
Abstract
As coronavirus disease 2019 (COVID-19) and aging are both accompanied by cognitive decline, we hypothesized that COVID-19 might lead to molecular signatures similar to aging. We performed whole-transcriptome analysis of the frontal cortex, a critical area for cognitive function, in individuals with COVID-19, age-matched and sex-matched uninfected controls, and uninfected individuals with intensive care unit/ventilator treatment. Our findings indicate that COVID-19 is associated with molecular signatures of brain aging and emphasize the value of neurological follow-up in recovered individuals.
Similar content being viewed by others

Main
COVID-19 is an acute respiratory disease often accompanied by neurological sequelae1. Individuals with previous severe COVID-19 exhibit a 10-year average drop in their global cognitive performance2, mimicking accelerated aging. Complementary studies combining neuroimaging and cognitive screening implicate COVID-19-induced impairment of the frontal cortex3,4, a critical area for cognitive function, but molecular evidence of aging-like effects in the brain is lacking.
To address this, we performed RNA-sequencing (RNA-seq) analysis of 54 postmortem frontal cortex samples, including samples from 21 individuals with severe COVID-19 (previous neurological history was limited to Alzheimer’s disease in one person and epilepsy in another) and 1 asymptomatic individual aged between 23 and 84 years old, 22 age-matched (±2 years) and sex-matched uninfected controls with no history of neurological or psychiatric disorders, an age-matched and sex-matched uninfected individual with Alzheimer’s disease, and an additional independent control group of 9 uninfected individuals with history of intensive care unit (ICU) or ventilator treatment (22–85 years old; ICU/VENT; Fig. 1a and Supplementary Fig. 1a; see Supplementary Table 1 for clinical information; COVID-19 cohort). All COVID-19-cases were determined by positive pre-mortem or peri-mortem testing for severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) infection via nasopharyngeal swab qPCR and history of hospitalization, whereas uninfected control samples were collected before the COVID-19 pandemic (with three exceptions in the ICU/VENT group that had negative SARS-CoV-2 qPCR tests at the time of death, and no COVID-19 history and/or negative serological test).

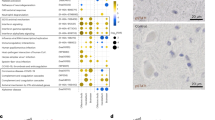
a, Age and sex of each individual in COVID-19 or uninfected age/sex-matched control (±2 years) groups (n = 22 per group) analyzed in this cohort. An asterisk indicates notable COVID-19 cases. The 23-year-old male presented with asymptomatic COVID-19, the 62-year-old female presented with severe COVID-19 history and comorbid epilepsy and the 84-year-old female who had a history of severe COVID-19 with comorbid Alzheimer’s disease (AD; an uninfected individual with AD was also included as an additional control; Supplementary Table 1). Created with BioRender.com. b, t-distributed stochastic neighbor embedding (t-SNE) analysis of frontal cortex transcriptomes from COVID-19-infected individuals, uninfected age-matched and sex-matched controls, and an independent group of uninfected controls with history of ICU and/or ventilator treatment (ICU/VENT). Black border, 23-year-old asymptomatic male with COVID-19. Red border, 62-year-old female with COVID-19 history and comorbid epilepsy. Blue border, 84-year-old female with COVID-19 history and comorbid AD. Black point, 84-year-old female without COVID-19 but with AD. Green border, uninfected age-matched and sex-matched control (non-AD) for the COVID-19-infected individual with comorbid AD. For age-matched/sex-matched controls and COVID-19 samples n = 22 per group; ICU/VENT-treated uninfected controls n = 9. c, Volcano plot representing the DEGs of the frontal cortex of individuals with COVID-19 versus age-matched and sex-matched controls (n = 22 per group). Red points denote significantly upregulated genes among COVID-19 cases (false discovery rate (FDR) < 0.05). Blue points denote significantly downregulated genes among COVID-19 cases. Black points highlight significant genes with corresponding gene symbols (Supplementary Table 2). d, Gene Ontology (GO) biological pathway enrichment analysis of COVID-19 versus age-matched/sex-matched control DEGs. Gene ranks were determined by signed −log10 FDRs of DEGs (Supplementary Table 3). e, GSEA of cognitive decline-regulated genes using COVID-19 (COVID-19 versus age-matched and sex-matched controls) DEGs. DEG ranks were assigned by signed −log10 FDR from the frontal cortex transcriptome of individuals with MMSE scores > 25 (high cognitive performance) versus the transcriptome of individuals with MMSE scores < 25 (low cognitive performance/cognitive decline) as measured in the ROSMAP study. f,g, GSEA of COVID-19 DEGs (COVID-19 versus age-matched/sex-matched control in f and COVID-19 versus ICU/VENT in g), using significantly upregulated (top) or downregulated genes (bottom) in our aging cohort as gene sets. DEG ranks were assigned by signed −log10 FDR from COVID-19 versus corresponding control frontal cortex. NES, normalized enrichment score. P, two-tailed GSEA P value (Supplementary Fig. 5).
By clustering analyses, COVID-19 transcriptomic cases broadly segregated away from controls, with two of the outliers being from the 23-year-old asymptomatic individual and the 62-year-old individual with comorbid epilepsy; the age-matched/sex-matched controls proximal to COVID-19 cases were from older adults (Fig. 1b and Supplementary Figs. 1 and 2). Uninfected older adults in the ICU/VENT group generally clustered closer to COVID-19-infected individuals, whereas younger ICU/VENT individuals clustered relatively close to controls; two cases cluster separately from all other samples (Supplementary Fig. 1b). Comparison of COVID-19 cases and their corresponding age-matched and sex-matched controls revealed 6,993 differentially expressed genes (DEGs), 3,330 of which were upregulated and 3,663 downregulated (Fig. 1c and Supplementary Table 2). For example, the S100A8 and S100A9 genes, which encode calprotectin and blood circulating levels of which distinguish severe from mild COVID-19 disease5, were upregulated among individuals with COVID-19. Pathway enrichment analysis identified numerous significant GO terms associated with aging in the human brain enriched upon severe COVID-19, including positive enrichment of immune-related pathways and negative enrichment of synaptic activity, cognition and memory pathways (Fig. 1d and Supplementary Fig. 3). We also observed significant associations of cellular response to DNA damage, mitochondrial function, regulation of response to stress and oxidative stress, vesicular transport, calcium homeostasis6, and insulin signaling/secretion7 pathways previously associated with aging processes and brain aging6,8. Altogether, our analyses suggest that many biological pathways that change with natural aging in the brain also change in severe COVID-19.
As natural brain aging is associated with cognitive decline, we further assessed associations of transcriptomic changes in COVID-19 and cognitive function. We collated frontal cortex transcriptomic data from 633 individuals who underwent the Mini-Mental State Examination (MMSE) while alive and donated their brains at the time of death as part of the ROSMAP study9,10. We split individuals and their corresponding transcriptomic data based on the median MMSE score: ≥25 as high cognitive performance versus MMSE < 25 as low cognitive performance. From gene-set enrichment analysis (GSEA), we found strong associations between low cognitive performance and COVID-19 (Fig. 1e).
Given the strong associations between aging-regulated pathways and severe COVID-19, we sought to directly test whether COVID-19 is associated with similar gene expression patterns as natural aging in the human brain. We performed RNA-seq analysis of postmortem frontal cortex samples of 10 young (≤38 years old) and 10 older (≥71 years old) uninfected individuals (Supplementary Table 1; aging cohort and Supplementary Fig. 4a) and compared these findings to COVID-19 DEGs. We found striking similarities between individuals with COVID-19 and aged individuals: genes upregulated in aging were upregulated in severe COVID-19; likewise, genes downregulated in aging were also downregulated in severe COVID-19 (see Fig. 1f for age-matched/sex-matched controls versus COVID-19). As further validation, we collated transcriptome-wide datasets from five independent aging cohorts and confirmed this association (Supplementary Fig. 5 and Supplementary Table 4). Intriguingly, we continued to observe a significant association between aging-associated genes and DEGs from individuals with COVID-19 versus uninfected individuals treated with ICU/VENT (Fig. 1g).
To delineate the effects of severe COVID-19 on brain aging directly, we leveraged our aging cohort to derive an aging index (Supplementary Fig. 4), comprising our aging DEGs and condensed by the first principal component across these transcriptomic data. As validation, we applied our predicted aging index to uninfected controls (COVID-19 cohort) and found similarly strong Pearson correlations between the training and test sets (Fig. 2a,b). Applying this model to individuals with COVID-19, we observed a significant increase in the predicted aging index compared to corresponding uninfected age-matched/sex-matched control and uninfected ICU/VENT control groups (Fig. 2c). Additional analysis revealed that the predicted aging index in individuals with COVID-19 was not significantly affected by the presence or absence of cerebrovascular disease (P > 0.05). Thus, severe COVID-19 appears to shift the molecular age of brains relative to both uninfected age-matched and sex-matched controls as well as uninfected ICU/VENT controls. Lastly, using qPCR analysis, we validated several of the top shared DEGs between our COVID-19 cohort and aging datasets (n = 22 per group; Supplementary Fig. 6).

a, First, a principal-component analysis using DEGs (FDR < 0.05) of young versus old uninfected controls estimated principal component 1 (PC1). The graph presents a two-tailed Pearson correlation of chronological age with aging index (PC1) among young versus old uninfected controls from the aging cohort (training set). n = 20. Gray shadow indicates the 95% confidence interval from a linear regression fit (R2 (18) = 0.58, P = 9.2 × 10−5). b, Two-tailed Pearson correlation of chronological age with predicted aging index (PC1) among uninfected age-matched and sex-matched controls from the COVID-19 cohort (test set). n = 22. Gray shadow indicates the 95% confidence interval from a linear regression fit (R2 (20) = 0.438, P = 7.9 × 10−4). c, Predicted aging index (PC1) of individuals with COVID-19 (n = 22), age-matched/sex-matched uninfected controls (control; n = 22), and an independent group of uninfected cases with ICU/VENT treatment history (n = 9). The line in each group represents the mean ± s.e.m. COVID-19 versus control Welch two-tailed t(42.0) = 5.68, P = 3.4 × 10−6; COVID-19 versus ICU/VENT Welch two-tailed t(21.0) = 3.14, P = 0.015. A Bonferroni correction was used to adjust for multiple comparisons. d, Significant interferon and TNF-related pathways identified using GO biological pathway enrichment analysis of COVID-19 versus age-matched/sex-matched control frontal cortex DEGs. FDR, GSEA FDR (Supplementary Table 3). e, Experimental design of in vitro cytokine treatment in human neurons. Created with BioRender.com. f, Effects of IFN-β, IFN-γ and TNF on predicted aging index, as assessed following in vitro treatment of primary human neurons. The line in each group represents the mean ± s.e.m. n = 3 independent wells (1 × 105 cells per well were plated) for each treatment. IFN-β_LO versus control Welch two-tailed t(2.68) = 4.48, P = 0.16; IFN-β_HI versus control Welch two-tailed t(3.51) = 8.26, P = 0.012; IFN-γ_LO versus control Welch two-tailed t(3.58) = 20.8, P = 4.4 × 10−4; IFN-γ_HI versus control Welch two-tailed t(3.35) = 12.3, P = 4.1 × 10−3; TNF_LO versus control Welch two-tailed t(3.63) = 33.6, P = 6.6 × 10−5; TNF_HI versus control Welch two-tailed t(2.28) = 15.8, P = 0.013. Bonferroni correction was used to adjust for multiple comparisons.
Finally, we sought to determine pathophysiologic mechanisms that may explain the association of COVID-19 with aging. We considered that this could be due to multiple factors, including SARS-CoV-2 viral infection in the frontal cortex or COVID-19-induced systemic inflammation. In agreement with previous studies11,12, SARS-CoV-2 viral RNA was not detected in samples from individuals with COVID-19 (at the time of death; Supplementary Fig. 7), suggesting that the observed gene expression changes are unlikely due to direct effects of the viral RNA in the frontal cortex. On the other hand, our transcriptomic pathway analyses identified upregulation of tumor necrosis factor (TNF) and type I/II interferon response pathways in the frontal cortex of individuals with COVID-19 (Fig. 2d). Indeed, interferons and TNF have been implicated in brain aging and aging-induced cognitive decline6,13,14,15. Among individuals with COVID-19 with available peripheral cytokine data, we indeed observed increased TNF levels (3.5–24.2 pg ml−1 at 0–2 d before death in three individuals; values > 2.8 pg ml−1 are considered elevated; data were not available for the rest of the individuals). In line with our findings, a mouse model of SARS-CoV-2 infection exhibited elevation of pro-inflammatory cytokines in cerebrospinal fluid, including interferon gamma (IFN-γ) and TNF, in the absence of viral neuroinvasion16. To test whether TNF and type I/II interferons can modulate the expression of aging-regulated genes, we performed a transcriptomic analysis (RNA-seq) on human primary neurons treated with different doses of TNF, interferon beta (IFN-β) or IFN-γ. Interestingly, TNF and to a lesser extent IFN-γ and IFN-β increased the predicted aging index of cytokine-treated neurons, suggestive of the induction of an aging-like effect (Fig. 2e,f). We also found that cytokines upregulated the expression of aging-regulated genes that were upregulated in individuals with COVID-19 such as TRIM22, CHI3L1, C1S and IFITM1 and downregulated the expression of aging-regulated genes that were downregulated in individuals with COVID-19 such as CCND2, ACTR3B and EPHA5 (Supplementary Fig. 8). Taken together, our data suggest that COVID-19-induced TNF and type I/II interferons may lead to significant deteriorating effects in the brain in the absence of SARS-CoV-2 neuroinvasion.
Aging is a major risk factor for the development of cognitive deficits. Our results, together with previously reported residual cognitive deficits reported in recovered cases2, imply that aging-associated and cognitive decline-associated gene expression changes observed in individuals with COVID-19 may lead to increased rates of cognitive decline. Furthermore, we provide evidence that these aging-regulated gene expression changes may be mediated in part by circulating TNF and type I/II interferons, suggesting that acute management of severe COVID-19-induced inflammation may be neuroprotective. We recognize limitations in our study including: the variability in illness duration, the imperfect quality of several samples and the specificity of our findings due to COVID-19. Despite these constraints, our study was sufficiently powered to identify substantial transcriptome-wide changes in individuals with COVID-19. In addition to being larger than previously reported COVID-19 brain transcriptome studies11,12,17, our COVID-19 cases were matched by age and sex to uninfected controls, enabling the identification of aging-associated gene expression signatures in our samples. Furthermore, we included ICU/VENT uninfected samples as an additional independent control group to distinguish COVID-19 from other comorbidities requiring ICU monitoring and/or ventilation. The generalizability of our results to individuals who had mild COVID-19 or who recovered from COVID-19 remains to be determined. Given our findings, we advocate for neurological follow-up of individuals who recovered from COVID-19 and suggest potential clinical value in modifying risk factors to reduce the risk or delay the development of aging-related neurological pathologies and cognitive decline18.
Methods
Human brain tissues
This study complies with all relevant ethical regulations. Postmortem brain tissue specimens from individuals with COVID-19 (n = 22) were collected through a protocol for waived consent for the use of excess tissue approved by the Mass General Brigham Institutional Review Board (IRB protocol no. 2018P001724; controls and samples for the aging cohort were obtained as de-identified from the National Institutes of Health (NIH) NeuroBioBank, which does not require IRB approval; postmortem human brain research of de-identified samples is not considered human subjects research). Consent for autopsy was provided by the individuals’ next of kin or healthcare proxy according to Massachusetts state law (participant compensation was not applicable). All autopsies were performed at Brigham and Women’s Hospital (BWH) from 01 September 2020 to 31 December 2021 with pre-mortem or peri-mortem positive testing for SARS-CoV-2 by nasopharyngeal swab qPCR. Individuals in the COVID-19 group had no known psychiatric or neurological disorder (two individuals had history of previous stroke; Supplementary Table 1) with the exception of one person with epilepsy and another with Alzheimer’s disease. Tissues collected within a postmortem interval no more than 50 h were included, and this criterion was applied to all tissues used in our study, including controls and the aging cohort described below. Eligibility criteria included adults with pre-mortem or peri-mortem positive testing for SARS-CoV-2 by nasopharyngeal swab qPCR and no known psychiatric or neurological disorder (however, we included the individual with epilepsy and the individual with Alzheimer’s disease due to limited tissue availability) and postmortem interval no more than 50 h. Sample size was determined based on availability of tissues. At the time of autopsy, brains were sectioned coronally, and samples of middle frontal gyrus (alternating between left and right sides in the absence of gross abnormalities) were collected and frozen at −80 °C. Frozen middle/superior frontal gyrus (Brodmann area 8) controls were obtained from the NIH NeuroBioBank (the Harvard Brain Tissue Resource Center (HBTRC), the University of Miami’s Brain Endowment Bank and the University of Maryland Brain and Tissue Bank; n = 22). Controls (two of which had reported ventilator history) were selected to be age (within ± 2 years) and sex matched to a COVID-19 case and were categorized as unaffected controls with no known psychiatric or neurological condition in the NIH NeuroBioBank system. Each COVID-19 case was matched to one age-matched and sex-matched control; in the instance of similar ages and sex between COVID-19 and control cases within sequencing batches, cases and controls were grouped together for differential expression analysis (Supplementary Table 1). As one individual with COVID-19 had comorbid Alzheimer’s disease, we also included an age-matched and sex-matched uninfected individual with Alzheimer’s disease. As an additional independent group (ICU/VENT), we included frozen frontal cortex (Brodmann area 8) from uninfected individuals with a history of ventilator treatment obtained from the NIH NeuroBioBank (the University of Miami’s Brain Endowment Bank) and the Human Brain Collection Core at NIH, and frozen frontal cortex of uninfected individuals with ICU treatment, which were collected at BWH as described above for the COVID-19 cases (Supplementary Table 1). This control group was included as most individuals with COVID-19 received ICU/VENT treatment. Each case in the ICU/VENT control group was age matched (within ± 3 years; with three exceptions that were ±5–8 years, but were also included due to limited tissue availability) and sex matched to a COVID-19 case (n = 9). Thus, each COVID-19 case was matched to one ICU/VENT control when available. All control samples were collected before November 2019 and the start of the COVID-19 pandemic in the United States, with the exception of samples from three individuals with a history of ventilator or ICU support, who all had negative SARS-CoV-2 nasopharyngeal swab qPCR at the time of death and no reported COVID-19 history (a negative result for nucleocapsid serological/antibody test at the time of death was also available for one of those individuals) and thus were considered uninfected by SARS-CoV-2. For the aging cohort, frozen frontal cortex specimens (Brodmann area 8) from ten young (≤38 years old) and ten old (≥71 years old) individuals (aging cohort) were obtained from the NIH NeuroBioBank (the University of Maryland Brain and Tissue Bank, the HBTRC and the University of Miami’s Brain Endowment Bank; n = 10 per group; Supplementary Table 1). Samples analyzed in the aging cohort were collected before the COVID-19 outbreak in the United States and thus were considered uninfected by SARS-CoV-2.
Frozen tissue was processed using Biosafety Level 2+ procedures approved by the Beth Israel Deaconess Medical Center (BIDMC) Institutional Biosafety Committee. Brain tissues were homogenized using TRIzol (Thermo Fisher) reagent and RNA were extracted from tissues by phase separation. Total RNA was quantified by NanoDrop (DeNovix DS-11) and TapeStation 4200 (RNA Screen Tape; Agilent Technologies).
Definition of severe COVID-19
Severe COVID-19 was determined by meeting the NIH criteria for severe or critical illness (that is, peripheral capillary oxygen saturation < 94% on room air at sea level, the ratio of arterial oxygen partial pressure to fractional inspired oxygen < 300 mm Hg, a respiratory rate > 30 breaths per minute, or lung infiltrates > 50%; respiratory failure, septic shock and/or multiple organ dysfunction). Asymptomatic COVID-19 was determined by testing for SARS-CoV-2 by nasopharyngeal swab qPCR and a lack of respiratory or other typical symptoms observed in COVID-19.
Library construction and RNA-seq
A total of 450 ng RNA for the frontal cortex samples and 80 ng of total RNA for the human primary neurons was used for library preparation via the KAPA RNA HyperPrep kit with RiboErase (HMR; Roche, 08098131702) according to the manufacturer’s recommendations. Briefly, hybridization with hybridization oligonucleotides (HMR) was performed at 95 °C for 2 min followed by rRNA depletion using RNAse H, which was performed at 45 °C for 30 min. Following rRNA depletion cleanup via KAPA pure beads, DNase digestion was performed at 37 °C for 30 min followed by cleanup, RNA elution, fragmentation (6 min at 94 °C for samples with an RNA integrity number ≥ 7 or 5 min at 85 °C for samples with an RNA integrity number ≤ 7) and priming. First-strand and second-strand synthesis and A-tailing were performed according to the manufacturer’s recommendations. A total of 1.5 μM KAPA Unique Dual-Indexed (UDI; Roche, 8861919702) adaptors were ligated to the second-strand synthesis product in the presence of a ligation master mix in a reaction that was performed at 20 °C for 15 min. Following cleanup, all libraries underwent 10 (frontal cortex samples) or 13 (human primary neurons) cycles of amplification. Successful library production, quality control and quantification were assessed using TapeStation (High sensitivity D1000 Screen Tape; Agilent Technologies). Libraries were pooled (four runs in total) and subjected to NovaSeq 6000.
RT–qPCR
A total of 400 ng RNA from each frontal cortex sample was processed for cDNA via the SuperScript IV Reverse Transcriptase kit (Thermo Fisher Scientific, 18090050) according to the manufacturer’s instructions. All qPCR experiments were performed in a 384-well plate using LightCycler 480 SYBR Green (Roche, 4887352001) via a Roche LightCycler 480 II PCR system using Roche LightCycler 480 Software v1.5.1.62.
To assess the expression of SARS-CoV-2, primers against the SARS-CoV-2 nucleocapsid (N) gene (primer set nCOV_N1 (IDT no. 10007031; 5′-GACCCCAAAATCAGCGAAAT-3′ for forward and 10007032; 5′-TCTGGTTACTGCCAGTTGAATCTG-3′ for reverse primer) and primer set nCOV_N2 (IDT no. 10007033; 5′-TTACAAACATTGGCCGCAAA-3′ for forward and 10007034; 5′-GCGCGACATTCCGAAGAA-3′ for reverse primer)) and RPP30 (RNase P gene; RP; used for normalization; IDT no. 10006827; 5′-AGATTTGGACCTGCGAGCG-3′ for forward and no. 10006828; 5′-GAGCGGCTGTCTCCACAAGT-3′ for reverse primer) were synthesized (IDT) as recommended by the US Centers for Disease Control and Prevention. The 2019-nCoV_N_Positive Control RUO Plasmid (IDT no. 10006625) was included as a positive control. All other primers (ACTB (QT00095431), S100A8 (QT00226121), S100A9 (QT00018739), IFITM1 (QT00064246), MYL12A (QT01665741), GRIN3A (QT00043617), RHOBTB3 (QT00072611), CORO1A (QT00066997), SST (QT00004277), MAP2 (QT00057358) and NPTXR (QT00015701); QuantiTect Primer Assays) were purchased from Qiagen. ACTB was used for normalization. qPCR data were analyzed via the \(2^{{-\Delta\Delta}{{c}_{\rm{T}}}}\) method19 using Microsoft Excel 2016.
RNA-seq analysis
For assessment of SARS-CoV-2 genome alignment, reads were aligned to the SARS-CoV-2 reference genome (NCBI reference sequence NC_045512.2) using bowtie2 v2.2.9 with options ‘--X 1000 --no-mixed’. Analysis of RNA-seq of Calu-3-infected samples from Blanco-Melo et al.20 were included as positive controls with default bowtie2 parameters.
For assessment of differential gene expression, raw sequencing reads were aligned to a reference transcriptome generated from the Ensembl v104 human transcriptome with salmon v1.4.0 using options ‘--seqBias --useVBOpt --gcBias --posBias --numBootstraps 30 --validateMappings’. Length-scaled transcripts per million were acquired using the tximport v1.18.0 function, and log2 fold changes and FDRs were determined by DESeq2 v1.30.1 in R. t-SNE analysis was performed using Rtsne v0.15, with counts transformed by the variance stabilizing transformation function from DESeq2. Heat maps were generated with pheatmap v1.0.12 using counts produced by variance stabilizing transformation, with further scaling across samples. For the analysis of COVID-19 versus uninfected controls, age/sex matching (Supplementary Table 1) was used as a covariate.
Gene-set enrichment analysis
Signed −log10 FDRs from DESeq2 analyses were used to rank genes for GSEA via fgsea v1.16.0, filtering out genes with an FDR > 0.5. Public gene sets used for analyses were: GO Biological Processes (GO.db v3.12.1), Kyoto Encyclopedia of Genes and Genomes (KEGG; KEGGREST v1.30.1) and ReactomeDB (reactome.db v1.74.0) pathways to gene mappings from fgsea via the ‘reactomePathways’ function. For enrichment analyses, Ensembl gene IDs were matched with corresponding gene symbols and Entrez IDs via biomaRt v2.46.3.
Brain aging-regulated molecular signatures
Significant DEGs (FDR < 0.05) from our aging cohort transcriptome analysis were used to assess gene-set enrichment. In addition, we leveraged previously published aging-regulated DEG set data generated in five independent cohorts6,8,9,10,21,22. Lu et al. performed a broad-spectrum gene expression analysis (Affymetrix Human Genome U95Av2) of human prefrontal cortex from 30 individuals aged 26–106 years old and determined age-regulated genes based on a comparison of individuals ≤ 42 years old versus individuals ≥ 73 years old (n = 10–11 per group)6. Loerch et al. performed a genome-wide gene expression analysis (Affymetrix Human Genome U133plus 2.0) of human prefrontal cortex from individuals aged 24–94 years old and determined age-regulated genes based on a comparison of individuals ≤ 40 years old versus individuals ≥ 70 years old (n = 28; n = 13–15 per group)8. For three additional human cohorts, we used DEGs as determined by ref. 23 in which gene expression data from individuals aged 85+ years old were compared to gene expression data of younger individuals; in those reanalyses, Zullo et al. included only individuals with annotated normal cognitive function. Those additional three cohorts include: (1) the Gibbs et al. cohort21 in which gene expression data (Illumina HumanRef-8 Expression BeadChips) from the frontal cortex of 37 individuals were analyzed and DEGs estimated by comparing individuals aged 85+ years to individuals aged 55–80 years23; (2) the ROSMAP cohort, part of the Religious Orders Study (ROS) and Rush Memory and Aging Project (MAP; ROSMAP) at the Rush Alzheimer’s Disease Center9,10,24, in which RNA-seq data from the dorsolateral prefrontal cortex of 117 individuals were analyzed and DEGs estimated by comparing individuals aged 85+ years to individuals 70–80 years old23; and (3) the Common Mind Consortium (CMC) cohort22 in which RNA-seq data from the dorsolateral prefrontal cortex of 155 individuals were analyzed and DEGs estimated by comparing individuals aged 85+ years to individuals 60–80 years old23.
Molecular signatures in the frontal cortex associated with cognitive decline in humans
ROSMAP clinical data and aligned RNA-seq counts9,10 were downloaded from https://www.synapse.org/. A total of 633 (406 females; 227 males) transcriptomic profiles with corresponding MMSE scores were available; the median MMSE score (25) was used to stratify transcriptomic profiles as either good or poor cognition. All available cases (individuals with no cognitive impairment, individuals with mild cognitive impairment and individuals with Alzheimer’s or other dementia; with ages ranging between 67 and 95+ years) were included. DEGs were determined by DESeq2 (using sex as a covariate; age was not included as a covariate due to the narrow age distribution, absence of younger individuals and lack of precise age information for cases above 95 years of age) in R v4.0.4, and enrichment of COVID-19-dysregulated DEGs was performed by GSEA as described above.
Aging index
Due to the differences in transcriptome profiling methods used in this study (total RNA-seq versus poly-A capture or microarray used by other studies), which may bias our modeling approaches, we derived an ‘aging index’ using our aging cohort as a training set and the ‘control’ group of the COVID-19 cohort as a test set. The aging index was determined as the first principal component derived from a principal-component analysis of log2(transcripts per milllion + 1) genes via ‘prcomp’ in R, with feature selection determined by FDR < 5% from DESeq2 differential expression analysis. Varying the FDR cutoff for feature selection did not substantially affect the Pearson correlation of the test set; further model training was not performed due to the relatively small size of the cohort. Similarly transformed gene expression datasets derived from total RNA-seq (COVID cohort and cytokine treatments) as in our aging study were vector multiplied to the PC1 rotation scalars to yield predicted aging index scores.
The contribution of cerebrovascular disease (presence of atherosclerosis, arteriolosclerosis or cerebral hemorrhage) on the predicted aging index was determined using a linear regression model that also assessed the potential interaction of chronological age with cerebrovascular disease in individuals with COVID-19 using IBM SPSS Statistics 21.
Cell culture maintenance and treatments
Human neurons (ScienCell Research Laboratories, 1520-5) were thawed and cultured in neuronal medium (ScienCell, 1521) for 3 d on poly-d-lysine-coated plates before cell treatments. In total, 1 × 105 human neurons per well were plated in a 24-well plate. Cells were treated with IFN-β (PBL, 11415-1; 1 ng ml−1 or 0.1 ng ml−1), IFN-γ (BioLegend, 570204; 1 µg ml−1 or 0.1 µg ml−1), TNF (BioLegend, 570102; 100 ng ml−1 or 10 ng ml−1) or nuclease-free water (control) for 72 h. Cells were collected and RNA was extracted via TRIzol (Thermo Fisher).
Statistical analysis
No statistical methods were used to predetermine sample size. The experimental groups were not randomized. Where possible, samples were processed together using de-identified numbers (RNA-seq library preparation, reverse transcription before qPCR, and qPCR). For qPCR and RNA-seq analyses, blinding was not possible as all changes had to be matched with corresponding controls. For qPCR, when no gene expression was detected (no CT value determined), the CT value was set to 40 (maximum number of cycles) to perform a statistical analysis. qPCR data were analyzed with a two-tailed t-test and via GraphPad Prism 9. RNA-seq statistical analyses were performed in R (v4.0.4)25 as described above. Comparisons of predicted aging index (PC1) between two groups were performed using a two-tailed Welch’s t-test in R v4.0.4 and Bonferroni’s adjustment for multiple comparisons was applied. Linear regression was performed to assess the potential interaction of cerebrovascular disease with age on the predicted aging index in individuals with COVID-19 using IBM SPSS Statistics 21.0.
Reporting summary
Further information on research design is available in the Nature Portfolio Reporting Summary linked to this article.
Data availability
RNA-seq fastq files generated for this study are available through the Gene Expression Omnibus (GEO) under accession number GSE188847. Raw Calu-3.fastq RNA-seq files from Blanco-Melo et al. are available through the GEO (GSE147507) under accession numbers GSM4462348–GSM4462353 (ref. 20).
Aging gene lists used for gene enrichment analyses are available in Supplementary Table 4. Preprocessed gene expression datasets used to collate these gene lists were obtained from: Lu et al.6 (https://static-content.springer.com/esm/art%3A10.1038%2Fnature02661/MediaObjects/41586_2004_BFnature02661_MOESM5_ESM.xls); Loerch et al.8; and ROSMAP (Supplementary Table 1 of Zullo et al.), CMC (Supplementary Table 3 of Zullo et al.) and Gibbs (Supplementary Table 5 of Zullo et al.23) (https://static-content.springer.com/esm/art%3A10.1038%2Fs41586-019-1647-8/MediaObjects/41586_2019_1647_MOESM3_ESM.zip).
The SARS-CoV-2 genome was obtained from https://www.ncbi.nlm.nih.gov/nuccore/1798174254/. The Ensembl v104 human reference transcriptome was obtained from http://ftp.ensembl.org/pub/release-104/fasta/homo_sapiens/cdna/Homo_sapiens.GRCh38.cdna.all.fa.gz. GO (http://geneontology.org/) was queried from org.Hs.eg.db v3.12.0 in R. Reactome pathway annotations (https://reactome.org/) were obtained via the ‘reactomePathways’ command in R package fgsea (https://bioconductor.org/packages/release/bioc/html/fgsea.html). KEGG hsa pathway annotations (https://www.genome.jp/kegg/) were obtained using the KEGGREST v1.30.1 API in R (https://www.bioconductor.org/packages/release/bioc/html/KEGGREST.html).
Source data are provided with this study. Any other data are available from the authors upon reasonable request.
Code availability
R scripts used are deposited on GitHub at https://github.com/jonathandlee12/covid19-brain/.
References
Solomon, I. H. et al. Neuropathological features of COVID-19. N. Engl. J. Med. 383, 989–992 (2020).
Hampshire, A. et al. Cognitive deficits in people who have recovered from COVID-19. EClinicalMedicine 39, 101044 (2021).
Kas, A. et al. The cerebral network of COVID-19-related encephalopathy: a longitudinal voxel-based 18F-FDG-PET study. Eur. J. Nucl. Med. Mol. Imaging 48, 2543–2557 (2021).
Douaud, G. et al. SARS-CoV-2 is associated with changes in brain structure in UK Biobank. Nature 604, 697–707 (2022).
Silvin, A. et al. Elevated calprotectin and abnormal myeloid cell subsets discriminate severe from mild COVID-19. Cell 182, 1401–1418 (2020).
Lu, T. et al. Gene regulation and DNA damage in the ageing human brain. Nature 429, 883–891 (2004).
Boehm, M. & Slack, F. A developmental timing microRNA and its target regulate lifespan in C. elegans. Science 310, 1954–1957 (2005).
Loerch, P. M. et al. Evolution of the aging brain transcriptome and synaptic regulation. PLoS ONE 3, e3329 (2008).
Bennett, D. A., Schneider, J. A., Arvanitakis, Z. & Wilson, R. S. Overview and findings from the religious orders study. Curr. Alzheimer Res. 9, 628–645 (2012).
Bennett, D. A. et al. Overview and findings from the rush Memory and Aging Project. Curr. Alzheimer Res. 9, 646–663 (2012).
Yang, A. C. et al. Dysregulation of brain and choroid plexus cell types in severe COVID-19. Nature 595, 565–571 (2021).
Fullard, J. F. et al. Single-nucleus transcriptome analysis of human brain immune response in patients with severe COVID-19. Genome Med. 13, 118 (2021).
Hu, W. T. et al. CSF cytokines in aging, multiple sclerosis and dementia. Front. Immunol. 10, 480 (2019).
Deczkowska, A. et al. Mef2C restrains microglial inflammatory response and is lost in brain ageing in an IFN-I-dependent manner. Nat. Commun. 8, 717 (2017).
Belarbi, K. et al. TNF-α protein synthesis inhibitor restores neuronal function and reverses cognitive deficits induced by chronic neuroinflammation. J. Neuroinflammation 9, 23 (2012).
Fernandez-Castaneda, A. et al. Mild respiratory COVID can cause multi-lineage neural cell and myelin dysregulation. Cell 185, 2452–2468 (2022).
Gagliardi, S. et al. Detection of SARS-CoV-2 genome and whole transcriptome sequencing in frontal cortex of COVID-19 patients. Brain Behav. Immun. 97, 13–21 (2021).
Livingston, G. et al. Dementia prevention, intervention, and care: 2020 report of the Lancet Commission. Lancet 396, 413–446 (2020).
Schmittgen, T. D. & Livak, K. J. Analyzing real-time PCR data by the comparative CT method. Nat. Protoc. 3, 1101–1108 (2008).
Blanco-Melo, D. et al. Imbalanced host response to SARS-CoV-2 drives development of COVID-19. Cell 181, 1036–1045 (2020).
Gibbs, J. R. et al. Abundant quantitative trait loci exist for DNA methylation and gene expression in human brain. PLoS Genet. 6, e1000952 (2010).
Fromer, M. et al. Gene expression elucidates functional impact of polygenic risk for schizophrenia. Nat. Neurosci. 19, 1442–1453 (2016).
Zullo, J. M. et al. Regulation of lifespan by neural excitation and REST. Nature 574, 359–364 (2019).
De Jager, P. L. et al. A multi-omic atlas of the human frontal cortex for aging and Alzheimer’s disease research. Sci. Data 5, 180142 (2018).
R Core Team. R: A Language and Environment for Statistical Computing (R Foundation for Statistical Computing, 2018).
Acknowledgements
This work was supported by a grant from the National Institute on Aging (R01 AG058816) to F.J.S. The funder had no role in study design, data collection and analysis, decision to publish or preparation of the manuscript. We thank the NIH NeuroBioBank, the HBTRC, the University of Miami Brain Endowment Bank, the University of Maryland Brain and Tissue Bank and the Human Brain Collection Core at the NIH for providing control brain tissues. We thank S. Berretta, Director of the HBTRC, for advice on the selection of appropriate brain area controls and D. Davis and S. Marenco for helping with identification of cases with history of being on a ventilator. We thank the Beth Israel Deaconess Medical Center Institutional Biosafety Committee and R. Griffin for advice on Biosafety Level 2+ protocols, R. Ozdemir for advice on data analysis, V. Petkova for RNA-seq library quality control, I. Vlachos for sequencing assistance, T. Saxena for advice on library preparation, and the Biopolymers Facility at Harvard Medical School for sample quality-control and sequencing services.
Author information
Authors and Affiliations
Contributions
M.M. conceived of the idea and designed the study. M.M. prepared libraries for RNA-seq and performed qPCR experiments. J.D.L. performed bioinformatic analyses, performed in vitro work, and contributed to the experimental design of the in vitro studies. I.H.S. generated the relevant IRB protocol, collected COVID-19-infected and uninfected ICU control brain tissues, provided clinical annotations and determined the appropriate brain area for the controls. F.J.S. and M.M. supervised the study. M.M., J.D.L. and F.J.S. wrote the manuscript. All authors reviewed and edited the manuscript before submission.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Peer review
Peer review information
Nature Aging thanks Robert Reichard and the other, anonymous, reviewer(s) for their contribution to the peer review of this work.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Supplementary Information
Supplementary Figs. 1–8
Supplementary Tables 1–6
Supplementary Table 1. List of samples that were included in the study. COVID-19 cohort (a). Aging cohort (b). Supplementary Table 2. Lists of DEGs identified in the COVID-19 and aging cohorts. List of DEGs based on the following comparisons: COVID-19 versus age-matched/sex-matched control (a), COVID-19 versus uninfected ICU/VENT (b) and young-old (c). Supplementary Table 3. Results of pathway analysis. Complete lists of pathways enriched among COVID-19 versus age-matched/sex-matched control DEGs by GSEA (FDR < 0.05), identified by GO, Reactome and KEGG analyses (a–c). Complete lists of pathways enriched among COVID-19 versus uninfected ICU/VENT control DEGs by GSEA (FDR < 0.05), identified by GO (d). Complete lists of pathways enriched among old versus young DEGs by GSEA (FDR < 0.05), identified by GO (e). Supplementary Table 4. COVID-19 DEGs that are also deregulated in the aged human frontal cortex. List of significant COVID-19 DEGs (COVID-19 versus age-matched and sex-matched controls) overlapping with DEGs from each of the aging cohorts used in our analysis: this paper (a), Lu et al. (b), Loerch et al. (c), Gibbs cohort (d), ROSMAP cohort (e) and CMC cohort (f). List of aged frontal cortex DEGs from each cohort (g). Supplementary Table 5. List of DEGs between low and high MMSE scores (ROSMAP). List of DEGs associated with differences in cognition split by median MMSE score in the ROSMAP patient cohort. Supplementary Table 6. Lists of DEGs following in vitro cytokine treatment in human neurons. List of DEGs based on the following: IFN-β low dose (0.1 ng ml−1) (a), IFN-β high dose (1 ng ml−1) (b), IFN-γ Low dose (0.1 µg ml−1) (c), IFN-γ high dose (1 μg ml−1) (d), TNF low dose (10 ng ml−1) (e) and TNF high dose (100 ng ml−1) (f) versus untreated control. n = 3 per group.
Source data
Source Data Fig. 2
Predicted aging index values.
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Mavrikaki, M., Lee, J.D., Solomon, I.H. et al. Severe COVID-19 is associated with molecular signatures of aging in the human brain. Nat Aging 2, 1130–1137 (2022). https://doi.org/10.1038/s43587-022-00321-w
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/s43587-022-00321-w
This article is cited by
-
Resistance training's impact on blood biomarkers and cognitive function in older adults with low and high risk of mild cognitive impairment: a randomized controlled trial
European Review of Aging and Physical Activity (2024)
-
The neurobiology of SARS-CoV-2 infection
Nature Reviews Neuroscience (2024)
-
Seeing through the fog: a neuroendocrine explanation for post-COVID cognitive deficits
Nature Reviews Endocrinology (2024)
-
Proteomic and transcriptomic profiling of brainstem, cerebellum and olfactory tissues in early- and late-phase COVID-19
Nature Neuroscience (2024)
-
Using autopsies to dissect COVID-19 pathogenesis
Nature Microbiology (2023)
