
Abstract
Pancreatic beta cells synthesize and secrete the neurotransmitter GABA (γ-aminobutyric acid) as a paracrine and autocrine signal to help regulate hormone secretion and islet homeostasis. Islet GABA release has classically been described as a secretory-vesicle-mediated event. Yet, a limitation of the hypothesized vesicular GABA release from islets is the lack of expression of a vesicular GABA transporter in beta cells. Consequentially, GABA accumulates in the cytosol. Here, we provide evidence that the human beta cell effluxes GABA from a cytosolic pool in a pulsatile manner, imposing a synchronizing rhythm on pulsatile insulin secretion. The volume regulatory anion channel, functionally encoded by LRRC8A or Swell1, is critical for pulsatile GABA secretion. GABA content in beta cells is depleted and secretion is disrupted in islets from patients with type 1 and type 2 diabetes, suggesting that loss of GABA as a synchronizing signal for hormone output may correlate with diabetes pathogenesis.
This is a preview of subscription content, access via your institution
Access options
Access Nature and 54 other Nature Portfolio journals
Get Nature+, our best-value online-access subscription
$29.99 / 30 days
cancel any time
Subscribe to this journal
Receive 12 digital issues and online access to articles
$119.00 per year
only $9.92 per issue
Buy this article
- Purchase on Springer Link
- Instant access to full article PDF
Prices may be subject to local taxes which are calculated during checkout






Similar content being viewed by others
Data availability
The unique biological materials used in the manuscript are available from the corresponding authors upon reasonable request with the exception of those materials that the authors obtained via a materials transfer agreement that prohibits transfer to third parties; these include the GABA biosensor cells (obtainable from K. Kaupmann, Novartis Institute for BioMedical Research, Basal, Switzerland), LRRC8A−/− MIN6 cells and LRRC8Afl/fl mice (obtainable from R. Sah, Washington University in St. Louis, MO, USA), and NPY-pHluorin (obtainable from H. Gaisano, University of Toronto, Toronto, ON, Canada). Other requests for materials should be addressed to corresponding author A.C. or E.A.P. Source data for Figs. 1–6 and Extended Data Figs. 1, 4, and 5 are provided with the paper. The data that support the findings of this study are available from the corresponding authors upon reasonable request.
References
Kanaani, J. et al. Compartmentalization of GABA synthesis by GAD67 differs between pancreatic beta cells and neurons. PLoS ONE 10, e0117130 (2015).
Rorsman, P. et al. Glucose-inhibition of glucagon secretion involves activation of GABAA-receptor chloride channels. Nature 341, 233–236 (1989).
Xu, E. et al. Intra-islet insulin suppresses glucagon release via GABA-GABAA receptor system. Cell Metab. 3, 47–58 (2006).
Bailey, S. J., Ravier, M. A. & Rutter, G. A. Glucose-dependent regulation of γ-aminobutyric acid (GABAA) receptor expression in mouse pancreatic islet ɑ-cells. Diabetes 56, 320–327 (2007).
Fiorina, P. GABAergic system in β-cells: from autoimmunity target to regeneration tool. Diabetes 62, 3674–3676 (2013).
Tian, J. et al. γ-Aminobutyric acid regulates both the survival and replication of human β-cells. Diabetes 62, 3760–3765 (2013).
Purwana, I. et al. GABA promotes human β-cell proliferation and modulates glucose homeostasis. Diabetes 63, 4197–4205 (2014).
Tian, J., Dang, H. & Kaufman, D. L. Combining antigen-based therapy with GABA treatment synergistically prolongs survival of transplanted β-cells in diabetic NOD mice. PLoS ONE 6, e25337 (2011).
Tian, J., Dang, H., Middleton, B. & Kaufman, D. L. Clinically applicable GABA receptor positive allosteric modulators promote β-cell replication. Sci. Rep. 7, 374 (2017).
Prud’homme, G. J., Glinka, Y. & Wang, Q. Immunological GABAergic interactions and therapeutic applications in autoimmune diseases. Autoimmun. Rev. 14, 1048–1056 (2015).
Untereiner, A. et al. GABA promotes β-cell proliferation, but does not overcome impaired glucose homeostasis associated with diet-induced obesity. FASEB J. 33, 3968–3984 (2019).
He, S. et al. Rapamycin/GABA combination treatment ameliorates diabetes in NOD mice. Mol. Immunol. 73, 130–137 (2016).
Feng, A. L. et al. Paracrine GABA and insulin regulate pancreatic alpha cell proliferation in a mouse model of type 1 diabetes. Diabetologia 60, 1033–1042 (2017).
Ben-Othman, N. et al. Long-term GABA administration induces alpha cell-mediated beta-like cell neogenesis. Cell 168, 73–85 e11 (2017).
Li, J. et al. Artemisinins target GABAA receptor signaling and impair ɑ cell identity. Cell 168, 86–100 e115 (2017).
Ackermann, A. M., Moss, N. G. & Kaestner, K. H. GABA and artesunate do not induce pancreatic α-to-β cell transdifferentiation in vivo. Cell Metab. 28, 787–792.e3 (2018).
van der Meulen, T. et al. Artemether does not turn α cells into β cells. Cell Metab. 27, 218–225.e214 (2018).
Barragan, A., Weidner, J. M., Jin, Z., Korpi, E. R. & Birnir, B. GABAergic signalling in the immune system. Acta Physiol. (Oxf.) 213, 819–827 (2015).
Mendu, S. K., Bhandage, A., Jin, Z. & Birnir, B. Different subtypes of GABA-A receptors are expressed in human, mouse and rat T lymphocytes. PLoS ONE 7, e42959 (2012).
Jin, Z., Mendu, S. K. & Birnir, B. GABA is an effective immunomodulatory molecule. Amino Acids 45, 87–94 (2013).
Bjurstom, H. et al. GABA, a natural immunomodulator of T lymphocytes. J. Neuroimmunol. 205, 44–50 (2008).
Bhat, R. et al. Inhibitory role for GABA in autoimmune inflammation. Proc. Natl Acad. Sci. USA 107, 2580–2585 (2010).
Tian, J. et al. Gamma-aminobutyric acid inhibits T cell autoimmunity and the development of inflammatory responses in a mouse type 1 diabetes model. J. Immunol. 173, 5298–5304 (2004).
Kim, J. et al. Differential expression of GAD65 and GAD67 in human, rat, and mouse pancreatic islets. Diabetes 42, 1799–1808 (1993).
Kanaani, J., Patterson, G., Schaufele, F., Lippincott-Schwartz, J. & Baekkeskov, S. A palmitoylation cycle dynamically regulates partitioning of the GABA-synthesizing enzyme GAD65 between ER-Golgi and post-Golgi membranes. J. Cell Sci. 121, 437–449 (2008).
Braun, M. et al. Regulated exocytosis of GABA-containing synaptic-like microvesicles in pancreatic β-cells. J. Gen. Physiol. 123, 191–204 (2004).
Jin, H. et al. Demonstration of functional coupling between gamma-aminobutyric acid (GABA) synthesis and vesicular GABA transport into synaptic vesicles. Proc. Natl Acad. Sci. USA 100, 4293–4298 (2003).
Reetz, A. et al. GABA and pancreatic beta-cells: colocalization of glutamic acid decarboxylase (GAD) and GABA with synaptic-like microvesicles suggests their role in GABA storage and secretion. EMBO J. 10, 1275–1284 (1991).
Braun, M. et al. Corelease and differential exit via the fusion pore of GABA, serotonin, and ATP from LDCV in rat pancreatic β cells. J. Gen. Physiol. 129, 221–231 (2007).
Soltani, N. et al. GABA exerts protective and regenerative effects on islet beta cells and reverses diabetes. Proc. Natl Acad. Sci. USA 108, 11692–11697 (2011).
Braun, M. et al. γ-aminobutyric acid (GABA) is an autocrine excitatory transmitter in human pancreatic β-cells. Diabetes 59, 1694–1701 (2010).
Bansal, P. et al. GABA coordinates with insulin in regulating secretory function in pancreatic INS-1 β-cells. PLoS ONE 6, e26225 (2011).
Smismans, A., Schuit, F. & Pipeleers, D. Nutrient regulation of gamma-aminobutyric acid release from islet beta cells. Diabetologia 40, 1411–1415 (1997).
Wang, C., Mao, R., Van de Casteele, M., Pipeleers, D. & Ling, Z. Glucagon-like peptide-1 stimulates GABA formation by pancreatic beta-cells at the level of glutamate decarboxylase. Am. J. Physiol. Endocrinol. Metab. 292, E1201–E1206 (2007).
Garry, D. J., Sorenson, R. L. & Coulter, H. D. Ultrastructural localization of gamma amino butyric acid immunoreactivity in B cells of the rat pancreas. Diabetologia 30, 115–119 (1987).
Semyanov, A., Walker, M. C. & Kullmann, D. M. GABA uptake regulates cortical excitability via cell type-specific tonic inhibition. Nat. Neurosci. 6, 484–490 (2003).
Garry, D. J., Sorenson, R. L., Elde, R. P., Maley, B. E. & Madsen, A. Immunohistochemical colocalization of GABA and insulin in beta-cells of rat islet. Diabetes 35, 1090–1095 (1986).
Kanaani, J., Diacovo, M. J., El-Husseini Ael, D., Bredt, D. S. & Baekkeskov, S. Palmitoylation controls trafficking of GAD65 from Golgi membranes to axon-specific endosomes and a Rab5a-dependent pathway to presynaptic clusters. J. Cell Sci. 117, 2001–2013 (2004).
Kanaani, J. et al. A combination of three distinct trafficking signals mediates axonal targeting and presynaptic clustering of GAD65. J. Cell Biol. 158, 1229–1238 (2002).
Chaudhry, F. A. et al. The vesicular GABA transporter, VGAT, localizes to synaptic vesicles in sets of glycinergic as well as GABAergic neurons. J. Neurosci. 18, 9733–9750 (1998).
Jenstad, M. & Chaudhry, F. A. The amino acid transporters of the glutamate/GABA-glutamine cycle and their impact on insulin and glucagon secretion. Front. Endocrinol. (Lausanne) 4, 199 (2013).
Gammelsaeter, R. Glycine, GABA and their transporters in pancreatic islets of Langerhans: evidence for a paracrine transmitter interplay. J. Cell Sci. 117, 3749–3758 (2004).
Cabrera, O. et al. Glutamate is a positive autocrine signal for glucagon release. Cell Metab. 7, 545–554 (2008).
Rodriguez-Diaz, R. et al. Alpha cells secrete acetylcholine as a non-neuronal paracrine signal priming beta cell function in humans. Nat. Med. 17, 888–892 (2011).
Rodriguez-Diaz, R. et al. Real-time detection of acetylcholine release from the human endocrine pancreas. Nat. Protoc. 7, 1015–1023 (2012).
Franek, M. et al. The heteromeric GABA-B receptor recognizes G-protein ɑ subunit C-termini. Neuropharmacology 38, 1657–1666 (1999).
Pagano, A. et al. C-terminal interaction is essential for surface trafficking but not for heteromeric assembly of GABAB receptors. J. Neurosci. 21, 1189–1202 (2001).
Tengholm, A. & Gylfe, E. Oscillatory control of insulin secretion. Mol. Cell. Endocrinol. 297, 58–72 (2009).
Allen, N. J., Káradóttir, R. & Attwell, D. Reversal or reduction of glutamate and GABA transport in CNS pathology and therapy. Pflugers Arch. 449, 132–142 (2004).
Richerson, G. B. & Wu, Y. Dynamic equilibrium of neurotransmitter transporters: not just for reuptake anymore. J. Neurophysiol. 90, 1363–1374 (2003).
Tian, N. et al. The role of the synthetic enzyme GAD65 in the control of neuronal γ-aminobutyric acid release. Proc. Natl Acad. Sci. USA 96, 12911–12916 (1999).
Patel, A. B., de Graaf, R. A., Martin, D. L., Battaglioli, G. & Behar, K. L. Evidence that GAD65 mediates increased GABA synthesis during intense neuronal activity in vivo. J. Neurochem. 97, 385–396 (2006).
Wang, C., Ling, Z. & Pipeleers, D. Comparison of cellular and medium insulin and GABA content as markers for living β-cells. Am. J. Physiol. Endocrinol. Metab. 288, E307–E313 (2005).
Baron, M. et al. A single-cell transcriptomic map of the human and mouse pancreas reveals inter- and intra-cell population structure. Cell Syst. 3, 346–360.e4 (2016).
Wang, Y. J. et al. Single-cell transcriptomics of the human endocrine pancreas. Diabetes 65, 3028–3038 (2016).
Grun, D. et al. De novo prediction of stem cell identity using single-cell transcriptome data. Cell Stem Cell 19, 266–277 (2016).
Tomi, M., Tajima, A., Tachikawa, M. & Hosoya, K. Function of taurine transporter (Slc6a6/TauT) as a GABA transporting protein and its relevance to GABA transport in rat retinal capillary endothelial cells. Biochim. Biophys. Acta 1778, 2138–2142 (2008).
Yahara, T., Tachikawa, M., Akanuma, S., Kubo, Y. & Hosoya, K. Amino acid residues involved in the substrate specificity of TauT/SLC6A6 for taurine and γ-aminobutyric acid. Biol. Pharm. Bull. 37, 817–825 (2014).
del Amo, E. M., Urtti, A. & Yliperttula, M. Pharmacokinetic role of L-type amino acid transporters LAT1 and LAT2. Eur. J. Pharm. Sci. 35, 161–174 (2008).
Lee, S. et al. Channel-mediated tonic GABA release from glia. Science 330, 790–796 (2010).
Voss, F. K. et al. Identification of LRRC8 heteromers as an essential component of the volume-regulated anion channel VRAC. Science 344, 634–638 (2014).
Lutter, D., Ullrich, F., Lueck, J. C., Kempa, S. & Jentsch, T. J. Selective transport of neurotransmitters and modulators by distinct volume-regulated LRRC8 anion channels. J. Cell Sci. 130, 1122–1133 (2017).
Syeda, R. et al. LRRC8 proteins form volume-regulated anion channels that sense ionic strength. Cell 164, 499–511 (2016).
Qiu, Z. et al. SWELL1, a plasma membrane protein, is an essential component of volume-regulated anion channel. Cell 157, 447–458 (2014).
Planells-Cases, R. et al. Subunit composition of VRAC channels determines substrate specificity and cellular resistance to Pt-based anti-cancer drugs. EMBO J. 34, 2993–3008 (2015).
Kinard, T. A. & Satin, L. S. An ATP-sensitive Cl− channel current that is activated by cell swelling, cAMP, and glyburide in insulin-secreting cells. Diabetes 44, 1461–1466 (1995).
Kang, C. et al. SWELL1 is a glucose sensor regulating β-cell excitability and systemic glycaemia. Nat. Commun. 9, 367. (2018).
Stuhlmann, T., Planells-Cases, R. & Jentsch, T. J. LRRC8/VRAC anion channels enhance β-cell glucose sensing and insulin secretion. Nat. Commun. 9, 1974 (2018).
Zhang, Y. et al. SWELL1 is a regulator of adipocyte size, insulin signalling and glucose homeostasis. Nat. Cell Biol. 19, 504–517 (2017).
Thorens, B. et al. Ins1 Cre knock-in mice for beta cell-specific gene recombination. Diabetologia 58, 558–565 (2015).
Korol, S. V. et al. Functional characterization of native, high-affinity GABAA receptors in human pancreatic β cells. EBioMedicine 30, 273–282 (2018).
Chang, Y., Ghansah, E., Chen, Y., Ye, J. & Weiss, D. S. Desensitization mechanism of GABA receptors revealed by single oocyte binding and receptor function. J. Neurosci. 22, 7982–7990 (2002).
Ekholm, R., Ericson, L. E. & Lundquist, I. Monoamines in the pancreatic islets of the mouse. Subcellular localization of 5-hydroxytryptamine by electron microscopic autoradiography. Diabetologia 7, 339–348 (1971).
Hutton, J. C., Peshavaria, M. & Tooke, N. E. 5-Hydroxytryptamine transport in cells and secretory granules from a transplantable rat insulinoma. Biochem. J. 210, 803–810 (1983).
Kennedy, R. T., Huang, L., Atkinson, M. A. & Dush, P. Amperometric monitoring of chemical secretions from individual pancreatic beta-cells. Anal. Chem. 65, 1882–1887 (1993).
Barbosa, R. et al. Real time electrochemical detection of 5-HT/insulin secretion from single pancreatic islets: effect of glucose and K+ depolarization. Biochem. Biophys. Res. Commun. 228, 100–104 (1996).
Barbosa, R. et al. Control of pulsatile 5-HT/insulin secretion from single mouse pancreatic islets by intracellular calcium dynamics. J. Physiol. 510(Pt 1), 135–143 (1998).
Huang, Y. J. et al. Mouse taste buds use serotonin as a neurotransmitter. J. Neurosci. 25, 843–847 (2005).
Almaca, J. et al. Human beta cells produce and release serotonin to inhibit glucagon secretion from alpha cells. Cell Rep. 17, 3281–3291 (2016).
Dishinger, J. F., Reid, K. R. & Kennedy, R. T. Quantitative monitoring of insulin secretion from single islets of Langerhans in parallel on a microfluidic chip. Anal. Chem. 81, 3119–3127 (2009).
Deeney, J. T., Bränström, R., Corkey, B. E., Larsson, O. & Berggren, P. O. 3H-serotonin as a marker of oscillatory insulin secretion in clonal β-cells (INS-1). FEBS Lett. 581, 4080–4084 (2007).
Makhmutova, M., Liang, T., Gaisano, H., Caicedo, A. & Almaca, J. Confocal imaging of neuropeptide Y-pHluorin: a technique to visualize insulin granule exocytosis in intact murine and human islets. J. Vis. Exp. 127, e56089 (2017).
Almaca, J. et al. Spatial and temporal coordination of insulin granule exocytosis in intact human pancreatic islets. Diabetologia 58, 2810–2818 (2015).
Phelps, E. A. et al. Aberrant accumulation of the diabetes autoantigen GAD65 in Golgi membranes in conditions of ER stress and autoimmunity. Diabetes 65, 2686–2699 (2016).
Kass, I. et al. Cofactor-dependent conformational heterogeneity of GAD65 and its role in autoimmunity and neurotransmitter homeostasis. Proc. Natl Acad. Sci. USA 111, E2524–E2529 (2014).
Martin, D. L. & Rimvall, K. Regulation of γ-aminobutyric acid synthesis in the brain. J. Neurochem. 60, 395–407 (1993).
Chessler, S. D., Simonson, W. T., Sweet, I. R. & Hammerle, L. P. Expression of the vesicular inhibitory amino acid transporter in pancreatic islet cells: distribution of the transporter within rat islets. Diabetes 51, 1763–1771 (2002).
Thomas-Reetz, A. C. & De Camilli, P. A role for synaptic vesicles in non-neuronal cells: clues from pancreatic beta cells and from chromaffin cells. FASEB J. 8, 209–216 (1994).
Wendt, A. et al. Glucose inhibition of glucagon secretion from rat alpha-cells is mediated by GABA released from neighboring beta-cells. Diabetes 53, 1038–1045 (2004).
Braun, M. et al. Corelease and differential exit via the fusion pore of GABA, serotonin, and ATP from LDCV in rat pancreatic β cells. J. Gen. Physiol. 129, 221–231 (2007).
Jo, J., Choi, M. Y. & Koh, D. S. Beneficial effects of intercellular interactions between pancreatic islet cells in blood glucose regulation. J. Theor. Biol. 257, 312–319 (2009).
Koeslag, J. H., Saunders, P. T. & Terblanche, E. A reappraisal of the blood glucose homeostat which comprehensively explains the type 2 diabetes mellitus-syndrome X complex. J. Physiol. 549, 333–346 (2003).
Braun, M. et al. Voltage-gated ion channels in human pancreatic β-cells: electrophysiological characterization and role in insulin secretion. Diabetes 57, 1618–1628 (2008).
Gassmann, M. & Bettler, B. Regulation of neuronal GABA(B) receptor functions by subunit composition. Nat. Rev. Neurosci. 13, 380–394 (2012).
Farrant, M. & Nusser, Z. Variations on an inhibitory theme: phasic and tonic activation of GABA(A) receptors. Nat. Rev. Neurosci. 6, 215–229 (2005).
Braun, M., Ramracheya, R. & Rorsman, P. Autocrine regulation of insulin secretion. Diabetes Obes. Metab. 14(Suppl. 3), 143–151 (2012).
Taneera, J. et al. γ-Aminobutyric acid (GABA) signalling in human pancreatic islets is altered in type 2 diabetes. Diabetologia 55, 1985–1994 (2012).
Campbell-Thompson, M. et al. Network for Pancreatic Organ Donors with Diabetes (nPOD): developing a tissue biobank for type 1 diabetes. Diabetes Metab. Res. Rev. 28, 608–617 (2012).
Pugliese, A. et al. The Juvenile Diabetes Research Foundation Network for Pancreatic Organ Donors with Diabetes (nPOD) Program: goals, operational model and emerging findings. Pediatr. Diabetes 15, 1–9 (2014).
Phelps, E. A. et al. Advances in pancreatic islet monolayer culture on glass surfaces enable super-resolution microscopy and insights into beta cell ciliogenesis and proliferation. Sci. Rep. 7, 45961 (2017).
Codazzi, F. et al. Synergistic control of protein kinase Cγ activity by ionotropic and metabotropic glutamate receptor inputs in hippocampal neurons. J. Neurosci. 26, 3404–3411 (2006).
Bolte, S. & Cordelieres, F. P. A guided tour into subcellular colocalization analysis in light microscopy. J. Microsc. 224, 213–232 (2006).
L. Lun, A. T., Bach, K. & Marioni, J. C. Pooling across cells to normalize single-cell RNA sequencing data with many zero counts. Genome Biol. 17, 75 (2016).
Dvoryanchikov, G., Huang, Y. A., Barro-Soria, R., Chaudhari, N. & Roper, S. D. GABA, its receptors, and GABAergic inhibition in mouse taste buds. J. Neurosci. 31, 5782–5791 (2011).
Cabrera, O. et al. The unique cytoarchitecture of human pancreatic islets has implications for islet cell function. Proc. Natl Acad. Sci. USA 103, 2334–2339 (2006).
Rasouli, M. Basic concepts and practical equations on osmolality: biochemical approach. Clin. Biochem. 49, 936–941 (2016).
Zandy, S. L., Doherty, J. M., Wibisono, N. D. & Gonzales, R. A. High sensitivity HPLC method for analysis of in vivo extracellular GABA using optimized fluorescence parameters for o-phthalaldehyde (OPA)/sulfite derivatives. J. Chromatogr. B 1055–1056, 1–7 (2017).
Fernandez, N. A., Liang, T. & Gaisano, H. Y. Live pancreatic acinar imaging of exocytosis using syncollin-pHluorin. Am. J. Physiol. Cell Physiol. 300, C1513–C1523 (2011).
Chang, Y. C. & Gottlieb, D. I. Characterization of the proteins purified with monoclonal antibodies to glutamic acid decarboxylase. J. Neurosci. 8, 2123–2130 (1988).
Hampe, C. S. et al. A novel monoclonal antibody specific for the N-terminal end of GAD65. J. Neuroimmunol. 113, 63–71 (2001).
Acknowledgements
This work was funded by the Intramural Research Program of UF’s Wertheim College of Engineering and J. Crayton Pruitt Family Department of Biomedical Engineering (E.A.P.), the Intramural Research Program of EPFL’s School of Life Sciences (S.B.), the Diabetes Research Institute Foundation, NIH grant nos. R56DK084321 (A.C.), R01DK084321 (A.C.) and R01DK106009 (R.S.), the NIDDK-supported Human Islet Research Network (HIRN, RRID:SCR_014393; https://hirnetwork.org; grant no. UC4DK104208 (E.A.P)), a JDRF award (grant no. 31-2008-416) to the European Consortium for Islet Transplantation (ECIT) Islets for Basic Research Program, a JDRF Faculty Transition Award (grant no. 1-FAC-2017-367-A-N) (E.A.P.), a JDRF Advanced Postdoctoral Fellowship (grant no. 3-APF-2014-208-A-N) (E.A.P.), a Whitaker International Program Postdoctoral Scholarship (E.A.P.), The Shepard Broad Foundation (E.A.P.), the Swedish Research Council (P.-O.B.), the Novo Nordisk Foundation (P.-O.B.), the Family Erling-Persson Foundation (P.-O.B.), the Stichting af Jochnick Foundation (P.-O.B.), the American Diabetes Association (grant no. 1-18-IBS-229) (R.S.) and the Canadian Institutes for Health Research (grant nos. PJT-159741 and PJT-148652) (H.Y.G.).
Human pancreatic islets were provided by the NIDDK-funded Integrated Islet Distribution Program (IIDP) at City of Hope, NIH grant no. 2UC4DK098085, and the JDRF-funded IIDP Islet Award Initiative (E.A.P.). This research was performed with the support of the Network for Pancreatic Organ donors with Diabetes (nPOD; RRID:SCR_014641), a collaborative type 1 diabetes research project sponsored by JDRF (grant no. 5-SRA-2018-557-Q-R) and The Leona M. & Harry B. Helmsley Charitable Trust (grant no. 2018PG-T1D053). The content and views expressed are the responsibility of the authors and do not necessarily reflect the official view of nPOD. Organ Procurement Organizations (OPO) partnering with nPOD to provide research resources are listed at http://www.jdrfnpod.org/for-partners/npod-partners/. The work with human pancreatic sections and islets was also made possible by the Human Islet Cell Processing Facility at the Diabetes Research Institute (University of Miami) and ECIT.
We wish to extend our thanks to the following individuals: D. Bosco and T. Berney, University of Geneva, and L. Piemonti, San Raffaele Scientific Institute, Milan, for human islets through ECIT; C. Mathews, University of Florida, and R. Bottino, Institute of Cellular Therapeutics, Allegheny Health Network, Pittsburgh, Pennsylvania, for human islets through the nPOD Islet Isolation Program; K. Kaupmann, Novartis Institutes for BioMedical Research, Switzerland, for providing the genetically modified GABA biosensor CHO cells; and S. D. Roper for conceptual input on the biosensor cell approach and for critically reading the manuscript; J. Hubbell and M. Swartz, EPFL and University of Chicago, for support; C. Rancourt, University of Florida, for mouse colony management; and M. Pasquier, K. Johnson, B. Benjamin, D. Garcia, P. Parente, A. Arzu, L. Barash, M. Formoso, R. Arrojo e Drigo and A. Tamayo for technical assistance.
Author information
Authors and Affiliations
Contributions
E.A.P. and S.B. conceived and carried out subcellular studies of GABA-ergic components in islet cells. D.M. and A.C. conceived and identified GABA release from islets in pulses and pioneered the biosensor cell technique for analyzing the dynamics of islet GABA release. E.A.P. conceived and identified the role of VRAC and TauT in GABA release and uptake. D.W.H. and E.A.P. analyzed the genetic models for LRRC8A−/− MIN6 cells, βc-LRRC8A−/− murine islets and knock-down LRRC8A-shRNA human islets. D.M., D.W.H. and E.A.P. performed experiments to detect GABA, taurine and serotonin/insulin secretion. J.M. and J.A. performed hormone assay experiments and ELISAs. J.A. conducted NPY-pHluorin experiments to measure exocytosis. H.Y.G. generated adeno-NPY-pHluorin vectors. R.S. generated genetic models for LRRC8A−/− MIN6 cells and LRRC8Afl/fl murine islets. C.K. isolated and shipped LRRC8Afl/fl murine islets. M.W.B. isolated rodent islets and performed western blot analyses. C.C. prepared cultures of primary rat hippocampal neurons. P.C.S. performed bioinformatics analysis. R.N. and F.L. isolated human islets for research. E.A.P., D.M., D.W.H., J.A., C.C., R.M.D. and R.R.-D. collected, analyzed and quantified immunohistochemical data. P.-O.B. provided critical equipment, reagents, expertise and support. D.M., D.W.H., S.B., A.C. and E.A.P. designed the study, analyzed data and wrote the paper. All authors discussed the results and commented on the manuscript.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Peer review information Primary Handling Editor: Ana Mateus.
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Extended data
Extended Data Fig. 1 Delta cells in human islets contain GABA and express GAD65; taurine content is preserved in diabetic islets.
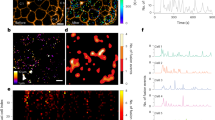
a-b. Islets in a non-diabetic human pancreas immunostained for GABA and somatostatin (a); or GAD65 and somatostatin (b). GABA and GAD65 are present in somatostatin producing human delta cells. Scale bar 50 µm. Right panels show higher magnification views of the boxed region showing channels for: (a) (1) GABA only, (2) somatostatin only, and (3) GABA and somatostatin; (b) (1) GAD65 only, (2) somatostatin only, and (3) GAD65 and somatostatin. Images are representative of data plotted in Figures 1b and 1j. Scale bar 20 µm. c. Quantification of taurine mean fluorescence intensity (MFI) per islet in confocal images of human pancreas sections from non-diabetic (n = 21 islets, 6 donors), type 2 diabetic (n = 24 islets, 8 donors), and type 1 diabetic donors (n = 24 islets, 8 donors). Background (BKGD) indicates average taurine MFI in acinar tissue outside of the islet. One-way ANOVA: ND vs. T2D (*P = 0.0430), ND vs. T1D (ns, P = 0.6667). Center line indicates the mean. d. Human pancreas sections immunostained for taurine, insulin, and glucagon from a non-diabetic, type 2 diabetic and, type 1 diabetic donor. Left panels show insulin and glucagon channels, while right panels show taurine channel from the same image. Images are representative of the dataset plotted in panel c. Scale bars 50 µm. e. Representative confocal image of a monolayer of human islet endocrine cells showing immunostaining for GAD65, insulin, and glucagon (left panel) and GAD65 alone (right panel). Images are representative of 3 human islet preparations. Scale bar 20 µm.
Extended Data Fig. 2 VGAT expression is concentrated in delta cells of human islets and alpha cells of rat islets; GABA colocalizes with GAD65 and VGAT in synaptic vesicles in neurons.
a-b. Islets in a non-diabetic human pancreas (a) and rat pancreas (b) immunostained for VGAT, insulin, glucagon, somatostatin, and pancreatic polypeptide. VGAT is absent in most beta cells but present in somatostatin producing human delta cells and glucagon producing rat alpha cells. Results are representative of the dataset plotted in Figure 1b. Scale bar 50 µm. c. Rat hippocampal neuron immunostained for GABA and the GABA biosynthesizing enzyme GAD65. Scale bar 10 µm. Right panels show higher magnification views of the boxed region showing channels for: (1) GABA only; (2) GAD65 only; (3) GABA and GAD65. GAD65 and GABA colocalize in vesicles. Results are representative of n = 3 rat neuron preparations. Scale bar 5 µm. d. Rat hippocampal neuron immunostained for GAD65 and the vesicular GABA transporter VGAT, which is present in synaptic vesicle membranes. Scale bar 10 µm. Right panels show higher magnification views of the boxed region showing channels for: (1) GAD65 only; (2) VGAT only; (3) GAD65 and VGAT. GAD65 and VGAT colocalize in synaptic vesicles. Results are representative of n = 3 rat neuron preparations. Scale bar 5 µm.
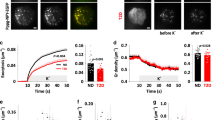
Extended Data Fig. 3 Characterization of GABA biosensor cells for detecting GABA released from human islets.
a. Schematic and image of the GABA biosensor cell assay setup (left panel). Biosensor cells consist of CHO cells stably expressing the heteromeric GABAB receptor (GABAB R1b and GABAB R2) and the G-protein α subunit, Gαqo5 to allow for GABA detection by intracellular Ca2+ mobilization (Δ[Ca2+]i) (right panel). GABA biosensor cells are pre-loaded with the [Ca2+]i indicator Fura-2 and plated on poly-d-lysine coated cover slips in a perfusion chamber. Individual islets are placed on top of this layer of biosensor cells and connected to a closed bath small volume imaging chamber to ensure linear solution flow and fast exchange. b. Titration of exogenous GABA showing concentration-dependence of Ca2+ flux in GABA biosensor cells. The plot shows the average 340/380 Fura-2 ratio of n = 5 GABA biosensor cells in the same field of view. Mean ± SEM. c. Effect of the selective GABAB receptor antagonist CGP5584 on biosensor cell responses to exogenously applied GABA. n = 5 biosensor cells in the field of view. Mean ± SEM. d. Biosensor cell intracellular Ca2+ responses remain elevated during sustained (30 min) exposure to GABA (100 µM shown). n = 5 GABA biosensor cells in the field of view. Mean ± SEM. e. GABA release from a human islet maintained in 3 mM glucose. n = 5 GABA biosensor cells located under or immediately downstream of the islet. This is a representative trace of experiments performed on 40 human islet preparations. Mean ± SEM. f. Biosensor cells have tonic responses to continuously applied GABA and phasic responses to GABA pulses released from islets. Periods of pulsatile GABA release measured from n = 22 human islet preparations, ≥ 3 islets per preparation (black circles) as shown in panel e. Calculated periods for biosensor cell responses to continuously applied GABA (gray circles) at 0.1, 1, 10, and 100 µM GABA as shown in panel d. Center line indicates the mean. g. GABA release from a human islet maintained in 3 mM glucose without addition of inhibitors. n = 5 GABA biosensor cells located under or immediately downstream of the islet. This is a representative trace of experiments performed on 40 human islet preparations. Mean ± SEM. h. Effect of the selective GABAB receptor antagonist CGP5584 on biosensor cell detection of GABA released from a single human islet. n = 5 GABA biosensor cells located under or immediately downstream of the islet. Trace is representative of 3 independent experiments with different human islet preparations. Mean ± SEM.
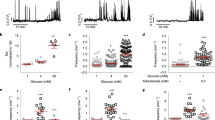
Extended Data Fig. 4 GABA release does not depend on glucose but is activated by VRAC opening.
a-b. Titration of glucose concentrations from 0-25 mM has no effect on islet GABA release. HPLC quantification of GABA released from rat (a) and human (b) islets during 30 mins static incubation in KRBH of the indicated glucose concentrations. n = 4 samples of 100 islets. One-way ANOVA, P = 0.5563 rat (a), P = 0.2053 human (b), ns = not significant. Mean ± SEM. c. HPLC quantification of GABA released from human islets in 5.5 mM glucose (n = 4 samples of 100 human islets), 30 mM KCl (n = 3 samples of 100 human islets), or diazoxide (100 µM) (n = 4 samples of 100 human islets). One-way ANOVA, P = 0.1511, ns = not significant. Mean ± SEM. d. Effect of the GAT inhibitors SNAP5114 (50 µM), NNC05-2090 (50 µM), and NNC711 (10 µM) on biosensor detection of GABA secretion from a human islet. GAT inhibitors were present throughout the shaded portion of trace. Results are representative of the data plotted in panel e. e. Quantification of GABA release following treatment with GAT inhibitors. Box extends from 25th to 75th percentiles, center line represents the median, whiskers represent smallest to largest values. n = 3 islets, One-way ANOVA, 0-20 min vs. 20-40 min (*P = 0.002), 0-20 min vs. 40-60 min (P = 0.9194), 0-20 min vs. 60-80 min (*P = 0.0058).
Extended Data Fig. 5 Allylglycine inhibition of beta cell GABA content and secretion.
a-b. Validation of GABA antibody via immunostaining of paraformaldehyde-fixed rat hippocampal neurons (a) or rat islet cell monolayers (b) for GAD65, GABA, and insulin (not shown) without or with addition of soluble GABA to the primary antibody incubation buffer; or without or with preincubation of cells with allylglycine (10 mM) to inhibit GABA biosynthesis. Images are representative of 3 experimental replicates. Scale bars 20 µm. c. Immunostaining of paraformaldehyde-fixed rat islets cell monolayers for GABA, insulin, and GAD65, following allylglycine (10 mM) addition and removal. Images are representative of the dataset plotted in panel d. d. Quantification of GABA mean fluorescence intensity (MFI) in rat islet cell monolayers in allylglycine timecourse experiments shown in panel c. n = 4 coverslips. Mean ± SEM. e. HPLC analysis of GABA release from human islets during a 30 min addition of allylglycine (no pre-incubation). n = 3 samples of ~100 islets each. Statistical analysis by two-tailed t-test, *P = 0.0104. Mean ± SEM.
Extended Data Fig. 6 Human islet single-cell RNA-seq for expression of genes of interest.
a. Expression of neurotransmitter transporter family genes (SLC6A). Mean ± SEM. b. Expression of genes of interest reported in the literature as related to GABA or putative GABA membrane transporters. Mean ± SEM. Data shown are from two datasets54,55, but results agree with and are representative of three different curated human single-cell RNA-seq datasets analyzed54,55,56 (see also Figure 4).
Extended Data Fig. 7 Kymographs of individual GABA biosensor cells.
a-b. Still image, kymographs, and average trace from timelapse videos of Fura-2 [Ca2+]i signals in GABA biosensor cells in a perfusion flow field in 3 mM glucose isotonic KRBH exposed to (a) 0.1, 1, and 10 μM GABA, (b) downstream from a wild type mouse islet, and (c) downstream from a βc-LRRC8A-/- mouse islet. GABA (1 μM) is added to (c) at 23 min. GABA-responsive cells were selected for analysis, while unresponsive cells were not analyzed. Data are representative of three independent experiments. See also Supplementary Videos 1–3.
Supplementary information
Supplementary Table 1
Data summary comparing the expression of GAD65, VGAT and GABA in human and rat islet endocrine cell subtypes.
Supplementary Tables 2 and 3
Average relative mRNA expression of all SLC genes in human islets from three different single-cell RNA-seq datasets. Buffer composition and osmolarity calculations.
Supplementary Video 1
GABA biosensor cell calibration. Timelapse video of Fura-2 [Ca2+]i signal in GABA biosensor cells exposed to 0.1, 1 and 10 μM GABA. Data are representative of five independent experiments. See also Extended Data Fig. 7.
Supplementary Video 2
GABA biosensor cell responses to a wild-type mouse islet. Timelapse video of Fura-2 [Ca2+]i signal in GABA biosensor cells in proximity to a wild-type mouse islet. Data are representative of four independent experiments. See also Extended Data Fig. 7.
Supplementary Video 3
GABA biosensor cell responses to a βc-LRRC8A−/− mouse islet. Timelapse video of Fura-2 [Ca2+]i signal in GABA biosensor cells in proximity to a βc-LRRC8A−/− mouse islet. GABA (1 μM) is added at 23 min. Data are representative of three independent experiments. See also Extended Data Fig. 7.
Rights and permissions
About this article

Cite this article
Menegaz, D., Hagan, D.W., Almaça, J. et al. Mechanism and effects of pulsatile GABA secretion from cytosolic pools in the human beta cell. Nat Metab 1, 1110–1126 (2019). https://doi.org/10.1038/s42255-019-0135-7
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/s42255-019-0135-7
This article is cited by
-
Cell volume controlled by LRRC8A-formed volume-regulated anion channels fine-tunes T cell activation and function
Nature Communications (2023)
-
Structural basis for assembly and lipid-mediated gating of LRRC8A:C volume-regulated anion channels
Nature Structural & Molecular Biology (2023)
-
Monitoring hormone and small molecule secretion dynamics from islets-on-chip
Analytical and Bioanalytical Chemistry (2023)
-
The physiological role of β-cell heterogeneity in pancreatic islet function
Nature Reviews Endocrinology (2022)
-
Phenome-wide analysis of Taiwan Biobank reveals novel glycemia-related loci and genetic risks for diabetes
Communications Biology (2022)
