two patterns of oxygen exposure: (i) exposure to different O2 concentrations (0, 0.7, 1.4, and 2.8% (v/v)) for 12 h; or (ii) exposure to air-saturating DO concentrations (ambient air) for different periods of time (0, 0.5, 1 and 2 h). After each oxygen exposure, biomass was harvested by centrifugation (4 °C, 10,000 rpm, 6 min) and concentrated 25-fold by resuspending in an ice-cold potassium phosphate buffer (50 mM, pH 7.4). To prevent protein degradation, 0.1 mM phenylmethylsulfonyl fluorid (PMSF) was added as a protease inhibitor. The cell suspension was disrupted by passing thrice through a French pressure cell press unit (AVESTIN, ON, Canada) at homogenizing pressure of 1200 MPa. Cell debris was removed by centrifugation (4 °C, 4500 rpm, 60 min). The resulting supernatant fraction was transferred to 15 mL sterile tubes and stored in an ice-cold bath. All cell-free extracts were tested within a day. At least, three independent determinations were performed for each enzyme activity in all conditions. All anti-oxidative enzyme activities were expressed as units of enzyme activity per milligram of protein of the cell-free extract.
Superoxide dismutase (Sod) activity was assayed spectrophotometrically at 560 nm by measuring its ability to inhibit the photochemical reduction of nitroblue tetrazolium (NBT) (superoxide anion O2•- reacts with NBT and reduces the yellow tetrazolium to a blue precipitate) according to the protocol previously described [39]. Since the assays were conducted in microplates, the reaction mixture contained in 200 µL: 50 mM potassium phosphate buffer (pH 7.8), 10 mM L-methionine, 60 µM NBT, 8 µM EDTA, 1.6 µM riboflavin, and 22 µL of cell-free extract (0.05–0.06 mg-protein). Riboflavin was added at the end, and the tubes were mixed by shaking [40]. The microplate was illuminated by a 4000 Lux cool LED lightboard (AXEL, Tokyo, Japan) for 20 min, and the absorbance at 560 nm was recorded at 2, 4, 6, 8, 10, 15, and 20 min in a multilabel plate counter (PerkinElmer, Waltham, USA). The samples containing the same components were placed in dark, which were used as a blank reference. One activity unit (1 U) of Sod was defined as the amount of enzyme required to inhibit the rate of riboflavin and illumination dependent NBT reduction by 50% at 25 °C, pH 7.8, and a light intensity of 4000 Lux [40].
Catalase (Cat) activity was assayed spectrophotometrically at 240 nm by measuring its ability to decompose H2O2 as described elsewhere [41]. The reaction mixture contained in 1 mL: 50 mM potassium phosphate buffer (pH 7.0), 10 mM H2O2, and 0.1 mL of cell-free extracts (0.2–0.3 mg-protein). The reaction was initiated by adding H2O2 into a quartz cuvette containing phosphate buffer and enzyme extracts. One activity unit (1 U) of Cat was defied as the amount of enzyme required to catalase the decomposition of 1 µmol H2O2 in 1 min at 25 °C and pH 7.0.
Cytochrome c peroxidase (Ccp) activity was assayed spectrophotometrically at 550 nm by monitoring the loss of reduced cytochrome c according to the protocol provided by Munkres et al. [42]. The reaction mixture contained in 1 mL: 50 mM acetate buffer (pH 6.0), 1 mM EDTA, 1 mM sodium azide, 20 µL of a reduced cytochrome c solution (1.2 mg mL−1), 180 µM H2O2, and 0.1 mL of cell-free extracts (0.2–0.3 mg-protein). The net rate of absorbance change was obtained by subtracting the rate of a control without cell-free extract. One activity unit (1 U) of Ccp was defended as the amount of enzyme required to catalase the oxidation of 1 µmol cytochrome c in 1 min at 25 °C and pH 6.0.
Glutathione peroxidase (Gpx) activity was assayed spectrophotometrically at 340 nm by monitoring the loss of NADPH according to the protocol provided by Drotar et al., [43]. The reaction mixture contained in 1 mL: 50 mM potassium phosphate buffer (pH 7.0), 2 mM EDTA, 2 mM reduced glutathione, 0.1 mM NADPH, 2.5 units of glutathione reductase, 0.09 mM H2O2, and 0.1 mL of cell-free extracts (0.2–0.3 mg-protein). The net rate of absorbance change was obtained by subtracting the rate of a control without cell-free extract. One activity unit (1 U) of Gpx was defined as the amount of enzyme required to catalase the reduction of 1 µmol NADPH in 1 min at 25 °C and pH 7.0.
Chemical analyses
For determination of 14+15N2 gas and O2 concentrations, 50 μL of headspace gas was collected from the tested vial with a gas tight syringe (VICI, Baton Rouge, LA, USA) and injected into a gas chromatography mass spectroscopy (GCMS-QP2010SE, Shimadzu, Japan) equipped with a CP-Pora Bond Q fused silica capillary column (Agilent Technologies, Santa Clara, CA, USA) as described previously [25]. The specific anammox activity (SAA) was calculated by dividing the 14+15N2 gas production rate by biomass (protein) concentration. In each experiment, SAA tests were performed in triplicate at least.
Concentrations of DO were measured by a Unisense oxygen needle sensor (OX-N 13621, Aarhus, Denmark). The oxygen sensor was calibrated using a two-point calibration (zero and a saturation point). The deionized water was purged with pure N2 gas for at least 1 h or dissolving 1 g of sodium sulfite (Na2SO3) to obtain a zero-DO solution and bubbled with air for at least 1 h to achieve the saturated oxygen concentration at 25 °C and 37 °C, respectively.
Biomass concentration was determined as protein concentration with the DC Protein Assay Kit (Bio-Rad Laboratories, Munich, Germany) using the bovine serum albumin (BSA) as the protein standard. In brief, 1 mL of cell suspension was collected, centrifuged at 12,000 rpm for 15 min, resuspended in the same amount of 10% (w/v) sodium dodecyl sulfate (SDS) solution and incubated for 30 min at 99 °C (Note this procedure is unnecessary for the protein measurement of cell-free extract in enzyme activity tests). Then, 5 µL of the cell suspension was incubated with Bio-Rad protein assay reagent in microplate for 15 min at 25 °C. The absorbance was measured at 750 nm.
Results and Discussion
Effect of O2 concentration on specific anammox activity (SAA)
It was confirmed that the Percoll-purified anammox biomass were highly enriched and well dispersed (Fig. 1). The high purity (> 98%) was also confirmed for all anammox species by FISH and the measurements of 16 S rRNA gene copy numbers by qPCR.

Cells were hybridized with combination of FITC-labeled EUB338 mix probe (most of the Eubacteria) and an anammox species specific probe; Alexa555-labeled AMX156 probe for “Ca. B. sinica” (A) and “Ca. B. sapporoensis” (B), TRITC-labeled JEC152 probe for “Ca. J. caeni” (C), CY3-labeled KST157 probe for “Ca. K. stuttgartiensis” (D), and TRITC-labeled Scal1129b probe for “Ca. Scalindua sp.” (E), respectively. Anammox bacteria were shown in orange yellow (green+red), whereas other bacteria were shown in green. All FISH images show highly enriched and well dispersed cells. Scale bars represent 20 µm.
The Percoll-purified planktonic anammox cells were cultured with 14NH4+ and 15NO2− in sealed vials with headspace containing different O2 concentrations (up to 3.32%, corresponding to ca. 60 µM and 50 µM dissolved O2 (DO) at 25 °C and 37°C at 1.5 atm, respectively), and then 14+15N2 production rates were measured for five anammox species (Fig. 2). DO concentrations in the vials gradually decreased during the batch incubations for all batch experiments (Fig. S2). The onset of 14+15N2 gas production was gradually delayed (lag-time), and the 14+15N2 gas production rates decreased along with the increase of O2 concentration for 4 freshwater anammox species (Fig. 2). However, a marine species “Ca. Scalindua sp.” could steady produce 14+15N2 up to 1.65% O2 (DO = ca. 30 µM) without any lag time.

“Ca. B. sinica” (A), “Ca. B. sapporoensis” (B), “Ca. J. caeni” (C), “Ca. K. stuttgartiensis” (D), and “Ca. Scalindua sp.” (E). Batch incubation experiments with 15NO2− (3 mM) and 14NH4+ (3 mM) were performed at varying headspace O2 concentrations for each species at least triplicate. One of representative data sets is presented here.
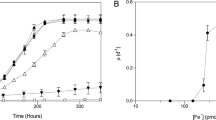
The effect of O2 concentration on the MSAA, as indicated by relative MSAA (%), was evaluated (Fig. 3). The relative MSAA dramatically decreased with increasing O2 concentrations, approaching zero asymptotically, for freshwater anammox species. No significant anammox activity was virtually detected at >1.0% O2 (corresponding to ca.15.2 µM and 18 µM DO at 25 °C and 37 °C, respectively) for freshwater anammox species. In contrast, the MSAA of “Ca. Scalindua sp.” could be detected up to 2.0% O2 (36 µM DO), showing the least O2 sensitivity among the anammox species examined in the present study.

“Ca. B. sinica” (A), “Ca. B. sapporoensis” (B), “Ca. J. caeni” (C), “Ca. K. stuttgartiensis” (D), and “Ca. Scalindua sp.” (E). The MSAA (µmole 14+15N2 mg-protein−1 h−1) was calculated from maximum 14+15N2 production rate and amount of biomass in the vial and given as a percentage of MSAA observed at DO = 0 µM (defined as 100%). The red lines represent DO inhibition kinetics as shown in Eq. (2) fitted to the data. Inserted figures show the effect of DO concentrations on lag times of 14+15N2 gas production after exposures to O2. The lag time means the time needed to reduce DO concentration to tolerable levels and to repair and reactivate damaged enzyme systems. DO concentrations were calculated based on the standard curves of the measured DO concentration (µM) vs. the headspace O2 concentration (%) at 25 °C and 37 °C at 1.5 atm, respectively (Fig. S1, see Materials and Methods). Error bars represent the standard deviations of three replicate samples.
The DO concentrations that cause 50% inhibition of MSAA (IC50) and the maximum DO concentrations above which anammox is completely inhibited (DOmax) were estimated for each anammox species based on the inhibition model Eq. (2). All freshwater anammox species showed similar responses to increased O2 (high sensitivity). The IC50 values were in the range 2.7 - 4.2 µM, and DOmax values were in the range 10.9 - 26.6 µM. In contrast, a marine anammox species, “Ca. Scalindua sp.” exhibited much higher values of IC50 = 18.0 µM and DOmax = 51.6 µM without significant lag times. The MSAA of “Ca. Scalindua sp.” was inhibited by only ca. 30% at 10 µM DO (the DO upper limit of suboxic condition [44]).
A wide range of oxygen tolerance capabilities have been reported for freshwater anammox species so far [6]. For example, DOmax was reported to be ~200 µM for “Ca. K. stuttgartiensis”, < 63 µM for “Ca. B. sinica”, < 1 µM for “Ca. B. anammoxidans”, 120 µM for “Ca. B. caroliniensis”, and 70 µM for “Ca. B. fulgida”, respectively. On the other hand, complete inhibition was observed at much lower levels of DO (1.25–3.75 µM) in lab-scale bioreactors [6, 12,13,14]. The reason for the variation in reported values could be partly attributed to formation of microbial aggregates and the presence of coexisting aerobes (i.e., the purity of anammox biomass). Therefore, intrinsic oxygen tolerance capabilities of these anammox bacteria cannot be simply evaluated and compared with these reported data.
In the present study, it was confirmed that since highly enriched (>98%) and well-dispersed cells were subjected to the oxygen inhibition studies, the influence of oxygen consumption by coexisting aerobes and/or oxygen shielding effects of aerobes in microbial aggregates can be excluded. Therefore, the observed oxygen tolerances could conceivably be their intrinsic properties. Anammox activities of freshwater species (“Ca. B. sinica”, “Ca. B. sapporoensis”, “Ca. J. caeni”, and “Ca. K. stuttgartiensis”) were completely inhibited at ~25 µM DO with IC50 of 2.7–4.2 µM DO, which are in the middle of the reported range.
The previous O2 amendment studies have shown that anammox activities of oceanic water samples (i.e., Namibian OMZ, Black sea, and Peruvian OMZ) were 50% inhibited at ~ 0.9 µM [4], ~ 8 µM [10], and 1.9 to 16 µM DO [11], respectively. The upper DO limits for anammox in marine systems were reported to be ~20 µM [9, 11, 45] or slightly lower ~10 µM [4, 8, 10]. The IC50 and DOmax values of “Ca. Scalindua sp.” obtained in the present study were higher than these previously reported values. One of the reasons could be due to that much higher cell density (~109 - 1011 copies mL−1) were utilized in the present study than those O2 amendment studies of oceanic OMZ samples (usually ~104–106 copies mL−1) [9, 45]. Since anammox activity is highly dependent on cell density [46, 47], lower cell density makes them more susceptible to oxygen inhibition (Fig. S3). Similar inoculum size-dependent oxygen tolerance ability was previously reported for an obligatory anaerobe, Clostridium butyricum [48].
Another reason could be that since planktonic free-living “Ca. Scalindua sp.” has been continuously cultured for more than 10 years in MBRs in our laboratory, in which it is difficult to maintain strict anoxic conditions, oxygen insensitive Scalindua cells could have been selectively enriched during such a long cultivation period. Since anammox bacterial cells in our MBRs are highly enriched and planktonic biomass, not aggregated one, as shown in FISH images (Fig. 1), oxygen shielding effect and/or oxygen consumption by coexisting planktonic aerobes seems to be minimum.
The experimental results clearly suggest that a marine anammox species “Ca. Scalindua sp.” intrinsically possess higher oxygen tolerance than freshwater species. More importantly, the upper DO limit for “Ca. Scalindua sp.” was much higher than the values reported so far (~20 µM). This might suggest that the ocean volume, where N2 production (N loss) by anammox is potentially expected, could be larger than we have been led to believe so far.
Oxygen reduction rates
“Ca. B. sinica”, “Ca. K. stuttgartiensis”, and “Ca. Scalindua sp.” were cultured in the presence of oxygen (ca. 0.18–0.20% headspace O2). For all three anammox species, O2 gradually decreased with the incubation time while producing 14+15N2 in batch cultures (Fig. 4). The O2 reduction rates were determined by dividing the slope of linear regression of DO concentration by the protein concentration. The oxygen reduction rates of “Ca. B. sinica” and “Ca. Scalindua sp.” were similar (0.26 nmole O2/g-protein/h), whereas “Ca. K. stuttgartiensis” reduced O2 at a more rapid rate (0.53 nmole O2/g-protein/h). The specific 14+15N2 production rates of “Ca. B. sinica”, “Ca. K. stuttgartiensis”, and “Ca. Scalindua sp.” were 3.31 µmole N2/g-protein/h, 3.91 µmole N2/g-protein/h, and 3.44 µmole N2/g-protein/h, respectively. The O2 reduction rates were 4 orders of magnitude lower than the specific 14+15N2 production rate, suggesting that O2 could be reduced for detoxification not for respiration. As discussed below (Section of comparative genome analysis), O2 can be enzymatically reduced to H2O by nonrespiratory proteins for detoxification.

The selected “Ca. B. sinica” (A), “Ca. K. stuttgartiensis” (B), and “Ca. Scalindua sp.” (C) were incubated with 15NO2− (3 mM) and 14NH4+ (3 mM) in the presence of oxygen at room temperature (ca. 25 °C). The headspace 14+15N2 and O2 concentrations were measured with GC/MS. DO concentrations were calculated based on the standard curves of the measured DO concentration (µM) vs. the headspace O2 concentration (%) at 25 °C and 1.5 atm (Fig. S1, see Materials and Methods). The protein concentration was 0.28, 0.15, and 0.47 mg-protein/mL for “Ca. B. sinica”, “Ca. K. stuttgartiensis”, and “Ca. Scalindua sp.”, respectively. The batch experiments were conducted in duplicates. One of representative data sets is presented here.
Recovery of anammox activity after O2 exposure
After 12-h exposure to varying headspace O2 concentrations (0, 0.7, 1.4, 2.1, and 21% (ambient air) O2 at 1.5 atm) in the absence of NH4+ and NO2−, the headspace gas was exchanged with highly pure Helium gas (> 99.9999%) by vacuuming and purging 3 times to restore anoxic conditions. (14NH4)2SO4 (3 mM) and Na15NO2 (3 mM) were supplemented at 0 h, and the recovery of SAA (14+15N2 production rate) was examined (Fig. S4). The 14+15N2 production was immediately detected without any lag time for “Ca. Scalindua sp.” even after exposure to ambient air (21% O2) for 12 h. Subsequent anoxic incubation resulted in almost full recovery: 94 ± 3%, 95 ± 13%, 97 ± 3%, and 65 ± 36% in MSAA after exposure to 0.7%, 1.4%, 2.1 %, and 21% (ambient air) O2 for 12 h, respectively (Fig. 5A). In contrast, “Ca. B. sinica” and “Ca. K. stuttgartiensis” were severely inhibited by all O2 concentrations and could not recover within 7 h anoxic incubation. However, “Ca. B. sinica” exposed to 21% O2 for 12 h regained 80 ± 15% of the initial activity 13 h after the restoration of anoxic incubation, whereas “Ca. K. stuttgartiensis” could not recover at all, indicating irreversible inhibition (Fig. S5). “Ca. J. caeni” gradually recovered 42 ± 2% of initial activity after exposed to 21% O2 with lag times (ca. 0.5–1.5 h). It should be noted that the Jettenia biomass concentrations were about 10 times higher than other species, which could result in the quicker and better recovery. “Ca. B. sapporoensis” exhibited an immediate recovery of 14+15N2 production, however, this could be due to the formation of small flocs during the recovery test, in which O2 transfer limitation could shield anammox bacteria from O2 exposure and thus anammox bacteria could remain active inside.

After 12 h exposure to different headspace O2 concentrations, relative maximum specific anammox activities (MSAA) were determined immediately (A) and after 24-h anoxic incubation (B) in the absence of NH4+ and NO2−. The initial MSAA measured under anoxic condition before O2 exposure was defined as 100% (Anoxic). The data represent the average ± standard deviations of triplicate samples. It should be noted that “Ca. B. sapporoensis” formed small flocs in this recovery experiments, and that the biomass concentrations of “Ca. J. caeni” were about 10 times higher than other species. These could be reasons for higher recovery from O2 exposure.
After 24-h anoxic incubation, the SAAs further recovered > 40% of anoxic control activity, except for “Ca. K. stuttgartiensis” (Fig. 5B). This high oxygen tolerance is partly because the oxygen sensitivity and reversibility of the oxygen inhibition were assessed in the absence of NH4+ and NO2− in the present study, indicating that cells were metabolically inactive. The metabolically inactive cells are likely less susceptible to oxygen inhibition than active cells because of the minimum production of NAD(P)H, which is probably acting as electron donor for O2 reduction (i.e., production of reactive oxygen species, ROS) [49].
The effect of longer O2 exposure time (24 h) on the MSAA recovery was also examined in the absence of NH4+ and NO2− under ambient air (Fig. S6). The 14+15N2 profiles after 24 h-exposure to ambient air exhibited a recovery trend similar to 12 h-exposure (Fig. S5). “Ca. Scalindua sp.” immediately reinitiated almost the same activities as anoxic controls even after 24 h exposure to ambient air. Interestingly, “Ca. B. sinica” gradually increased to 80 ± 15% of the initial activity after 18 h lag time, whereas “Ca. K. stuttgartiensis” could not recover during > 30 h of anoxic incubation. These results coincided with those of the 12 h-exposure test (Fig. S5).
These experimental results suggest that anammox bacteria were aerotolerant anaerobes, although anammox bacteria are previously classified as strict anaerobes. A marine species, “Ca. Scalindua sp.”, exhibited the highest aerotolerance and reversibility among anammox species studied. Furthermore, the oxygen inhibition was reversible except for “Ca. K. stuttgartiensis”. The reversibility seems to depend on the exposure conditions; reversible at low DO levels (0.25–2% O2) but probably irreversibly inhibited at high DO levels (> 46.9 µM or 20% O2) for freshwater anammox species [12,13,14]. The O2 toxicity arises directly from itself or from reactive oxygen species (ROS). O2 and/or ROS reacts unspecifically with catalytic centres or accessory metals of redox enzymes and proteins. For example, key enzymes involved in energy metabolism in anaerobes such as pyruvate ferrodoxin oxidoreductase (PFOR), pyruvate formate lyase (PFL), CO-dehydrogenease, 4-hydroxy-butyryl-CoA dehaydratase, and proteins containing low potential iron-sulfur (Fe-S) clusters are known to be O2 sensitive [50, 51]. However, unfortunately transcriptomic and/or proteomic studies have not been conducted to confirm that these enzymes or proteins were indeed inactivated by molecular O2 and repaired after the restoration of anoxic incubation in the present study.
In the present study, since the anammox biomass was highly enriched and well dispersed, the obtained responses to O2 exposure and recovery from O2 inhibition are most likely intrinsic oxygen tolerance capability of anammox species. For granular biomass, co-existing micro-aerobic heterotrophic bacteria and O2 transfer limitation could shield anammox bacteria from O2 exposure, resulting in higher resilience as compared to planktonic free-living anammox bacteria. For example, floc-style “Brocadia” biomass (dominated only 75%) reinitiated activity to 55-80% of pre-exposed activity after exposure to air-saturated DO (ca. 250 µM) for 24 h [14].
Comparative genome analysis
ROS detoxification mechanisms
Upon exposure to O2, anaerobes need to detoxify O2 and generated ROS (e.g., superoxide anion (O2•-) and hydrogen peroxide (H2O2)) for survival [52]. O2 can be enzymatically reduced to H2O by nonrespiratory flavodiiron proteins (Fdp), named rubredoxin:oxygen oxidoreductase (Roo) [50, 53], reverse rubrerythrins (revRbr) [50, 54], and/or terminal oxidase [55] (Fig. S7A). Fdps (Roo) and revRbr are widely distributed among anaerobic or microaerophilic bacteria and archaea but not in aerobes [50]. Since O2•- is more toxic than H2O2, the detoxification of O2•- is a pivotal function in O2 tolerance. O2•- can be converted to H2O2 and O2 by superoxide dismutase (Sod), which plays a key role in ROS detoxification. The generated H2O2 can be further converted to H2O and O2 by catalase (Cat). In general, aerobes and facultative anaerobes utilize the canonical Sod-Cat detoxification system with O2 generation.
Alternatively, O2•- can be reduced to H2O2 by superoxide reductase (Sor) such as neelaredoxin (Nlr) and desulfoferrodoxin (Dfx) with reduced rubredoxin (Rdred) as an electron donor [56] (Fig. S7B). The oxidized rubredoxin (Rdox) is reduced back to the reduced form by NADH-dependent rubredoxin oxidoreductase (Nror). The generated H2O2 is reduced to H2O by peroxidase such as cytochrome c peroxidases (Ccps) and rubrerythrins (Rbrs).
The electron donor of Ccps and Rbrs is cytochrome c and Rdred, respectively. The Sor-Ccp and/or Rbr detoxification system confers a selective advantage on anammox bacteria because O2 is not generated.
All anammox species possess the genes encoding the class A Fdp [57] with the N-terminal methallo-β-lactamase domain and the C-terminal flavodoxin domain and rubrerythrin, which were considered to function in O2 and/or NO detoxification [58, 59] (Table 1, Table S1). Intriguingly, only the Scalindua genomes (except for the Scalindua sp001828595) contained an operon encoding Fdp and rubredoxin (Rd) which is a potential electron carrier of Fdp. In the operon, the rd gene is located downstream of the fdp gene, suggesting the expression of Scalindua fdp and rd is regulated under the same regulation factor. As for the terminal oxidase, the Scalindua and Kuenenia genomes have the genes encoding cbb3-type cytochrome c which can be involved in O2 reduction [60]. Cytochrome ba3 oxidase and cytochrome oxidase were not identified in all anammox species. In addition, the gene encoding a flavorubredoxin (flavoRd) with a NO-binding non-heme diiron center was identified in all anammox genomes, which possibly function in NO and/or O2 reduction and detoxification [61, 62]. The gene encoding Nror with N-terminal FAD/NAD-binding domain and C-terminal Rd-binding domain [63, 64] was identified in the Scalindua, Jettenia, and Brocadia genomes.
The gene encoding putative Sod (the kustd1303 protein) was widely conserved in the genomes of B. sinica, B. sapporoensis, J. caeni, and K. stuttgartiensis, and also in the other Brocadiaceae genomes except for those affiliated into the Brocadiaceae-family (Table S1). However, only the Scalindua sp. (affiliated into the species SCALAELEC01 sp004282745) and other Scalindua genomes (Scalindua sp001828595 and Scalindua japonica) have a gene encoding typical Fe/Mn-type Sod (sodA or sodB) (Table 1). Multiple sequence alignment of the anammox bacterial Sod showed that all the metal binding sites were conserved among the Scalindua Sod (Fig. S8). On the other hand, all the Brocadiaceae Sod lacked the histidine residues requiring for the metal binding (i.e., His24 and His75), and N-terminal alpha-hairpin domain was also not conserved in the Brocadiaceae Sod. Furthermore, phylogenetic analysis of anammox bacterial Sod revealed that Scalindua Sod and Brocadiaceae Sod are affiliated into different phylogenetic clades (Fig. S9). These evidences indicate the Brocadiaceae Sod is likely not to function.
As for O2•- reduction to H2O2 by Sor, the gene encoding neelaredoxin (Nlr) with functionally-important amino acid residues [65] (Fig. S10) was widely conserved among the anammox bacterial genomes except for Jettenia (Table S1). As for reduction of the generated H2O2, all genera possess the genes encoding cytochrome c peroxidase (Ccp) that obtains reducing equivalents from cytochrome c, whereas B. sinica, K. stuttgartiensis, and Scalindua sp. possess the genes encoding catalase (Cat). The B. sinica and Scalindua sp. Cat were affiliated into a phylogenetic clade apart from that of K. stuttgartiensis Cat (Fig. S11). The genes encoding another peroxidase, rubrerythrins (Rbr) that obtains reducing equivalents from reduced rubredoxin (Rdred) and glutathione peroxidase (Gpx) that obtains reducing equivalents from glutathione, were conserved in only Scalindua sp.
Based on the comparative genome analysis, it was clearly confirmed that all anammox species commonly possess the genes considered to function for O2 reduction (i.e., fdpA and fdpF), O2•- reduction (i.e., nlr), and H2O2 reduction (i.e., rbr and ccp) for survival under microaerobic conditions. The Sor-peroxidase dependent O2•- and H2O2 reduction system is much advantageous to anammox bacteria because O2 is not generated in these reduction reactions. However, the Sor-peroxidase dependent detoxification system alone may not be sufficient for cell survival under high O2 conditions. Intriguingly, only “Ca. Scalindua sp.” possess the genes for a classical Sod-Cat dependent O2•- and H2O2 detoxification system (i.e., sodA or sodB) and a functional Fdp-rubredoxin operon, which could be responsible for the higher O2 tolerance than other freshwater anammox species. The presence of Sod-Cat system on the Scalindua genome triggered the experimental investigation of those enzymatic activities.
Activities of anti-oxidative enzymes in different anammox bacteria
To explain the inter-species difference in O2 tolerance, the activities of major anti-oxidative enzymes (Sod, Cat, Ccp, and Gpx) of “Ca. Scalindua sp.”, “Ca. J. caeni”, “Ca. B. sinica”, “Ca. B. sapporoensis” and “Ca. K. stuttgartiensis” were assayed for their cell-free extracts prepared from biomass collected from respective anaerobic MBR cultures. Only Scalindua exhibited high Sod (converts O2•- to H2O2 and O2) activity of 22.6 ± 1.9 U/mg-protein with relatively low Cat (converts H2O2 to O2) activity of 1.6 ± 0.7 U/mg-protein (Fig. 6A). This Sod activity level is similar to those of aerobic and facultative bacteria (10.9–49.7 U/mg-protein) [66], but slightly higher than aerotolerant anaerobic bacteria (0.44–19.6 U/mg-protein) and higher than intermediate and extremely oxygen sensitive (obligately) anaerobic bacteria (almost not detected) [67]. In contrast, other four freshwater anammox species possessed very low or virtually no Sod activity with moderately high levels of Cat activity (5.9–16.5 U/mg-protein), which might reflect their lower oxygen tolerance capability. The gene encoding catalase was not identified in the J. caeni and B. sapporoensis genomes, which may have been a result of the incomplete nature of the genomes. Since the activities of Sod and Cat were highly dependent on medium compositions and culture conditions [68], the effect of salinity on these activities needs to be investigated in the future. Activities of cytochrome c peroxidase (Ccp converts H2O2 to H2O) were detected in all species except for “Ca. B. sapporoensis” but 3–4 orders of magnitude lower than the Cat activities (Fig. 6B). Activities of glutathione peroxidase (Gpx converts H2O2 to H2O) were barely detected only in “Ca. Scalindua sp.” and “Ca. J. caeni”.

(A: Sod and Cat, B: Ccp and Gpx) in cell-free extracts of anammox bacteria harvested from respective anaerobic MBR cultures. Results are presented as the standard error of the mean of at least 6 samples in two independent experiments.
Effect of exposed O2 concentrations and exposure time on the activities of anti-oxidative enzymes
Cell-free extracts were prepared from anaerobic MBR cultures of “Ca. Scalindua sp.”, “Ca. B. sinica”, “Ca. J. caeni”, and “Ca. K. stuttgartiensis” were subjected to two patterns of O2 exposure: (1) exposed to different O2 concentrations (0, 0.7, 1.4 and 2.1% O2) for 12 h and (2) exposed to ambient air (21% O2) for different periods of time (0, 0.5, 1 and 2 h). High activity levels of Sod were detected in all samples of “Ca. Scalindua sp.”, but no significant effects of exposed O2 concentrations and duration of O2 exposure on the Sod activity levels were observed even after exposure to ambient air for 2 h (Fig. S12 and Fig. S13). The Cat activity in “Ca. Scalindua sp.” was also remained unchanged for different O2 concentrations and durations of O2 exposure. Similarly, O2 concentrations and O2 exposure times did not significantly affect the activity levels of all anti-oxidative enzymes for other three freshwater species. These results revealed that the activities of all anti-oxidative enzymes could be constitutively expressed and active in all anammox bacteria.
The oxygen tolerance is directly related to the ability of the bacteria to reduce (scavenge) O2 and to detoxify O2•- (namely Sod activity). Thus, Sod activity is a primary important determinant of oxygen tolerance because O2•- is more toxic than H2O2 [66], whereas Cat is the secondary importance since catalase activity showed no clear correlation to oxygen tolerance [66]. “Ca. Scalindua sp.” possessed significantly higher Sod activity and therefore exhibited higher oxygen tolerance than other freshwater species. It is likely that O2•- was primarily detoxified by Sod in “Ca. Scalindua sp.”, whereas other four freshwater species cannot efficiently detoxify O2•- due to lack of Sod activity. The generated H2O2 could be converted to O2 and H2O by Cat and/or to H2O by peroxidases (Ccp, Gpx, and/or rubrerythrin (Rbr)) (Fig. S7). However, since Ccp and Rbr were active only in the absence of O2 with low levels of H2O2 and also unable to degrade H2O2 quickly [69, 70], it is not clear if Ccp and Rbr play a vital role in H2O2 decomposition or not in “Ca. Scalindua sp.”. Catalase is the most prominent of the stationary-phase induced scavengers and able to degrade H2O2 more quickly at higher concentrations, whereas rubrerythrins are used to scavenge low levels of H2O2 [70]. This suggests that “Ca. Scalindua sp.” most likely employs different peroxidases including Cat depending on H2O2 concentration [51]. Further study is definitely required to identify which enzyme(s) acts effectively in the H2O2 detoxification in “Ca. Scalindua sp.”.
In the presence of oxygen, anammox bacteria reduce O2 to form toxic ROS, or reduce to H2O by Sor. If bacteria reduce no oxygen and therefore no toxic ROS is generated, resulting in higher oxygen tolerance [66]. “Ca. Scalindua sp.” with high Sod activity reduced O2 at a relatively slow rate (0.26 µmole O2/g-protein/h), whereas “Ca. K. stuttgartiensis” that has substantially no Sod activity reduced O2 at a more rapid rate (0.53 µmole O2/g-protein/h). Although “Ca. B. sinica” has no Sod activity, they reduced O2 at a relatively slow rate (0.26 µmole O2/g-protein/h). These results suggest that “Ca. K. stuttgartiensis” generate SOR rapidly but could not detoxify it quickly, resulting in lower O2 tolerance and irreversible recovery. It should be also noted that the oxygen reduction rate is an important factor in determining oxygen tolerance.
In conclusion, a marine species, “Ca. Scalindua sp.”, exhibited the higher aerotolerance and reversibility than other freshwater anammox species. This is primarily because “Ca. Scalindua sp.” possesses the classical Sod-Cat ROS detoxification system in addition to the Sor-peroxidase dependent O2 and ROS reduction system. The upper DO limit for “Ca. Scalindua sp.” (~51.6 µM) was much higher than the values reported so far (~20 µM). This might suggest that the contribution of anammox to oceanic nitrogen loss could be larger than we have ever thought.
Data availability
Anammox bacterial genomes and metagenome-assembled genomes examined in the present study were available in the GenBank and DDBJ database under the following accession number; GCA_900696655.1, GCA_000296795.1, GCA_000786775.1, GCA_000949635.1, GCA_000987375.1, GCA_001723765.1, GCA_001753675.2, GCA_001824485.1, GCA_001828415.1, GCA_001828515.1, GCA_001828545.1, GCA_001828565.1, GCA_001828595.1, GCA_001828605.1, GCA_001828645.1, GCA_001830285.1, GCA_002009475.1, GCA_002050315.1, GCA_002050325.1, GCA_002443295.1, GCA_004282745.1, GCA_005524015.1, GCA_007859995.1, GCA_016187625.1, GCA_016206465.1, GCA_900232105.1, GCF_000949635.1, GCF_001753675.2, GCF_000296795.1, GCF_900232105.1, GCA_004282745.1, BQMK01000001-BQMK01000087, BQMM01000001-BQMM01000139, BQMT01000001-BQMT01000095, BQMU01000001-BQMU01000391, BQMX01000001-BQMX01000120.
References
Lam P, Kuypers MMM. Microbial nitrogen cycling processes in oxygen minimum zones. Ann Rev Mar Sci. 2011;3:317–45.
Dalsgaard T, Thamdrup B, Farias L, Revsbech PN. Anammox and denitrification in the oxygen minimum zone of the eastern South Pacific. Limnol Oceanogr. 2012;57:1331–46.
Kalvelage T, Lavik G, Lam P, Contreras S, Arteaga L, Loscher CR, et al. Nitrogen cycling driven by organic matter export in the South Pacific oxygen minimum zone. Nat Geosci. 2013;6:228–34.
Dalsgaard T, Stewart FJ, Thamdrup B, De Brabandere L, Revsbech NP, Ulloa O, et al. Oxygen at nanomolar levels reversibly suppresses process rates and gene expression in anammox and denitrification in the oxygen minimum zone off northern Chile. mBio. 2014;5:e01966–14.
Paulmier A, Ruiz-Pino D. Oxygen minimum zones (OMZs) in the modern ocean. Prog Oceanogr. 2009;80:113–28.
Oshiki M, Satoh H, Okabe S. Ecology and physiology of anaerobic ammonium oxidizing bacteria. Environ Microbiol. 2016;18:2784–96.
Zhang L, Okabe S. Ecological niche differentiation among anammox bacteria. Water Res. 2020;171:115468.
Babbin AR, Keil RG, Devol AH, Ward BB. Organic matter stoichiometry, flux, and oxygen control nitrogen loss in the ocean. Science. 2014; 344: https://doi.org/10.1126/science.1248364.
Kuypers MMM, Lavik G, Woebken D, Schmid M, Fuchs BM, Amann R, et al. Massive nitrogen loss from the Benguela upwelling system through anaerobic ammonium oxidation. Proc Natl Acad Sci USA. 2005;102:6478–83.
Jensen MM, Kuypers MM, Gaute L, Thamdrup B. Rates and regulation of anaerobic ammonium oxidation and denitrification in the Black Sea. Limnol Oceanogr. 2008;53:23–36.
Kalvelage T, Jensen MM, Contreras S, Revsbech NP, Lam P, et al. Oxygen sensitivity of anammox and coupled N-cycle processes in oxygen minimum zones. PLoS ONE. 2011;6:e29299. https://doi.org/10.1371/journal.pone.0029299.
Strous M, Van Gerven E, Kuenen JG, Jetten M. Effects of aerobic and microaerobic conditions on anaerobic ammonium-oxidizing (anammox) sludge. Appl Environ Microbiol. 1997;63:2446–8.
Egli K, Fanger U, Alvarez PJ, Siegrist H, Meer JR, Zehnder AJ. Enrichment and characterization of an anammox bacterium from a rotating biological contactor treating ammonium-rich leachate. Arch Microbiol. 2001;175:198–207.
Seuntjens D, Carvajal-Arroyo JM, Ruopp M, Bunse P, De Mulder CP, Lochmatter S, et al. High-resolution mapping and modeling of anammox recovery from recurrent oxygen exposure. Water Res. 2018;144:522–31.
Yue X, Yu G, Lu Y, Liu Z, Li Q, Tang J, et al. Effect of dissolved oxygen on nitrogen removal and the microbial community of the completely autotrophic nitrogen removal over nitrite process in a submerged aerated biological filter. Bioresour Technol. 2018;254:67–74.
Kuypers, MM, Lavik, G, Thamdrup, B Anerobic ammonium oxidation in the marine environment. In: Neretin L (eds) Past and Present Water Column Anoxia. Nato Science Series: IV: Earth and Environmental Sciences, 64. Springer, Dordrecht 2006; https://doi.org/10.1007/1-4020-4297-3_13.
Oshiki M, Shimokawa M, Fujii N, Satoh H, Okabe S. Physiological characteristics of the anaerobic ammonium-oxidizing bacterium “Candidatus Brocadia sinica”. Microbiol. 2011;157:1706–13.
Carvajal-Arroyo JM, Sun W, Sierra-Alvarez R, Field JA. Inhibition of anaerobic ammonium oxidizing (anammox) enrichment cultures by substrates, metabolites and common wastewater constituents. Chemosphere. 2013;91:22–27.
Yan J, Haaijer SC, Op den Camp HJ, van Niftrik L, Stahl DA, Könneke M, et al. Mimicking the oxygen minimum zones: stimulating interaction of aerobic archaeal and anaerobic bacterial ammonia oxidizers in a laboratory‐scale model system. Environ Microbiol. 2012;14:3146–58.
Morris JG, The physiology of obligate anaerobiosis. In Advances in microbial physiology. Academic Press. 1975;12:169–246.
Jenney FE Jr., Verhagen MFJM, Cui X, Adams MWW. Anaerobic microbes: oxygen detoxification without superoxide dismutase. Science. 1999;286:306–9.
Kartal B, van Niftrik L, Keltjens JT, Op den Camp HJM, Jetten MSM. Anammox—Growth physiology, cell biology, and metabolism. Adv Microb Physiol. 2012;60:211–62.
Ji X, Wu Z, Sung S, Lee PH. Metagenomics and metatranscriptomics analyses reveal oxygen detoxification and mixotrophic potentials of an enriched anammox culture in a continuous stirred-tank reactor. Wat Res. 2019;166:115039.
Kartal B, Geerts W, Jetten MSM. Cultivation, detection and ecophysiology of anaerobic ammonium-oxidizing bacteria. Methods Enzymol. 2011;486:89–108.
Ali M, Oshiki M, Okabe S. Simple, rapid and effective preservation and reactivation of anaerobic ammonium oxidizing bacterium “Candidatus Brocadia sinica.”. Water Res. 2014;57:215–22.
Oshiki M, Ishii S, Yoshida K, Fujii N, Ishiguro M, Satoh H, et al. Nitrate-dependent ferrous iron oxidation by anaerobic ammonium oxidation (Anammox) bacteria. Appl Environ Microbol. 2013;79:4087–93.
Narita Y, Zhang L, Kimura ZI, Ali M, Fujii T, Okabe S. Enrichment and physiological characterization of an anaerobic ammonium-oxidizing bacterium “Candidatus Brocadia sapporoensis”. Syst Appl Microbiol. 2017;40:448–57.
Zhang L, Narita Y, Gao L, Ali M, Oshiki M, Okabe S. Maximum specific growth rate of anammox bacteria revisited. Water Res. 2017;116:296–303.
Zhang L, Narita Y, Gao L, Ali M, Oshiki M, Ishii S, et al. Microbial competition among anammox bacteria in nitrite-limited bioreactors. Water Res. 2017;125:249–58.
Okabe S, Shafdar AA, Kobayashi K, Zhang L, Oshiki M. Glycogen metabolism of the anammox bacterium “Candidatus Brocadia sinica”. ISMEJ.2020;15: https://doi.org/10.1038/s41396-020-00850-5.
Okabe S, Kamigaito A, Kobayashi K. Maintenance power requirements of anammox bacteria “Candidatus Brocadia sinica” and “Candidatus Scalindua sp. ISME J. 2020;15: https://doi.org/10.1038/s41396-021-01031-8.
van de Graaf AA, de Bruijn P, Robertson LA, Jetten MSM, Kuenen JG. Autotrophic growth of anaerobic ammonium-oxidizing microorganisms in a fluidized bed reactor. Microbiol. 1996;142:2187–96.
Oh J, Silverstein J. Oxygen inhibition of activated sludge denitrification. Water Res. 1999;33:1925–37.
Chaumeil P-A, Mussig AJ, Hugenholtz P, Parks DH. GTDB-Tk: A toolkit to classify genomes with the genome taxonomy database. Bioinformatics. 2019;36:btz848.
Oshiki M, Takaki Y, Hirai M, Nunoura T, Kamigaito A, Okabe S. Metagenomic analysis of five phylogenetically distant anammox bacterial enrichment cultures. Microb Environ. 2022;37:ME22017.
Thompson JD, Higgins DG, Gibson TJ. CLUSTAL W: improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Res. 1994;22:4673–80.
Robert X, Gouet P. Deciphering key features in protein structures with the new ENDscript server. Nuc Acids Res. 2014;42:W320–W324.
Tamura K, Stecher G, Kumar S. MEGA11: Molecular evolutionary genetics analysis version 11. Mol Biol Evol. 2021;38:3022–7.
Beauchamp C, Fridovich I. Superoxide dismutase: Improved assays and an assay applicable to acrylamide gels. Anal Biochem. 1971;44:276–87.
Datkhilea KD, Mukhopadhyaya R, Dongrec TK, Natha BB. Increased level of superoxide dismutase (SOD) activity in larvae of Chironomus ramosus (Diptera: Chironomidae) subjected to ionizing radiation. Comp Biochem Physiol C. 2009;149:500–6.
Aebi, H Catalase. In: Bergmeyer HU (Eds) Methods in Enzymatic Analysis. Academic Press, New York, 1983; 276–86.
Munkres KD, Rana RS, Goldstein E. Genetically determined conidial longevity is positively correlated with superoxide dismutase, catalase, glutathione peroxidase, cytochrome c peroxidase, and ascorbate free radical reductase activities in Neurospora crassa. Mech Ageing Dev. 1984;24:83–100.
Drotar A, Phelps P, Fall R. Evidence for glutathione peroxidase activity in cultured plant cells. Plant Sci. 1985;42:35–40.
Bianchi D, Dunne JP, Sarmiento JL, Galbraith FD. Data-based estimates of suboxia, denitrification, and N2O production in the ocean and their sensitivities to dissolved O2. Global Biogeochem Cycles. 2012;26:GB2009 https://doi.org/10.1029/2011GB004209.
Hamersley M, Robert LG, Woebken D, Rattray JE, Lam P, Hopmans EC Anaerobic ammonium oxidation in the Peruvian oxygen minimum zone, Limnol Oceanogr. 2007; 52: https://doi.org/10.4319/lo.2007.52.3.0923.
Oshiki M, Hiraizumi H, Satoh H, Okabe S. Cell density-dependent anammox activity of “Candidatus Brocadia sinica” regulated by N-acyl homoserine lactone-mediated quorum sensing. Microb Environ. 2020;35:ME20086.
Strous M, Fuerst J, Kramer E, Logemann S, Muyzer G, van de Pas-Schoonen K, et al. Missing lithotroph identified as new planctomycete. Nature. 1999;400:446–9.
Kawasaki S, Nakagawa T, Nishiyama Y, Benno Y, Uchimura T, Komagata M, et al. Effect of oxygen on the growth of Clostridium butyricum (type species of the genus Clostridium), and the distribution of enzymes for oxygen and for active oxygen species in Clostridia. J Ferment Bioeng. 1998;86:368–72.
Abreu IA, Saraiva LM, Carita J, Huber H, Stetter KO, Cabelli D, et al. Oxygen detoxification in the strict anaerobic archaeon Archaeoglobus fulgidus: superoxide scavenging by Neelaredoxin. Mol Microbiol. 2000;38:322–34.
Morvan C, Folgosa F, Kint N, Teixeira M, Martin-Verstraete I. Responses of Clostrida to oxygen: From detoxification to adaptive strategies. Environ Microbiol. 2021;23:4112–25.
Lu Z, Imlay JA. When anaerobes encounter oxygen: Mechanisms of oxygen toxicity, tolerance and defence. Nat Rev Microbiol. 2021;19:774–85.
Kint N, Morvan C, Martin-Verstraete I. Oxygen response and tolerance mechanisms in Clostridioides difficile. Curr Opin Microbiol. 2022;65:175–82.
Wildschut JD, Lang RM, Voordouw JK, Voordouw G. Rubredoxin: Oxygen oxidoreductase enhances survival of Desulfovibrio vulgaris Hildenborough under microaerophilic conditions. J Bacteriol. 2006;188:6253–60.
Kint N, Feliciano CA, Martins MC, Morvan C, Fernandes SF, Folgosa F, et al. How the anaerobic enteropathogen Clostridioides difficile tolerates low O2 tensions. mBio. 2020;11:e01559–20.
Baughn AD, Malamy MH. The strict anaerobe Bacteroides fragilis grows in and benefits from nanomolar concentrations of oxygen. Nature. 2004;427:441–4.
Rodrigues J, Saraiva LM, Abreu IA, Teixeira M, Cabelli DE. Superoxide reduction by Archaeoglobus fulgidus desulfoferrodoxin: Comparison with neelaredoxin. J Biol Inorg Chem. 2007;12:248–56.
Folgosa F, Martins MC, Teixeira M. Diversity and complexity of flavodiiron NO/O2 reductases. FEMS Microbiol Lett. 2018;365:fnx267. https://doi.org/10.1093/femsle/fnx267.
Folgosa F, Martins MC, Teixeira M. The multidomain flavodiiron protein from Clostridium difficile 630 is an NADH: Oxygen oxidoreductase. Sci Rep. 2018;8:10164.
Goncalves VL, Vicente JB, Pinto L, Romão CV, Frazão C, Sarti P, et al. Flavodiiron oxygen reductase from Entamoeba histolytica modulation of substrate preference by tyrosine 271 and lysine 53. J Biol Chem. 2014;289:28260–70.
Kartal B, Almeida NM, de Maalcke WJ, Op den Camp HJM, Jetten MSM, Keltjens JT. How to make a living from anaerobic ammonium oxidation. FEMS Microbiol Rev. 2013;37:428–61.
Gardner AM, Helmick RA, Gardner PR. Flavorubredoxin, an inducible catalyst for nitric oxide reduction and detoxification in Escherichia coli. J Biol Chem. 2002;277:8172–7.
Silaghi-Dumitrescu R, Ng KY, Viswanathan R, Kurtz DM Jr. A flavo-protein from Desulfovibrio vulgaris with oxidase and nitric oxide reductase activities. Evidence for an in vivo nitric oxide scavenging function. Biochem. 2005;44:3572–9.
Guedon E, Petitdemange H. Identification of the gene encoding NADH-rubredoxin oxidoreductase in Clostridium acetobutylicum. Biochem Biophys Res Commun. 2001;285:496–502.
Nishikawa K, Shomura Y, Kawasaki S, Niimura Y, Higuchi Y. Crystal structure of NADH: Rubredoxin oxidoreductase from Clostridium acetobutylicum: A key component of the dioxygen scavenging system in obligatory anaerobes. Proteins. 2010;78:1066–70.
Pereira AS, Tavares P, Folgosa F, Almeida RM, Moura I, Moura JJG. Superoxide reductases. Eur J Inorg Chem. 2007;18:2569–81.
Rolfe RD, Hentges DJ, Campbell BJ, Barrett JT. Factors related to the oxygen tolerance of anaerobic bacteria. Appl Environ Microbiol. 1978;36:306–13.
Tally FP, Goldin BR, Jacobus NV, Gorbach SL. Superoxide dismutase in anaerobic bacteria of clinical significance. Infect Immun. 1977;16:20–25.
Gregory EM, Kowalski JB, Holdeman LV. Production and some properties of catalase and superoxide dismutase from the anaerobe Bacteroides distasonis. J Bacteriol. 1977;129:1298–302.
Khademian M, Imlay JA. Escherichia coli cytochrome c peroxidase is a respiratory oxidase that enables the use of hydrogen peroxide as a terminal electron acceptor. Proc Natl Acad Sci USA. 2017;114:E6922–E6931.
Mishra S, Imaly J. An anaerobic bacterium, Bacteroides thetaiotaomicron, uses a consortium of enzymes to scavenge hydrogen peroxide. Mol Microbiol. 2013;90:1356–71.
Acknowledgements
This research was financially supported by JSPS KAKENHI Grant-in-Aid for Scientific Research (A) Grant Number 19H00776, which was granted to Satoshi Okabe, and by JST FOREST Program Grant Number JPMJFR216Z granted to Mamoru Oshiki. Computations were partially performed on the NIG supercomputer at ROIS National Institute of Genetics.
Author information
Authors and Affiliations
Contributions
Conceptualization: SO; Data curation: SY, XL, KN, HZ, KK; Formal analysis: SO, SY, MO; Funding acquisition: SO, MO; Investigation: SO, MO; Visualization: SO, MO; Writing – original draft: SY, SO; Writing – review & editing: SO, MO.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Okabe, S., Ye, S., Lan, X. et al. Oxygen tolerance and detoxification mechanisms of highly enriched planktonic anaerobic ammonium-oxidizing (anammox) bacteria. ISME COMMUN. 3, 45 (2023). https://doi.org/10.1038/s43705-023-00251-7
Received:
Revised:
Accepted:
Published:
DOI: https://doi.org/10.1038/s43705-023-00251-7
