Thank you for visiting nature.com. You are using a browser version with limited support for CSS. To obtain
the best experience, we recommend you use a more up to date browser (or turn off compatibility mode in
Internet Explorer). In the meantime, to ensure continued support, we are displaying the site without styles
and JavaScript.
The taxonomically diverse terrestrial tetrapod fauna from the late Mississippian East Kirkton Limestone includes the earliest known members of stem Amphibia and stem Amniota. Here we name and describe a new East Kirkton tetrapod with an unusual hindlimb morphology reminiscent of that of several stem- and primitive crown amniotes. It displays a unique ilium with two slender and elongate processes and a 5-digit pes with a long, stout metatarsal IV and a greatly elongate digit IV. The new taxon broadens our knowledge of East Kirkton tetrapods, adding to the remarkable diversity of their hindlimb constructions, functional specializations, locomotory modes, and adaptations to a wide variety of substrates. An unweighted character parsimony analysis places the new taxon in a polytomy alongside some other Carboniferous groups. Conversely, weighted parsimony and Bayesian analyses retrieve it among the earliest diverging stem amniotes, either as the basalmost anthracosaur or within a clade that includes also Eldeceeon and Silvanerpeton, crownward of an array of chroniosaurs plus anthracosaurs.
The Carboniferous tetrapod fauna from the East Kirkton Limestone of the Bathgate Hills in Scotland provides a unique window onto the diversity of terrestrial vertebrates during the late Mississippian (early Carboniferous), ~336 Ma (million years ago). The seven East Kirkton tetrapods described and named so far are both taxonomically diverse and morphologically disparate, exhibiting a wide range of body shapes and sizes, vertebral constructions, and limb proportions. This level of diversity among tetrapods is not encountered again until the mid-Pennsylvanian, some 25 million years (Myr) later, by which time several clades, particularly among crown Amniota, are well established and greatly diversified1. Among the tetrapods represented at East Kirkton are the temnospondyl Balanerpeton2, the anthracosaur-like Silvanerpeton3,4 and Eldeceeon5,6, and the amniote-like Westlothiana7. In some recent studies8,9,10, these taxa have been placed phylogenetically as the earliest known members of stem Amphibia (Balanerpeton) or stem Amniota (Westlothiana; Silvanerpeton; Eldeceeon; but see ref. 11 for alternative hypotheses on the affinities of many of these taxa, e.g., as stem tetrapods), and thus provide a minimum age estimate for the origin of crown Tetrapoda8,12. Other East Kirkton taxa include the baphetid-like Eucritta12,13, the aïstopod Ophiderpeton14, and the putative microsaur Kirktonecta15. Except for Ophiderpeton, all these taxa are among the earliest diverging members of clades that are more commonly found throughout the later Palaeozoic and combine several of the most plesiomorphic traits of those clades.
In this article, we name and describe a new tetrapod from East Kirkton, based upon postcranial remains that show unusual specializations to the pelvic and hindlimb skeletons, until now observed only in many later tetrapods, as well as a suite of unique characteristics. The holotype was collected by the late Stan Wood and donated to the University Museum of Zoology, Cambridge (UMZC), probably in the 1990s, although details of its collection were, unusually, not recorded by him. Possible reasons for this are that the specimen derived either from one of the spoil heaps from the old quarry workings of the locality or from one of the boundary walls that Stan had bought and collected from before the main East Kirkton quarry was discovered16. The matrix lithology suggests that the specimen may have originated from Black Shale Unit 8217, a particularly fossil-rich horizon that has yielded several compressed, but often near-complete and slightly disarticulated tetrapods. The unusual circumstances of fossil preservation, in a locality affected by volcanic activity and including a mineral-rich lake fed by warm or hot springs, have been described in a series of papers in Rolfe et al.18 and summarized in Clack19. As well as the earliest known terrestrial tetrapods, the East Kirkton quarry has also yielded fish, arthropods, and plants18.
The new taxon prompts a reconsideration of the importance of the East Kirkton site for our understanding of the evolutionary and ecological contexts in which tetrapods radiated and diversified during the Viséan stage of the early Carboniferous. More broadly, its significance lies in the fact that it casts new light on the emergence of tetrapod terrestrial adaptations, such as are revealed by its unusual pelvic and hindlimb skeletons.
Termonerpeton makrydactylus gen. et sp. nov. (Fig. 1)
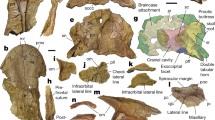
Fig. 1: Termonerpeton makrydactylus gen. et sp. nov. holotype UMZC 2019.1.
a Specimen photograph. b Interpretive drawing. Scale bars 10 mm. Abbreviations: acet acetabulum, fem femur, fib fibula, ha haemal arch, ic intercentrum, l left, na neural arch, phal phalanx, piliac p post-iliac process, plc pleurocentrum, r right, sac rib sacral rib, tib tibia.
Genus: from τέρμωυ (térmon) meaning boundary and ερπετό (erpetó) meaning ‘crawler’, referring to the field boundary walls near the East Kirkton quarry where the late Stan Wood initially discovered fossils from the East Kirkton Limestone and from where the type specimen may have been collected; species: from μακρύς (makrýs) meaning ‘elongate’ and δάχτυλο (dáchtylo; more precisely, δάχτυλο ποδιού, dáchtylo podioú) meaning ‘toe’, referring to the very long pedal digit IV.
Holotype
University of Cambridge Museum of Zoology (UMZC) 2019.1. A partial tetrapod postcranium, preserving both pelves, a femur, fibula, tibia, and an almost complete but disarticulated pes. Closely associated with the appendicular elements are dorsally open hoop-shaped centra, a few neural and haemal arches, curved ribs, and a section of articulated gastralia.
Locality and horizon
East Kirkton quarry, near Bathgate, Scotland, UK. East Kirkton Limestone, Bathgate Hills Volcanic Formation. Exact horizon is unknown. Brigantian, Viséan, early Carboniferous (=Mississippian)21.
Differential diagnosis
Possible autapomorphies: ilium with drawn-out, flat, blade-like dorsal process; very large, stout, and elongate metatarsal IV, greatly exceeding the length of metatarsals III and V (~30% or more). Possible tetrapod synapomorphies among post-Devonian taxa: distinct interepipodial space between tibia and fibula; well-ossified tarsus comprising tibiale, fibulare, intermedium, four centralia, and five distal tarsals. Possible amniote synapomorphies, but often showing reversed polarity in several stem- and crown amniote taxa: presumed pedal phalangeal formula 23454; robust and long pedal digit IV; enlarged intermedium and fibulare, together occupying more than half of proximal moiety of tarsus; curved ribs. Characters of uncertain polarity (also present in Caerorhachis): elongate, slender, and posterodorsally oblique post-iliac process; short puboischiadic plate with almost vertical anterior margin; stout femur with poorly pronounced waisting along the shaft, longer than puboischiadic plate; hoop-shaped centra.
Attributed specimen
National Museums Scotland (NMS) G.1992.22.1. An articulated, partially complete, large tetrapod pes, preserving a nearly complete array of tarsals, all metatarsals, and the proximal phalanges of digits I–III. Unit 82, East Kirkton Limestone, East Kirkton quarry, near Bathgate, Scotland, UK.
Specimen description
Appendicular skeleton
Most of the description is based upon the holotype. Both pelves are preserved, one mainly as a natural mould. The puboichiadic plates are short and deep, with an almost vertical anterior margin to the pubis (Fig. 1). In one, the surface of the puboischiadic plate is strongly convex, in the other it is strongly concave. The concave plate may belong to the left pelvis, with the concavity indicating the acetabulum. Both iliac processes of the presumed right ilium are overlain by a neural arch and part of the femur and cannot be seen. The presumed left ilium shows a long, posteriorly pointing post-iliac process that extends as far backward as the posterior edge of the ischium. It retains the proximal, stump-like portion of a dorsal iliac process, continued distally in natural mould as a mediolaterally flattened and blade-like structure. Both processes sit above a short iliac neck. The dorsal iliac process is proportionally longer than in other tetrapods and its knife-like appearance is unique. The angle between the two processes is much more acute than in most other tetrapods, and the nearest comparison is with the divided iliac process of the microsaurs Tuditanus and Ricnodon22 which, however, could merely represent a bifid post-iliac process. Two gaps in ossification are taken as evidence of an ilio-ischiadic suture half-way down the posterior margin on the left pelvis and an ilio-pubic suture halfway down the anterior margin of the right pelvis (Fig. 1). There is no evidence of a puboischiadic suture, although a shallow depression along the ventral margin of the left puboischiadic plate probably marks the junction between pubis and ischium. The complete left puboischiadic plate is 20 mm deep behind the ilium and 30 mm long, with the pubis contributing about one-third of its length and the ischium the remaining two thirds. The anterior margin of the pubis is almost vertical. The dorsal margin of the ischium is shallowly convex for half its length before extending posteroventrally to meet the upturned posterior extremity of the ischium’s ventral margin. There is no evidence as to the angle at which the two pelvic plates met at the symphysis, which would affect the position of the acetabulum relative to the substrate, and thus the effective resting posture of the hindlimb.
The left femur is at least 39 mm in length, and longer than the puboischiadic plate. The entire bone is crushed, and its distal end lies partly beneath one of the pelvic halves and a neural arch so that its features cannot easily be made out. A possible intercondylar groove may be present distally, and the extensor surface of its proximal extremity appears to show a subcentral depression. The femur itself is robust with little waisting at mid-shaft. A small internal trochanter lies near its proximal end. The left fibula is approximately 26 mm long along its lateral margin. Its proximal end is narrow and grooved. Its broad and strongly flared distal end suggests a broad contact with the tarsus. The medial turn of the distal end indicates a large interepipodial space. The left tibia is about 20 mm long, slender, and shallowly waisted at mid-shaft. It is not clear which end is proximal and which distal, although probably the proximal is the broader. The tibia is probably more than half the length of the femur. Based upon the femur and tibia lengths, and omitting the ankle and pes, the above figures indicate a total stylopod-zeugopod length of about 65 mm, assuming a fully extended limb.
Most of the morphology of the left pes is preserved, showing many well-ossified tarsal bones (Fig. 2). Several of these, including possible distal tarsals II and III lie more or less in anatomical continuity relative to metatarsals II and III, respectively. Other tarsal elements, including possible fibulare, tibiale, centralia, and distal tarsals, are illustrated in Fig. 2. Metatarsal IV lies in anatomical position relative to metatarsals II and III and, at 7 mm in length, it is significantly larger than the latter. The presumed first phalanx of pedal digit IV lies close to metatarsal IV, at an angle of nearly 90° to the latter. It is long and slender, indicating an unusually elongate fourth pedal digit.
Fig. 2: Termonerpeton makrydactylus gen. et sp. nov. left hindlimb of UMZC 2019.1.
a Specimen photograph, showing close-up view of hindlimb skeleton, b Interpretive drawing, with centralia, distal tarsals, and metatarsals indicated by red, blue, and black Roman numerals, respectively, c Interpretive drawing with dashed lines connecting elements of individual digits, d Reconstruction of left tibia, fibula and pes. Scale bars 10 mm. Abbreviations: interm intermedium, tib tibialia.
An array of about 12 phalanges is preserved. They are all disrupted but occur in proximity to one another and, like the first phalanx of pedal digit IV, also mainly lie at right angles to metatarsals III and IV. An additional, acutely angled pointed ungual phalanx, possibly associated with digit II, is also visible. A further two phalanges have been displaced and rest along the anterior edge of the left pelvis. In total, we were, therefore, able to identify 15 elements. The preservation of the pes suggests it was strongly flexed either at death or from tissue shrinkage thereafter. An isolated metatarsal, presumably from the other, missing foot, lies some distance away near the edge of the block. Together, the pedal elements suggest a relatively large foot.
A second specimen, NMS G.1992.22.1 (Fig. 3), is represented by an isolated pes. It may belong to Termonerpeton, although it is from a much larger individual. It shows five metatarsals of which the fourth is much longer and more robust than the other four and about twice as long as that of the holotype, while metatarsal V is the smallest. There are three phalanges, plus five distal tarsals. A D-shaped element closely associated with three centralia could be either a fibulare, a displaced intermedium, or centrale IV.
Fig. 3: Termonerpeton makrydactylus gen. et sp. nov partial pes, attributed specimen NMS G.1992.22.1.
a Specimen photograph, b with centralia, distal tarsals, and metatarsals indicated by red, blue, and black Roman numerals, respectively. Scale bars 10 mm.
Where visible, neural arches have short neural spines and prominent zygapophyses, but their shape is hard to assess as none is well preserved. The element overlying part of the right pelvis and the femur is 7 mm high in total. Numerous dorsally open, hoop-shaped centra about 5 mm in diameter are visible, as well as a few small, oval, shallowly curved elements (Fig. 1). Without further evidence, it is uncertain which of these elements are intercentra and which pleurocentra, though we assume that the larger elements are pleurocentra. The preserved ribs are slender and curved, and include trunk ribs, a possible presacral rib, a possible sacral rib, and a possible postsacral rib. This is long but more or less straight. A bone situated among a cluster of centra, somewhat distant from the other tarsal bones, was originally interpreted by us as a possible fibulare, similar to the fibulare in Proterogyrinus23. However, it might also be interpreted as a sacral rib. If so, its morphology is unique. It is short and widens distally into a fan-shaped structure but does not appear to have a bifid proximal end, unlike the sacral rib in Proterogyrinus23. Three haemal arches are present, one still attached to its half-hoop centrum, a second slightly longer, and a third very short and presumably from a more posterior region of the tail.
Comparisons
The exceptional preservation of tetrapods from the East Kirkton Limestone provides a unique opportunity to study portions of the skeletal anatomy that are otherwise poorly preserved or absent among Mississippian tetrapods. In particular, hindlimbs with a complete or near-complete array of tarsal elements and digits are notably rare. The unusual construction of the pes of Termonerpeton prompted us to examine the hindlimb morphology of six other East Kirkton tetrapods (Fig. 4a–g) alongside a selection of additional, mostly Carboniferous taxa (Fig. 4h–n). We focus on epipodials, tarsi, phalangeal formulae and digit length and proportions. To facilitate visual inspection of these elements, all hindlimbs are drawn to a common tibial length, except for the stem diapsid Petrolacosaurus, in which the epipodials are greatly elongate.
Fig. 4: Comparison of the left tibia, fibula, tarsus, and digits of early tetrapods.
aBalanerpeton after 2, bEucritta after 12, cEldeceeon after 6, dSilvanerpeton after 4, eWestlothiana after 7, fKirktonecta original, see 15 (the grey area marks the estimated position and extent of the tarsus), gTermonerpeton, hPederpes after 24, i, Greererpeton after 27, jCaerorhachis after 31, kArcheria after 30, lHylonomus after 28, mTuditanus after 22, nPetrolacosaurus after 29. Drawn to the same tibial length apart from n. Scale bars 10 mm.
In terms of pes size relative to the tibia, the East Kirkton taxa Balanerpeton, Eucritta, and Silvanerpeton (Fig. 4a, b, d) are similarly proportioned. In contrast, Eldeceeon and Westlothiana (Fig. 4c, e) exhibit somewhat larger pedes. Kirktonecta has proportionally the largest pedes of all (Fig. 4f). Termonerpeton (Fig. 4g) has a pes of similar size to the first three taxa except that digit IV is relatively much longer than in any of the others, with an exceptionally large metatarsal IV. In all those taxa in which digit IV is fully preserved, it is the longest, especially in Eldeceeon and Kirktonecta, but in none does it approach in size and proportions that of Termonerpeton. The illustrated limbs also differ from one another in the degree of ossification of the tarsal bones. Most taxa except Eucritta have some indication of ossified tarsal elements, and some of them, including Balanerpeton and Silvanerpeton, show a complete or almost complete tarsal set. Kirktonecta does have an ossified tarsus, but specimen preservation does not allow us to identify individual elements. The phalangeal count, where known, also varies: 22343 in Balanerpeton2; 223?? in Eucritta12; 23455 in Silvanerpeton4; 23454 in Eldeceeon6, Kirktonecta15, Termonerpeton, and Westlothiana7.
In addition, we compared the pedes of East Kirkton tetrapods with those of seven other taxa (Fig. 4h–n): one earlier, Pederpes24; one almost contemporary, Caerorhachis25; four later Carboniferous, Greererpeton26, Hylonomus27, Tuditanus22, and Petrolacosaurus28; and one early Permian, Archeria29. Of these, Greererpeton has relatively the smallest pes. In most, digit IV is the longest, though in Pederpes and Caerorhachis it is incomplete. The pes of Caerorhachis was originally restored with only three phalanges in digit IV30. This is probably incorrect and would be unusual in Carboniferous tetrapods. The pes of the anthracosaur Archeria was originally reconstructed with digit V as the longest29, but again this is unusual among later Carboniferous and early Permian tetrapods and we suspect that digits IV and V have been transposed, and Romer himself expressed doubt about this reconstruction29. In either case, the phalangeal formula of Archeria is similar to that of the East Kirkton anthracosaur Silvanerpeton, as 23455.
Among Carboniferous tetrapods, temnospondyls such as Balanerpeton and colosteids such as Greererpeton show a digit IV that is somewhat longer than the others, but metatarsal IV is very similar in length and breadth to the adjacent metatarsals. In anthracosaurs, digit IV is the longest, but again metatarsal IV is not significantly broader than adjacent metatarsals. This is also the case in the early amniote Hylonomus and the microsaur Tuditanus. Among the taxa illustrated here, Termonerpeton shows a strikingly similar pes to that of the Late Pennsylvanian araeoscelidian diapsid Petrolacosaurus (Fig. 4n). In both, metatarsal IV is significantly longer and stouter than others and forms part of a similarly long digit IV. In early amniotes, an elongate digit IV coupled with an elongate metatarsal IV is a common occurrence in other taxa, such as protothyridids (e.g. Anthracodromeus31), basal araeoscelidians (e.g. Spinoaequalis32), younginids (e.g. Youngina33), saurians33, and basal synapsids (e.g. Heleosaurus34,35,36), among others.
Based upon available evidence, an elongate digit IV is likely to be the plesiomorphic condition for crown amniotes, being present in Hylonomus, Paleothyris, and Petrolacosaurus (Fig. 4l, n), and shortening of this digit certainly represents a derived feature. In later crown amniotes, the conditions vary, with larger, heavier-bodied tetrapods such as dicynodonts and diadectids having generally shorter toes and adopting a more clearly plantigrade posture. An elongate metatarsal IV and associated digit, however, are not universal among Palaeozoic amniotes, and modifications of these conditions occur repeatedly across clades. For instance, in the eureptile captorhinid Eocaptorhinus, digit IV is also the longest, but the length of metatarsal IV does not greatly exceed that of other metatarsals37. The same is true of some early Permian clades, including seymouriamorphs (e.g. Seymouria38; Discosauriscus39), and diadectids (e.g. Diadectes40), although in the diadectomorph Orobates digit III is a little longer than digit IV41. Among synapsids, dicynodonts such as Diictodon42 and caseids43, to name a few, have five pedal digits of approximately uniform length.
We further point out that, while digit IV attains a certain degree of elongation in other early tetrapod groups, such as temnospondyls, in none of them do the relative proportions of this digit (where known) compare to those of several stem and crown amniotes (Fig. 4).
Phylogenetic relationships
The results of various phylogenetic analyses lend some support to the interpretation of Termonerpeton as a stem amniote, despite its uncertain placement in the unweighted character parsimony analysis (Fig. 5a). In the latter analysis, Termonerpeton appears in a polytomous node alongside baphetids (Eucritta; Baphetes; Megalocephalus), temnospondyls (Balanerpeton; Dendrysekos), the anthracosauroids Eldeceeon and Silvanerpeton, and the problematic Caerorhachis. In all other analyses—implied weights, reweighted characters, and Bayesian—Termonerpeton is placed on the amniote stem group, albeit in different positions, among a diverse array of ‘reptiliomorph’ clades and grades. In the implied weights analysis (Fig. 5b), Termonerpeton, Silvanerpeton, and Eldeceeon form a monophyletic group branching crownward of chroniosaurs plus anthracosaurs and anti-crownward of paraphyletic gephyrostegids. In the reweighted analysis (Fig. 5c), Termonerpeton and Caerorhachis appear as successive sister taxa, in that order, to monophyletic anthracosaurs. In the Bayesian analysis (Fig. 5d), the amniote total group receives moderate support with a credibility value (c.v.) of 76 with Caerorhachis as the most plesiomorphic stem amniote. Crownward of Caerorhachis is a polytomy with low support (c.v. = 59) that subtends Termonerpeton, a clade consisting of Eldeceeon plus Silvanerpeton, a clade of anthracosaurs, and a clade that includes all remaining taxa. In crownward succession, these taxa include chroniosaurs, gephyrostegids, seymouriamorphs, Solenodonsaurus, and Westlothiana as successive sister groups to a strongly supported (c.v. = 100) clade containing diadectomorphs, synapsids, and eureptiles. Although eureptile monophyly is not retrieved, strong support (c.v. = 100) is given to the branch subtending diadectomorphs plus synapsids44.
Fig. 5: Results of phylogenetic analyses.
a Strict consensus of 120 shortest trees from unweighted analysis (tree length = 1286 steps, ensemble consistency index C.I. = 0.2738 without uninformative characters, ensemble retention index R.I. = 0.5768), b Single tree from implied weights analysis (tree length = 1298 steps, Goloboff fit = −202.59266, C.I. = 0.2712, R.I. = 0.5713), c Single tree from reweighted analysis (tree length = 212,68965 steps, C.I. = 0.4755, R.I. = 0.774), d Bayesian topology with branches reporting credibility values.
The East Kirkton biota includes a diverse tetrapod fauna with unique morphofunctional and ecological specializations. In particular, the diversity of their limb and digit proportions and phalangeal constructions suggests a variety of locomotory modes. Kirktonecta features aquatic adaptations15, as evidenced by its deep tail and paddle-like hindlimbs. Five taxa (Balanerpeton; Eucritta; Eldeceeon; Silvanerpeton; Westlothiana) appear to have been more fully terrestrial or semi-terrestrial2,3,4,5,6,7,12,13,14, as suggested by the presence of fully ossified and tightly appressed tarsal bones (such as are observed in Balanerpeton, Silvanerpeton and Westlothiana), robust and strongly ossified limbs with well-developed muscle attachment processes, finished margins of dermal pectoral elements, and absence of lateral line canals3. The limbless Ophiderpeton is also considered to be terrestrial or semi-terrestrial, due to the lack of sensory canals14. Except for Ophiderpeton, all other East Kirkton tetrapods exhibit a five-digit pes, a condition first recorded in the Tournaisian24. There is no evidence of polydactyly in any other known Carboniferous tetrapod, unlike in late Devonian taxa (e.g. Acanthostega; Ichthyostega; Tulerpeton)45.
Although Termonerpeton is known from incomplete material, its hindlimb is known in sufficient detail and permits reasonable inferences about its locomotion. Comparisons with modern analogues from among squamate reptiles—a focal group in biomechanical studies of amniote locomotion—are helpful in this respect. In several extant lizards, a large pes with a long metatarsal IV and associated digit occur commonly and are often highly specialized. For example, in runners they act, inter alia, as an energy-saving mechanism generally linked to increased stride length46. Digit IV is the last to leave the ground during a stride cycle and confers extra leverage when the animal moves fast. In addition, the tendons of elongate digits are known to operate as springs47, further assisting energy-saving during running. Ruta et al.6 proposed a similar mechanism in the East Kirkton anthracosauroid Eldeceeon (Fig. 4c). The unusually large pes of this tetrapod was hypothesized to increase stride length and to facilitate forward propulsion at the end of each stride cycle when the pes was fully extended backwards. In Eldeceeon, however, digit IV is comparatively less long and robust—relative to digits III and V–than its homologue in Termonerpeton. If indeed Termonerpeton used its pes in a similar way, then its digit IV is likely to have functioned as the principal, or sole, lever during locomotion.
It is noteworthy that a long digit IV also provides remarkable biomechanical versatility in extant squamates. As an example, it allows the Zebra-tailed lizard to run on compact and granular substrates46. The varied terrain of the East Kirkton environment17,18,19,20,21 suggests that, perhaps, the pes of Termonerpeton afforded this animal some degree of biomechanical versatility. However, it is possible to rule out cursorial habits in Termonerpeton. Its hindlimb lacks the suite of traits usually associated with running capabilities in several extant and fossil tetrapods, such as the basilisk lizard and the Early Permian bolosaurid parareptile Eudibamus, to name two well-known examples. The traits in question include, inter alia, very elongate, subparallel, and slender tibia and fibula delimiting a narrow interepipodial space, rod-like and closely packed metatarsals III-V, and elongate and gracile proximal phalanges of digits III–V. The construction of the Termonerpeton pes, particularly its sturdy and wide tarsus and the extensive sutural contact between its fibulare and the broad distal extremity of its fibula, appears to us to be indicative of a graviportal stance.
Indirect support for this conclusion comes from a consideration of the estimated body size of this animal. If we assume that Termonerpeton scaled isometrically with Eldeceeon and that both taxa had comparable presacral:femur length ratios, then Termonerpeton is estimated to have reached a presacral length of ~75 cm, more than twice the length of the Eldeceeon holotype5 (~35 cm; 18 mm femur length). If we use Balanerpeton as a reference taxon, we obtain a more modest figure. One of the most complete paratypes of Balanerpeton measures 17.5 cm in length, with a femur of 14.23 mm2. Scaling Termonerpeton to this specimen gives an estimated length of ~48 cm. Based upon these figures, we conclude that Termonerpeton was probably much more heavily built than other East Kirkton tetrapods. However, it remains a possibility, that the elongate digit IV enabled Termonerpeton to move more quickly than its heavy build might suggest.
Evolutionary implications
The major finding of our study is that Termonerpeton is the earliest known tetrapod to exhibit a fundamentally amniote-like pes48, typified by elongate metatarsal IV and corresponding digit IV, and by enlarged intermedium and fibulare that occupy more than half of the proximal moiety of the tarsus. More proximal segments of the hindlimb appear less specialized. Thus, the tibia and fibula are relatively stout, and the robust femur has a weakly pronounced mid-shaft waisting. This combination of plesiomorphic and apomorphic traits suggests that the emergence of fully terrestrially adapted limbs among stem amniotes followed a less regular pattern than previously surmised. Both embolomeres and gephyrostegids retain unspecialized tibia and fibula which, discounting minor proportional differences, resemble their homologues in Termonerpeton. In contrast, their femora appear more derived than those of Termonerpeton in that they display a more pronounced waisting of the shaft. Their pedes, where known, demonstrate a gradual acquisition of amniote-like characteristics48. These include enlargement of the proximal part of the tarsus, progressive elongation of digits (especially digits III–V), and the transverse expansion of the laterodistal extremity of the fibula. These features also occur in seymouriamorphs38,39, diadectomorphs (albeit modified towards metatarsal and phalanx shortening)40,41, and several basal crown amniotes48. Some of these features have recently been recognised in fossil footprints from the late Carboniferous and Permian49,50. However, given the range of hindlimb morphologies present at East Kirkton, and the amniote-like pes found in Termonerpeton, we would advise caution when ascribing particular footprints to specific clades.
Methods
Preparation and visualization
The holotype specimen was prepared using an Emax Evolution Grinder and Polisher fitted with a chuck to accommodate a tungsten carbide rod. Photography by JAC and TRS used a Panasonic Lumix DMC-LZ5. Specimen drawings by JAC and TRS were executed using a camera lucida and followed by image processing with Photoshop CC 2017 and 2019.
Nomenclatural acts
This published work and the nomenclatural acts it contains have been registered in ZooBank, the proposed online registration system for the International Code of Zoological Nomenclature (ICZN). The ZooBank LSIDs (Life Science Identifiers) can be resolved and the associated information viewed through any standard web browser by appending the LSID to the prefix ‘http://zoobank.org/’’ The LSID for this publication is: 72B609CB-DCC1-4BDA-9494-BB97BBEB70E4.
Phylogenetic analyses
To evaluate the phylogenetic position of Termonerpeton, we coded this taxon using a slightly expanded version of a recently published taxon-character data matrix44 which was subjected to maximum parsimony and Bayesian inference analyses by MR. Prior to parsimony analyses, the data matrix was inspected for possible occurrences of ‘rogue’ species51 causing loss of resolution among other, more stable species. No ‘rogue’ taxa were detected using the ‘safe_taxonomic_reduction’ function of the package Claddis52 in the R (v. 4.1.1) environment for statistical computing (https://cran.r-project.org). In the case of parsimony analyses, we explored the results from unweighted, posteriorly weighted and standard implied weighted characters53. Posterior weighting employed the maximum value (best fit) of the rescaled consistency index of each character, such as was obtained from the original unweighted analysis, whereas implied weighting used a value of 6 for the constant of concavity K53. All parsimony analyses were performed in PAUP* v. 4.0a build 169 (https://paup.phylosolutions.com)54 using identical settings, as follows: heuristic search method; tree bisection-reconnection branch-swapping algorithm with 10,000 random stepwise taxon addition sequences, holding a single tree in memory at each replicate; 10 consecutive rounds of branch-swapping applied to all trees stored in memory from this initial search, but with the option for saving multiple trees in effect. Node support was evaluated with bootstrap55 and jackknife56, in each case with 10,000 random character resampling replicates using the fast stepwise addition option.
The Bayesian inference analysis was carried out with MrBayes v. 3.2.6 (http://nbisweden.github.io/MrBayes/index.html)57 using the standard data type option for morphological characters, and an equal-rate model of character-state change. The analysis employed four chains with 107 generations, sampling every 1000 generations, discarding 25% of the obtained samples, and storing branch lengths alongside clade credibility values. Convergence was regarded as satisfactory, with the values of the Potential Scale Reduction Factor58 approaching 1.
The holotype of Termonerpeton makrydactylus is accessioned in the University Museum of Zoology Cambridge (UMZC) register number 2019.1. The attributed specimen is accessioned in the National Museums of Scotland (NMS) register number G.1992.22.1.
The data matrix used in this study is available in Supplementary Data 1 and the character list in Supplementary Note 1 with the Supplementary Information file.
References
Mann, A. et al. Reassessment of historic ‘microsaurs’ from Joggins, Nova Scotia, reveals hidden diversity in the earliest amniote ecosystem. Pap. Palaeontol.6, 605–625 (2020).
Milner, A. R. & Sequeira, S. E. K. The temnospondyl amphibians from the Viséan of East Kirkton, West Lothian, Scotland. Trans. R. Soc. Edinb.: Earth Sci.84, 331–361 (1994).
Clack, J. A. Silvanerpeton miripedes, a new anthracosauroid from the Viséan of East Kirkton, West Lothian, Scotland. Trans. R. Soc. Edinb.: Earth Sci.84, 369–376 (1994).
Ruta, M. & Clack, J. A. A review of Silvanerpeton miripedes, a stem amniote from the Lower Carboniferous of East Kirkton, West Lothian, Scotland. Trans. R. Soc. Edinb.: Earth Sci.97, 31–63 (2006).
Smithson, T. R. Eldeceeon rolfei, a new reptiliomorph from the Viséan of East Kirkton, West Lothian, Scotland. Trans. R. Soc. Edinb.: Earth Sci.84, 377–382 (1994).
Ruta, M., Smithson, T. R. & Clack, J. A. A review of the stem amniote Eldeceeon rolfei, from the Viséan of East Kirkton. Earth Environ. Sci. Trans. R. Soc. Edinb.111, 173–192 (2020).
Smithson, T. R. et al. Westlothiana lizziae from the Viséan of East Kirkton, West Lothian, Scotland, and the amniote stem. Trans. R. Soc. Edinb.: Earth Sci.84, 383–412 (1994).
Marjanovic, D. & Laurin, M. Phylogeny of Paleozoic limbed vertebrates reassessed through revision and expansion of the largest published relevant data matrix. PeerJ6, 1–191 (2019).
Clack, J. A. Eucritta melanolimnetes from the Early Carboniferous of Scotland, a stem tetrapod showing a mosaic of characteristics. Trans. R. Soc. Edinb.: Earth Sci.92, 75–95 (2001).
Clack, J. A. A new microsaur from the early Carboniferous (Viséan) of East Kirkton, Scotland, showing soft tissue evidence. Spec. Pap. Palaeontol.86, 1–11 (2011).
Rolfe, W. D. I. et al. The East Kirkton Limestone, Viséan of Westlothian, Scotland: introduction and stratigraphy. Trans. R. Soc. Edinb.: Earth Sci.84, 177–188 (1994).
Holmes, R. B. The Carboniferous amphibian Proterogyrinus scheelei Romer, and the early evolution of tetrapods. Philos. Trans. R. Soc. Lond., Ser. B306, 431–527 (1984).
Clack, J. A. & Finney, S. M. Pederpes finneyae, an articulated tetrapod from the Tournaisian of Western Scotland. J. Syst. Palaeontol.2, 311–346 (2005).
Ruta, M., Milner, A. R. & Coates, M. I. The tetrapod Caerorhachis bairdi Holmes and Carroll from the lower Carboniferous of Scotland. Trans. R. Soc. Edinb.: Earth Sci.92, 229–261 (2002).
Godfrey, S. The postcranial skeletal anatomy of the Carboniferous tetrapod Greererpeton burkemorani. Philos. Trans. R. Soc. Lond., Ser. B323, 75–133 (1989).
Carroll, R. L. Eosuchians and the origins of archosaurs. In Athlon: Essays on Paleontology in Honour of Loris Shano Russell. (ed C. S. Churcher) 58–76 (ToronToronto: Royal Ontario Museum, 1976).
Reisz, R. R. & Modesto, S. P. Heleosaurus scholtzi from the Permian of South Africa: a varanopid synapsid, not a diaspid reptile. J. Vert. Paleontol.27, 734–739 (2007).
Heaton, M. & Reisz, R. R. A skeletal reconstruction of the early Permian captorhinid reptile Eocaptorhinus laticeps Williston. J. Paleontol.54, 136–143 (1986).
Klembara, J. & Bartik, I. The postcranial skeleton of Discosauriscus Kuhn, a seymouriamorph tetrapod from the Lower Permian of the Boskovice Furrow (Czech Republic). Trans. R. Soc. Edinb.: Earth Sci.90, 287–316 (1999).
Berman, D. S. et al. A new diadectid (Diadectomorpha), Orobates pabsti, from the Early Permian of central Germany. Bull. Carnegie Mus. Nat. Hist.35, 1–36 (2004).
Ray, S. & Chinsamy, A. Functional aspects of the postcranial anatomy of the Permian dicynodont Diictodon and their ecological implications. Palaeontology46, 151–187 (2003).
Stovall, J. W., Price, L. I. & Romer, A. S. The postcranial skeleton of the giant Permian pelycosaur Cotylorhynchus romeri. Bull. Mus. Comp. Zool.135, 1–30 (1966).
Klembara, J. et al. Inner ear morphology of diadectomorphs and seymouriamorphs (Tetrapoda) uncovered by high-resolution x-ray microcomputed tomography and the origin of the amniote crown. Palaeontology63, 131–154 (2020).
Clack, J. A. Gaining Ground: The Origin and Evolution of Tetrapods 2nd edn (Indiana University Press, 2012).
Irschick, D. J. & Jayne, B. C. Comparative three-dimensional kinematics of the hindlimb for high-speed bipedal and quadrupedal locomotion of lizards. J. Exp. Biol.202, 1047–1065 (1999).
Li, C., Tonia Hsieh, S. & Goldman, D. I. Multi-functional foot use during running in the zebra-tailed lizard (Callisaurus draconoides). J. Exp. Biol.215, 3293–3308 (2012).
Piñeiro, G., Demarco, P. N. & Meneghel, M. D. The ontogenetic transformation of the mesosaurid tarsus: a contribution to the origin of the primitive amniotic astragalus. PeerJ4, e2036 (2016).
Lucas, S. G., et al. Carboniferous tetrapod footprint biostratigraphy, biochronology and evolutionary events. Geol. Soc. Spec. Publ. https://doi.org/10.1144/SP512-2020-235.(2021)
Buchwitz, M., Jansen, M., Renaudie, J., Marchetti, L. & Voigt, S. Evolutionary change in locomotion close to the origin of amniotes inferred from trackway data in an ancestral state reconstruction approach. Front. Ecol. Evol. https://doi.org/10.3389/fevo.2021.674779 (2021).
Wilkinson, M. Majority-rule consensus trees and their use in bootstrapping. Mol. Biol. Evol.13, 437–444 (1996).
Lloyd, G. T. Estimating morphological diversity and tempo with discrete character-taxon matrices: implementation, challenges, progress and future directions. Biol. J. Linn. Soc.118, 131–151 (2016).
We thank Daniel Delbarre who prepared the UMZC specimen during an undergraduate summer project and Sarah Wallace-Johnson (formerly Finney; Earth Sciences Department, University of Cambridge), for the loan of the Emmax Evolution Grinder and Polisher. We are grateful to Matt Lowe (University Museum of Zoology Cambridge) for access to UMZC 2019.1 and Nick Fraser and Stig Walsh (National Museums Scotland) for the loan of NMS G 1992.22.1. We thank our journal editor, Luke Grinham, two anonymous reviewers and Torsten Scheyer (University of Zurich) for their constructive criticism and helpful remarks. Tim Smithson thanks Jason Head and Rebecca Kilner (University Museum of Zoology Cambridge) for access to research facilities. This project was supported by a John Templeton Foundation Grant (No 61408) awarded to M.R.
Author information
Author notes
Deceased: Jennifer A. Clack.
Authors and Affiliations
University Museum of Zoology Cambridge, Downing Street, Cambridge, CB2 3EJ, UK
Jennifer A. Clack & Timothy R. Smithson
School of Life and Environmental Sciences, University of Lincoln, Joseph Banks Laboratories, Green Lane, Lincoln, LN6 7DL, UK
J.A.C. conceived the study. J.A.C. and T.R.S. photographed the specimens and prepared the figures. M.R. undertook the phylogenetic analyses. All authors described and analysed the fossil material, interpreted the data, and wrote the manuscript.
Communications Biology thanks Torsten Scheyer, Min Zhu and the other, anonymous, reviewers for their contribution to the peer review of this work. Primary handling editor: Luke R. Grinham. Peer reviewer reports are available.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
Clack, J.A., Smithson, T.R. & Ruta, M. A Mississippian (early Carboniferous) tetrapod showing early diversification of the hindlimbs.
Commun Biol5, 283 (2022). https://doi.org/10.1038/s42003-022-03199-x
Anyone you share the following link with will be able to read this content:
Sorry, a shareable link is not currently available for this article.
Provided by the Springer Nature SharedIt content-sharing initiative
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.