
Abstract
Forest disturbance, including deforestation, is a major driver of global biodiversity decline. Identifying the underlying socioeconomic drivers can help guide interventions to halt biodiversity decline. Here, we quantified spatial overlaps between the distributions of 6164 globally threatened terrestrial vertebrate species and five major forest disturbance drivers at the global scale: commodity-driven deforestation, shifting agriculture, forestry, wildfire, and urbanization. We find that each driver has a distinct relative importance among species groups and geographic regions with, for example, the dominant disturbance drivers being forestry in northern regions and shifting agriculture in the tropics. Overall, shifting agriculture was more prevalent within threatened forest species’ ranges in the tropics, and some temperate nations. Our findings suggest that, globally, threatened forest species are exposed to a disproportional decrease in habitat area. Combining forest disturbance maps and species ranges can help evaluate agricultural landscape management and prioritize conservation efforts to reduce further biodiversity loss.
Similar content being viewed by others


Introduction
Across the globe, forest ecosystems are increasingly altered and fragmented by land uses, including agriculture, forestry, industry, and urban development1,2. Accordingly, forest disturbance, including deforestation and other modifications, is one of the main causes of biodiversity decline3,4,5. Since forest disturbance reduces species’ habitat availability, the spatial relationships between forest disturbance and biodiversity have received considerable empirical attention, especially for forest-dwelling threatened species6. For conservation, understanding the spatial co-distribution of drivers underlying forest disturbances and species ranges is also critical because the effectiveness of political and practical conservation measures is likely to differ among drivers7, and the relative importance of indirect drivers differs from species to species. Community-based forestry and agriculture, for example, may require local-scale responses such as building capacities for biodiversity-friendly operations and improving livelihoods8,9. Commodity-driven agriculture and plantations, in contrast, may require large-scale actions and transboundary agreements on land use to regulate the balance between ecosystem provisioning services and global demands that come through the supply chains10,11,12,13. These examples suggest that the quantification of spatial overlaps between biodiversity and drivers underlying forest disturbances is essential, not only for assessing its effects on biodiversity, but also for identifying appropriate conservation strategies. In particular, the effectiveness of area-based conservation measures would depend on the overlap patterns. Protected areas are one of the most common area-based conservation measures14 and are basically effective for preventing forests from disturbances. However, for forests that have been already largely disturbed, different mechanisms are likely to be essential to regulate and manage the land use by incorporating conservation as one of the targets in addition to the major driver of the land use. The other effective area-based conservation measures (OECMs) that have drawn international attention in their implementation recently15 would be one of such promising measures. Recent work by Curtis et al.2 that clarified the spatial distribution of major drivers of forest disturbances at the global scale provides a promising opportunity to examine the spatial co-distribution of drivers of forest disturbances and species ranges as well as how this knowledge might affect the conservation strategies.
Here we examined how drivers of forest disturbances unevenly overlap with biodiversity at the global scale. Specifically, we overlaid global-scale forest disturbance drivers on the ranges of 6164 globally threatened forest species of mammals, birds, reptiles, and amphibians. For each species, we spatially discriminated the proportion of five major drivers of forest disturbances: commodity-driven deforestation, shifting agriculture, forestry, wildfire, and urbanization, as well as forest with zero or minor loss, following the classification of Curtis et al.2. We then examined how the proportion of forest disturbances is changed within each species range compared with that in the baseline. Since we targeted threatened forest species, we hypothesized the proportion of forest with zero or minor loss within the species ranges would be low. We also hypothesized that the bias in the spatial overlaps would depend on geographic region and species characteristics. We tested these hypotheses by an analysis that examined how the quantified proportions of drivers within species ranges are related to the species’ forest habitat specificity, taxonomic class, IUCN Red List status16, and geographic distribution to generalize the global patterns of effects of the forest disturbance drivers on biodiversity. Finally, we hypothesized that species with higher proportions of drivers within their range would be more threatened by drivers that have operated for longer periods. We tested this hypothesis by analyzing how the current proportions of drivers within species ranges have affected the temporal patterns of changes in the species’ IUCN Red List status among the multiple assessment times.
Results
Global patterns of driver proportions
To illustrate the differences in the effects of the major driver proportions, we quantified two types of driver proportions: original and within-species-range (our proxy for a driver’s proportional effect on each species). To calculate the original driver proportion we overlaid subregion polygons on the global driver map2 and summarized the proportions of drivers within each subregion polygon. We then calculated driver proportions within each species range and averaged them among species (see Methods).
We found a dramatic increase in the effect of shifting agriculture on species (49.19% for the mean driver proportion within species ranges) from baseline at the global scale (vs. 13.83% for the original driver proportion; Fig. 1; Supplementary Table 1). To examine regional patterns, we assessed seven subregions: Russia/Asia, South-East Asia, Oceania, Europe, Africa, North America, and Latin America, following Curtis et al.2. Distinct increases in shifting agriculture were found in four subregions: Latin America (from 30.75% to 62.58%), Africa (from 49.85% to 75.18%), South-East Asia (from 26.05% to 39.02%), and Oceania (from 5.03% to 22.28%). Striking increases in forestry were found in the other three regions: Europe (from 24.29% to 58.44%), North America (from 21.66% to 42.62%), and Russia/Asia (from 22.22% to 44.66%). Note that this map-based pattern was robust even after the accuracy of driver classification was accounted for (Supplementary Fig. 1; Supplementary Table 1).

Values in parentheses are the numbers of threatened species analyzed.
Differences in driver proportions among species groups and regions
Through Dirichlet regression analysis17, we found contrasting differences in the effect of IUCN threatened status (near threatened, NT; vulnerable, VU; endangered, EN; or critically endangered, CR), forest habitat specialization (forest specialist or generalist; see Methods for definition), taxonomic class (mammal, bird, reptile, or amphibian), and subregion (Russia/Asia, South-East Asia, Oceania, Europe, Africa, North America, or Latin America) on the proportion of the five forest disturbance drivers and forest with zero or minor loss within each species range at the global scale (Fig. 2; Supplementary Fig. 2). Specifically, forest habitat specialists tend to be exposed to forest disturbance irrespective of driver type, and CR species (all drivers) and EN species (shifting agriculture and wildfire) have higher proportions of disturbance drivers within their distribution ranges than NT species (Fig. 2; Supplementary Table 2). Among taxonomic classes, amphibians are exposed to larger proportions of drivers of all types, and reptiles have a higher proportion of shifting agriculture as a disturbance driver. By subregion, each driver had distinct differences: species in South-East Asia, Oceania, Africa, and Latin America, for example, tend to have higher proportions of shifting agriculture when Russia/Asia is considered as the control (Supplementary Fig. 2; Supplementary Table 2). The patterns in the map-based proportions reported above were unchanged or even strengthened by considering driver classification accuracy (Supplementary Table 2; also see Methods for details).

a–e forest habitat specialization (generalist or specialist); f–j threatened status on IUCN Red List (NT, near threatened; VU, vulnerable; EN, endangered; CR, critically endangered); k–o taxonomic class (mammal, bird, reptile, amphibian). Center line, median; box limits, upper and lower quartiles; whiskers, 1.5× interquartile range. In the Dirichlet regression, all categorical variables were considered at the same time as explanatory variables (i.e., multiple regressions), and each category on the left (i.e., generalist, NT, mammal) was treated as the control. For the IUCN status, NT was selected as the control since it is the lowest risk category. Note that effect of subregion was also considered in the regression (Supplementary Fig. 2). Asterisks represent statistically significant positive effect of the category against the control category (*P < 0.05, **P < 0.01, ***P < 0.001) in the Dirichlet regression using the proportion of zero/minor forest loss as baseline (see also Supplementary Fig. 2; Supplementary Table 2).
Driver effects at a national scale
To highlight the differences in effects of the major driver proportions among nations and of shifting agriculture, in particular, we quantified two types of driver proportions at the national level: driver proportion in each nation (original driver proportion) and driver proportion within species ranges in each nation (driver proportion within species range). The original driver proportion is calculated by overlaying country polygons on the global driver map2; the driver proportion within species ranges is calculated by overlaying species range polygons intersected by national borders on the global driver map.
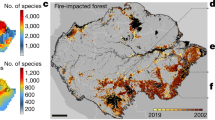
Commodity-driven deforestation, shifting agriculture, and forestry are the strongest drivers and, as suggested by Curtis et al.2, their proportions are distinctly different among nations (Supplementary Figs. 3, S3, S4). Specifically, the original driver proportion for shifting agriculture tends to dominate nations in the tropics (Fig. 3a). Moreover, the mean driver proportion within species ranges over all threatened species revealed that the dominance of shifting agriculture was increased in all tropical nations and in some temperate and boreal nations (Fig. 3b, c). Drastic increases in shifting agriculture proportions were found for forest habitat specialists, especially in central Africa (Fig. 3d); for amphibians in several nations in central and northern Africa (Fig. 3e); and for critically endangered species in a few nations in Latin America and central Africa (Fig. 3f). The map-based driver proportions at the national level are provided in an open repository (https://doi.org/10.6084/m9.figshare.19471001). Because the effects of classification accuracy on the driver proportions may not be trivial at national scale (Supplementary Figs. S5, S6), we have also provided the accuracy-adjusted mean and 95% confidence interval of all driver proportions.

a Original shifting agriculture proportion calculated on a 10-km × 10-km-grid forest disturbance driver map2. b Shifting agriculture proportion within all species ranges (see Methods for details). c Difference between proportions (b minus a). d Difference between proportions with forest habitat specialist range and all species. e Difference between proportions with amphibian species range and all species. (f) Difference between proportions with critically endangered (CR) species range and all species range. Gray: no available data.
Effects of current driver proportions on temporal change in IUCN Red List status
In tropical subregions (South-East Asia, Africa, and Latin America), the current proportions of three major drivers—commodity-driven deforestation, shifting agriculture, and forestry—within a species range had comparable negative associations with the change in IUCN Red List status of 2129 forest species (Supplementary Table 3) from 1988 to 2018 quantified by moving a 10-year window by 1-year steps to cover the period (see Methods). Note that each Red List category was transformed to a category weight prior to the analysis (i.e., 1.0 for Least Concern (LC), 0.8 for NT, 0.6 for VU, 0.4 for EN, 0.2 for CR, and 0 for EX), such that negative change indicates worsening of the status (see Methods).
We also found that the proportions of all three drivers significantly interacted with year trends in the change of Red List status but in different ways (Fig. 4). For commodity-driven deforestation, the negative impact on the status change of the driver’s larger proportion tended to remain throughout the period (Fig. 4a). For shifting agriculture, however, the larger proportion brought more negative effects on the status change especially earlier in the period (Fig. 4b). The pattern for forestry was similar to that of shifting agriculture, but the negative impacts of the larger proportion was more limited to the beginning of the period (Fig. 4c). The patterns were robust regardless of the width of time window, as we obtained qualitatively the same patterns when we used 7- and 13-year windows instead of a 10-year window (Supplementary Table 3).

a commodity-driven deforestation, b shifting agriculture, and c forestry. The change in species’ Red List status was measured as a mean status change within a 10-year window sequentially moved to cover 1988–2018. Positive and a negative changes represent improving and worsening of the status, respectively. Black, orange, and red solid lines represent status change predicted by a generalized linear mixed model (GLMM; see Methods for details) when proportion of focal disturbance driver was assumed to be mean, 10th percentile, and 90th percentile of observed proportions, respectively, keeping all other drivers at the mean level. Each polygon represents a 90% prediction interval based on fixed effects of the GLMM, and the color of each polygon corresponds to the prediction shown as a solid line.
Discussion
Our results show that the effects of the forest disturbance drivers on biodiversity are likely to be different from those simply expected from the baseline proportions of the forest disturbance drivers if we take into account the threatened species’ distributions. The amount of forest habitat is a primary factor for species diversity of many taxa, including mammals, amphibians, reptiles, birds, insects, and plants18. Indeed, our results revealed that threatened forest species have been exposed to a disproportional decrease in their habitat amount globally (i.e., lower proportions of forest with no or minor loss in all regions when species ranges were considered). Although this finding may be intuitive as population size and/or species range are part of the criteria in the IUCN assessment19, the detected pattern supports the validity of our approach of combining a forest disturbance map and species ranges for evaluating the impact of forest disturbances on threatened species. Moreover, we found that the dominant drivers differ among regions: the proportion of forestry, for example, increased in northern regions such as North America and Europe, whereas that of shifting agriculture increased in tropical regions when threatened species’ distributions were considered. These facts indicate although several influential international schemes for conservation have been implemented for regulating forestry20,21, different mechanisms aiming to directly tackle the over land use for local agriculture may increase their importance when we consider conservation in tropical regions. Our findings suggest that the social and economic drivers underlying the forest disturbance that impacts biodiversity differ among regions or nations, and it is important to establish specific conservation strategies in order to be effective.
Based on the findings, we further emphasize that the combinations of multiple interacting drivers are likely to vary among regions. For example, the frequency and extent of stand-replacing natural disturbances such as wildfires have clearly been magnified by climate change, particularly in the Northern Hemisphere (e.g.,22). After such natural disturbances, societal demand for timber and/or pest reduction compels forest managers to ‘salvage’ timber by logging before it deteriorates, a common practice even in locations otherwise exempt from conventional green-tree harvesting, such as national parks or wilderness areas23. Thus, salvage logging clearly mediates the interaction between disturbances by forestry and wildfires and is likely to further affect biodiversity under climate change. Especially in regions where infrastructure (e.g., irrigation systems) has not been well developed, unpredictable changes in precipitation due to climate change was reported to increase forest disturbance by unregulated increases of agricultural land use24. Such regions largely overlapped with regions where shifting agriculture was identified as a dominant disturbance driver for threatened species in this study. Moreover, species themselves shift their ranges in response to climate change25, which would also shift major disturbance drivers and influential interactions of drivers to which the species are exposed, given the region-specific driver patterns. These examples clearly suggest the necessity to understand both the region-specific interrelations among multiple drivers and species’ responses for better prediction of land-use change and thus its effects on biodiversity.
Shifting agriculture was the most dominant driver in all tropical regions corresponding to the recent estimates suggesting that the cover of regenerating secondary forest is increasing worldwide26. We demonstrated that this tendency is more drastic especially within the range of threatened species. The effect of shifting agriculture per unit area might be more limited than that of commodity-driven deforestation, which permanently alters forests into other land uses, since habitat structure might recover as the forest vegetation regenerates to a secondary state following the abandonment of the small clearings. However, ample evidence shows that many types of agricultural activities significantly degrade the conservation value of primary forest, especially in the tropics27, which often recovers very slowly if ever28 with the loss of irreplaceable conservation values. Therefore, given the wide areas of dominance of shifting agriculture across all tropical regions, its effect is likely to be pervasive. Consistently, our results show that species extinction risk (i.e., IUCN Red List status) is positively related to the proportional coverage of shifting agriculture (Fig. 2). In addition, as expected, a larger current proportion of shifting agriculture within a species range worsens the change rate in IUCN Red List status of the species (Fig. 4b). Furthermore, the effect is anticipated to be magnified for forest specialists because they are exposed to larger proportions of shifting agriculture than are forest generalist (Fig. 2), and they are also reported to recover more slowly than do forest habitat generalists27,28.
A guideline for forest restoration suggested that appropriately sized landscapes should contain ≥40% forest cover (higher percentages are likely needed in the tropics), with about 10% in a very large forest patch and the remaining 30% in many evenly dispersed smaller patches and semi-natural wooded elements (e.g., vegetation corridors)29. Importantly, the guideline also suggests that the patches should be embedded in a high-quality matrix. Although younger secondary forest cannot be a substitute for pristine forest until 50 years or more after a disturbance, it can help to improve the quality of matrix in agricultural landscapes30. Indeed, we show that the negative impacts of shifting agriculture and forestry on IUCN status change have improved over time (Fig. 4b, c), presumably corresponding to the forest regenerating and recovery process. In contrast, the pattern of commodity-driven deforestation, a land use accompanied with permanent forest loss, showed a prolonged negative impact on IUCN status change (Fig. 4a). Notably, whether regenerating forests can move towards a highly diverse and structurally complex state or towards a state of low to intermediate levels of biodiversity and structural complexity depends on the amount of remaining intact mature forest in the landscape29. Therefore, a promising direction for future research would be to develop our analysis further to include spatiotemporal relationships among mature forest remnants, secondary forests, disturbance drivers, and threatened species populations.
For conserving the core patches of mature forests, the establishment of protected areas (PAs) is one of the most effective legal measures that has been widely used to regulate land use for biodiversity31. On the other hand, for improving matrix quality, balancing conservation and use of the ecosystem would be critically important; shifting agriculture, for example, causes forest degradation, but it also contributes to food supply chains sourced from smallholder farmers and to food security of local communities8. In fact, establishing mechanisms for managing biodiversity-friendly landscapes has been intensively discussed recently, given the large potential influence of these landscapes on conservation32. These mechanisms include setting an international target on OECMs15. Our finding of a disproportional decrease in forest proportions with minor or no loss within species ranges supports the urgency of the discussion. At the same time, our results highlight an opportunity because large portions of the disturbed forests for threatened species are dominated by shifting agriculture at the global scale, especially in the tropics. As suggested above, if manged properly, such landscapes can still retain or improve functions as essential habitats and/or matrix for a variety of forest-dwelling species. Our analytical method provides a tool set to identify and prioritize areas where such attempts are urgently needed.
Global demands for natural resources and ecosystem services drive land use in forests33 and thus affect biodiversity. Therefore, connecting the supply chains to the five major drivers of forest disturbance and their spatial overlaps with biodiversity is essential to inform how we should regulate and design material flows from forest ecosystems to keep them sustainable by minimizing the effects on biodiversity. Existing studies examining the impacts of resource consumption on biodiversity through supply chains of various sectors have often been assessed at the country scale (e.g.,12), partly because the availability of statistics needed to estimate material flows in supply chains is usually limited at finer (i.e., subnational) scales (but see34). We believe that our study provides the first basis for filling the resolution gap between trade statistics and local biodiversity effects by identifying patterns of the local co-occurrence of biodiversity and the forest disturbance drivers that can be directly linked to resource production at the national scale. Note, however, that downscaling a remotely sensed global data set into finer scales inevitably propagates errors and biases which include both those in the original maps and those in the processed data produced by analyses. Thus, preparation of more high-resolution data sets is essential, especially for disturbance drivers and threatened species’ distributions in our case, to keep the errors and biases at a reasonable level at focal spatial scales.
The effectiveness of area-based conservation measures to regulate land use for conservation including PAs and OECMs also depends strongly on social and ecosystem conditions. For example, a few studies show that the effectiveness of PAs in halting or slowing forest disturbances depends on PA characteristics such as size and history, as well as on the management entities such as subnational governments or indigenous peoples35,36,37. Moreover, there has been no attempt to elucidate whether PAs and OECMs are effective at regulating supply chains as a supply-side measure by balancing resource production, ecosystem services for local communities, and biodiversity conservation; to tackle this issue, it will be necessary to conduct extensive analyses integrating spatial and temporal patterns of biodiversity, forest loss, its drivers, and material flows in global food supply chains. Though it is challenging and beyond the scope of this paper, solving this issue is urgent and raises a promising opportunity for future research.
Methods
Global data of species range and forest disturbances
We obtained range maps for amphibians, reptiles, and mammals from the IUCN Red List16 and those for birds from BirdLife International38. For each species, we used only range polygons where presence was not classified as ‘Extinct’ and origin was classified as ‘Native’ or ‘Reintroduced’. The dataset consisted of 28 139 species, 7752 of which are listed as threatened: those classified as near threatened (NT), vulnerable (VU), endangered (EN), or critically endangered (CR) on the Red List. We classified species as ‘non-forest’ or ‘forest’ according to the IUCN Red List habitat classification data16. We treated species using only forest habitat as forest specialists, those using forest habitat and at least one other habitat type as forest generalists, and those not using forest at all as non-forest species. We excluded non-threatened (including ‘Data deficient’ and ‘Extinct in the wild’) species, non-forest species, and species that have no overlaps with the forest driver maps (explained in the next paragraph) from the subsequent analysis, resulting in 6164 threatened forest species: 1227 mammals, 1855 birds, 881 reptiles, and 2201 amphibians.
We calculated two types of driver proportions for comparison and mapping: original and within species range. The original driver proportion is based solely on the global driver map2, whereas the within-species-range driver proportion is calculated by overlaying species range polygons on the global driver map. Specifically, within each species range, we calculated the areas of each of five major drivers of forest disturbances2: (i) commodity-driven deforestation, defined as the long-term, permanent conversion of forest and shrubland to a non-forest land use such as agriculture (including oil palm), mining, or energy infrastructure; (ii) shifting agriculture, defined as small- to medium-scale forest and shrubland conversion for agriculture that is later abandoned and followed by subsequent forest regrowth; (iii) forestry, defined as large-scale forestry operations occurring within managed forests and tree plantations with evidence of forest regrowth in subsequent years; (iv) wildfire, defined as large-scale forest loss resulting from the burning of forest vegetation with no visible human conversion or agricultural activity afterwards; and (v) urbanization, defined as forest and shrubland conversion for the expansion and intensification of existing urban centers, as well as forest areas with zero or minor loss. Note that grid cells with no forest cover have no data in the driver map. We then took the mean of each driver proportion among species as a proxy for the driver’s effect on biodiversity (within species range). Note that all of these calculations can be conducted at national and subregion scales as well as at the global scale, because we also took summaries at the national scale by overlaying the nation polygons on the species range polygons and the global driver maps.
Subregional classification accuracy and error matrix of the drivers were provided in the original study2 and Supplementary Table 6 therein. To consider the classification uncertainties, the original driver proportion was recalculated using the user’s accuracy (proportion of correctly classified areas within the total classified areas as a driver) of each subregion randomly drawn from its 95% confidence interval. A “misclassified area” according to the drawn accuracy was redistributed to the other four drivers proportionally to their error rates in the classification error matrix thereby we considered the full error matrix for each subregion including both omission and commission errors. Similarly, we recalculated the within-species-range driver proportion. We applied the same procedure for driver proportions within each species range and then took the mean of each of the recalculated driver proportions among species. We repeated the recalculations of the accuracy-adjusted original driver proportion and the within-species-range driver proportion 1000 times at global, subregion, and nation scales. We then used the 1000 replicated data to quantify uncertainties due to the classification errors in the results of subsequent analyses.
Temporal change of IUCN Red List status
To examine temporal change in Red List status in relation to driver proportions, we collected the assessment history records of the Red List status of each species between 1988 and 2018 using the R package rredlist39. The IUCN Red List categories were transformed into an index by applying the RLI calculation method40. Specifically, each Red List category was transformed to a category weight (1.0 for LC, 0.8 for NT, 0.6 for VU, 0.4 for EN, 0.2 for CR, and 0 for EX). We calculated the mean temporal change of the weight for each species by moving a 10-year window sequentially from the 1988–1997 window to the 2009–2018 window. Within each time window, the difference in the category weights between the nearest neighboring assessments were calculated and divided by the interval between the assessments to obtain annual change. When there were more than two assessments within a time window, we took the mean of annual changes as a proxy for the temporal change for the time window. We excluded species that did not have multiple assessments in any time window. We then extracted species in subregions where shifting agriculture is a major driver (i.e., South-East Asia, Africa, and Latin America). Consequently, we obtained temporal changes of IUCN Red List status for 2129 threatened forest species: 633 mammals, 900 birds, 98 reptiles, and 498 amphibians. In order to evaluate the sensitivity of the result to the width of time window, we conducted the same calculation for 7-year and 13-year time windows.
Statistical analysis
To examine how the areas of the five forest disturbance drivers as proportions of total forest area differed among regions and species groups, we conducted a Dirichlet regression analysis17. Each species was treated as a sample, and the proportions of the five drivers and zero/minor loss within the species range at the global scale are response variables that are assumed to follow a Dirichlet distribution. We used the ‘alternative’ parameterization17, where mean and precision parameters are explicitly modeled. We assumed that the mean of each proportion is a function of four categorical variables (i.e., explanatory variables): forest habitat specialization (forest specialist or generalist), status on IUCN Red List (NT, VU, EN, or CR), taxonomic class (mammal, bird, reptile, or amphibian), and subregion (Russia/Asia, South-East Asia, Oceania, Europe, Africa, North America, and Latin America). We used the same classification of subregions as used in Curtis et al.2; for species whose range contains more than two subregions, the subregion that has the largest area in the species range was assigned. In Dirichlet regression, since a separate model for each category (driver proportion here) is over-determined, only C−1 models are fitted, and the Cth category is treated as a baseline category and modeled implicitly as the ‘residual’ category after the others are accounted for. We set the proportion of zero/minor forest loss as the baseline category. We estimated the parameters of the Dirichlet regression using the DirichletReg v. 0.741 package for R software with its default settings, except as explained above. In addition, to consider the uncertainties due to the driver classification errors, we conducted the same Dirichlet regression analysis using each of the 1000 replicated data sets that incorporate the classification accuracy, and thus we obtained 1000 regression results. We then summarized the 1000 replications for each regression coefficient to calculate its mean and 95% confidence interval.
We analyzed the relationships between the temporal change of IUCN Red List status and the current driver proportions in South-East Asia, Africa, and Latin America by using a generalized linear mixed model (GLMM). In the GLMM, temporal change of IUCN Red List status within time windows for each species was the response variable and proportion of major drivers in the subregions (i.e., commodity-driven deforestation, shifting agriculture, and forestry) within the species range were used as explanatory variables. In addition, we included the initial year of each time window and square of the year (centralized before squared) as explanatory variables to control temporal trends across the focal period (1988–2018). We also considered the interaction between year and each driver proportion. The species’ subregion (South-East Asia, Africa, and Latin America) and species ID nested in taxonomic class (mammal, bird, reptile, or amphibian) were incorporated as random intercepts. The GLMM was estimated using the lmer function42 in the R software package.
Data availability
Source data used for generating figures were provided in supplementary information and deposited to a repository (https://doi.org/10.6084/m9.figshare.19471097).
References
Furukawa, T. et al. Forest harvest index: accounting for global gross forest cover loss of wood production and an application of trade analysis. Glob. Ecol. Conserv. 4, 150–159 (2015).
Curtis, P. G., Slay, C. M., Harris, N. L., Tyukavina, A. & Hansen, M. C. Classifying drivers of global forest loss. Science 361, 1108–1111 (2018).
Bradshaw, C. J., Sodhi, N. S. & Brook, B. W. Tropical turmoil: a biodiversity tragedy in progress. Front. Ecol. Environ. 7, 79–87 (2009).
Laurance, W. F. Have we overstated the tropical biodiversity crisis? Trends Ecol. Evol. 22, 65–70 (2007).
Pimm, S. L. & Raven, P. Biodiversity: extinction by numbers. Nature 403, 843 (2000).
Betts, M. G. et al. Global forest loss disproportionately erodes biodiversity in intact landscapes. Nature 547, 441 (2017).
IPBES. Global Assessment Report on Biodiversity and Ecosystem Services of the Intergovernmental Science-Policy Platform on Biodiversity and Ecosystem Services. (IPBES secretariat, 2019).
Sunderland, T. & Vasquez, W. Forest conservation. Rights, and diets: untangling the issues. Front. For. Glob. Change 3, 29 (2020).
Wren-Lewis, L., Becerra-Valbuena, L. & Houngbedji, K. Formalizing land rights can reduce forest loss: experimental evidence from Benin. Sci. Adv. 6, eabb6914 (2020).
Nishijima, S. et al. Evaluating the impacts of wood production and trade on bird extinction risks. Ecol. Indic. 71, 368–376 (2016).
Moran, D. & Kanemoto, K. Identifying species threat hotspots from global supply chains. Nat. Ecol. Evol. 1, 0023 (2017).
Lenzen, M. et al. International trade drives biodiversity threats in developing nations. Nature 486, 109–112 (2012).
Ferrer Velasco, R., Köthke, M., Lippe, M. & Günter, S. Scale and context dependency of deforestation drivers: Insights from spatial econometrics in the tropics. PloS One 15, e0226830 (2020).
UNEP-WCMC & IUCN. Protected Planet: The World Database on Protected Areas (WDPA)s. https://www.protectedplanet.net/en. (2021).
IUCN-WCPA Task Force on OECMs. Guidelines for Recognising and Reporting Other Effective Area-based Conservation Measures. IUCN, Switzerland.(2019).
IUCN. The IUCN Red List of Threatened Species, Version 2017-2. (2017).
Douma, J. C. & Weedon, J. T. Analysing continuous proportions in ecology and evolution: a practical introduction to beta and Dirichlet regression. Methods Ecol. Evol. 10, 1412–1430 (2019).
Watling, J. I. et al. Support for the habitat amount hypothesis from a global synthesis of species density studies. Ecol. Lett. 23, 674–681 (2020).
IUCN, Species Survival Commision. IUCN Red List categories and criteria: version 3.1. Prepared by the IUCN Species Survival Commission (2001).
Gardner, T. A. et al. A framework for integrating biodiversity concerns into national REDD+ programmes. Biol. Conserv. 154, 61–71 (2012).
Johansson, T., Hjältén, J., de Jong, J. & von Stedingk, H. Environmental considerations from legislation and certification in managed forest stands: a review of their importance for biodiversity. For. Ecol. Manag. 303, 98–112 (2013).
Holden, Z. A. et al. Decreasing fire season precipitation increased recent western US forest wildfire activity. Proc. Natl Acad. Sci. USA. 115, E8349–E8357, https://doi.org/10.1073/pnas.1802316115 (2018).
Thorn, S. et al. Impacts of salvage logging on biodiversity: a meta-analysis. J. Appl. Ecol. 55, 279–289, https://doi.org/10.1111/1365-2664.12945 (2018).
Zaveri, E., Russ, J. & Damania, R. Rainfall anomalies are a significant driver of cropland expansion. Proc. Natl Acad. Sci. 117, 10225–10233 (2020).
Pecl, G. T. et al. Biodiversity redistribution under climate change: Impacts on ecosystems and human well-being. Science 355, eaai9214 (2017).
Hansen, M. C. et al. High-resolution global maps of 21st-century forest cover change. Science 342, 850–853 (2013).
Gibson, L. et al. Primary forests are irreplaceable for sustaining tropical biodiversity. Nature 478, 378–+, https://doi.org/10.1038/nature10425 (2011).
Acevedo‐Charry, O. & Aide, T. M. Recovery of amphibian, reptile, bird and mammal diversity during secondary forest succession in the tropics. Oikos 128, 1065–1078 (2019).
Arroyo-Rodriguez, V. et al. Designing optimal human-modified landscapes for forest biodiversity conservation. Ecol. Lett. 23, 1404–1420, https://doi.org/10.1111/ele.13535 (2020).
Rozendaal, D. M. A. et al. Biodiversity recovery of Neotropical secondary forests. Sci. Adv. 5, https://doi.org/10.1126/sciadv.aau3114 (2019).
Laurance, W. F. et al. Averting biodiversity collapse in tropical forest protected areas. Nature 489, 290–+, https://doi.org/10.1038/nature11318 (2012).
Melo, F. P., Arroyo-Rodríguez, V., Fahrig, L., Martínez-Ramos, M. & Tabarelli, M. On the hope for biodiversity-friendly tropical landscapes. Trends Ecol. Evol. 28, 462–468 (2013).
Hoang, N. T. & Kanemoto, K. Mapping the deforestation footprint of nations reveals growing threat to tropical forests. Nat. Ecol. Evol. 5, 845–853 (2021).
Wakiyama, T., Lenzen, M., Kadoya, T., Takeuchi, Y. & Nansai, K. Forest tax payment responsibility from the forest service footprint perspective. Environ. Sci. Technol. 55, 3165–3174 (2021).
Geldmann, J. et al. A global analysis of management capacity and ecological outcomes in terrestrial protected areas. Conserv. Lett. 11, https://doi.org/10.1111/conl.12434 (2018).
Gray, C. L. et al. Local biodiversity is higher inside than outside terrestrial protected areas worldwide. Nat. Commun. 7, https://doi.org/10.1038/ncomms12306 (2016).
Blankespoor, B., Dasgupta, S. & Wheeler, D. Protected areas and deforestation: new results from high-resolution panel data. Nat. Resour. Forum 41, 55–68, https://doi.org/10.1111/1477-8947.12118 (2017).
BirdLife International. <http://www.birdlife.org/datazone/home>
Chamberlain, S. rredlist:‘IUCN’Red List Client. 0.4. 0. https://github.com/ropensci/rredlist (2017).
Bubb, P. et al. IUCN Red List index: Guidance for national and regional use. Version 1.1. (IUCN, 2009).
Maier, M. J. DirichletReg: Dirichlet regression for compositional data in R. (2014).
Kuznetsova, A., Brockhoff, P. B. & Christensen, R. H. B. lmerTest package: tests in linear mixed effects models. J. Stat. Softw. 82, 1–26 (2017).
Acknowledgements
T.K., Y.S., and Y.T. were partly supported by the Moonshot Agriculture, Forestry and Fisheries Research and Development Program MS509.
Author information
Authors and Affiliations
Contributions
T.K. and K.N. designed and led the research. T.K., Y.T., and Y.S. conducted the analysis. T.K., Y.T., Y.S., and K.N. wrote the manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Peer review
Peer review information
Communications Earth & Environment thanks Dolors Armenteras, Eric Bullock and the other, anonymous, reviewer(s) for their contribution to the peer review of this work. Primary Handling Editors: Clare Davis.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Kadoya, T., Takeuchi, Y., Shinoda, Y. et al. Shifting agriculture is the dominant driver of forest disturbance in threatened forest species’ ranges. Commun Earth Environ 3, 108 (2022). https://doi.org/10.1038/s43247-022-00434-5
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s43247-022-00434-5
This article is cited by
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
