
Abstract
The recent proliferation of pelagic Sargassum spp. in the Tropical Atlantic causes major ecological and socioeconomic impacts to the wider Caribbean when it washes ashore, with regional fisheries and tourism industries particularly affected. The Caribbean influxes have been tracked to a new bloom region known as the North Equatorial Recirculation Region (NERR) encompassing the area between the South Equatorial Current and the North Equatorial Counter Current and extending from Africa to South America. The vast biomass of Sargassum presents serious problems when it washes ashore but also represents significant commercial opportunities, especially with biofuel and fertilizer. The floating Sargassum mats are themselves diverse ecosystems that vary both in their biodiversity and biochemical attributes. Two major species (Sargassum fluitans and S. natans) have been identified as well as several distinguishable morphotypes of each. Oceanic mixing tends to blend the morphotypes together making it difficult to determine if there are regions of the NERR that favour bloom and growth of the distinct types. In this study, we quantify the species and morphotype composition of Sargassum strandings in Barbados and test if this is related to separate oceanic origins and routes travelled using a backtracking algorithm based on ocean drifter data. We found significant seasonal variation in the relative abundance of three morphotypes and this could be traced to two distinct easterly sub-origins and/or transport pathways; one area around 15° N that travels directly E–W across the Atlantic, and another area generally south of 10° N that takes a more meandering route coming close the coast of South America. These findings contribute towards our understanding of why the Tropical Atlantic bloom is presently occurring as well as towards addressing valorisation constraints surrounding variation in the supply of the three commonly occurring morphotypes.
Similar content being viewed by others

Introduction
Pelagic Sargassum spp. (Sargassum natans and S. fluitans) subsequently referred to simply as ‘Sargassum’ forms a floating oceanic ecosystem that provides habitat, shelter and foraging opportunities for a wide diversity of endemic and associated species1,2,3. Historically, this structural habitat was largely confined to the Sargasso Sea and the Gulf of Mexico4,5 with very low abundance in the northern Caribbean and Tropical Atlantic6,7. This dramatically changed in 2011, when massive quantities of Sargassum began to strand and decompose along the coasts throughout the Caribbean, NE South America and West Africa8,9,10 seriously impacting regionally important industries. This proliferation of Sargassum in the Tropical Atlantic has continued and increased, and has been linked to a bloom region bounded latitudinally by the South Equatorial Current (SEC) and the North Equatorial Counter Current (NECC) lying between Africa and South America and defined, for simplicity, as the North Equatorial Recirculation Region (NERR)9,10. Ocean eutrophication and climate change are likely driving factors11,12,13 although little is known about the effects of ocean circulation patterns on the taxonomic composition of Sargassum blooms. In this study, the distribution of Sargassum morphotypes within the Tropical Atlantic is determined as a first step in unravelling this complex problem.
The negative impacts of decomposing Sargassum have been well documented in nearshore ecosystems8,14, fisheries15,16,17, tourism and other coastal businesses8,18,19 and human health20,21,22. Clean-up efforts have cost hundreds of millions of dollars to national economies8,23. Influx events are now considered a new norm to which countries must adapt12,24. There is a rapidly growing interest in utilizing Sargassum and turning it into business opportunities25,26,27,28. However, what constitutes sustainable harvesting and the human and/or environmental health risks of utilizing Sargassum are currently not well understood29.
Sargassum mats originating in the Tropical Atlantic are widely recognised as composing of two species (three dominant morphotypes); Sargassum natans I, S. natans VIII and S. fluitans III (Fig. 1). Whilst there remains some controversy over the taxonomy and nomenclature30,31,32,33,34, these three morphotypes are genetically distinct30 and have distinctive ecological, biological, and chemical traits3,35 and even accumulate toxic heavy metals at different rates36,37.

General appearance of the three pelagic Sargassum morphotypes collected on Morgan Lewis beach, Barbados. Inset on S. natans I shows the presence of spines on the circular air bladders. Inset on S. natans VIII shows the absence of spines on the circular air bladders. The top right inset on S. fluitans III shows the absence of spines on the elongated air bladders and the bottom left inset shows the presence of spines along the stem. Photograph taken from Ref.38.
There is evidence of substantial interannual and/or intra-annual variation in the morphotype composition of Sargassum mats originating from the NERR29,39,40,41. Initial influxes were reported to be dominated by S. fluitans III and S. natans VIII, whilst more recent observations report a dominance of S. fluitans III and S. natans I29,40. To date, however, the variability in relative abundance of Sargassum morphotypes remains poorly understood29. Furthermore, given that morphotype composition can influence both the biodiversity associated with Sargassum3 and chemical composition35,37, variations in the relative abundances of the three commonly occurring morphotypes will have implications for sustainable harvesting strategies and for the utilization of Sargassum by entrepreneurs.
Recent findings42 revealed two potential pathways for Sargassum transport into the Caribbean. We hypothesised that the morphotype composition of Sargassum influx events varies seasonally and that this is related to its oceanic origin and transport pathway across the Atlantic. We assessed variation in relative abundance of the three morphotypes in Sargassum strandings in Barbados during 2021–2022 and analysed their origins with the use of a backtracking algorithm based on ocean drifter data with addition of 0.5% wind43. Barbados is uniquely positioned as a study site, being near the 15° N latitude ‘separation’ line between North Atlantic gyre water and tropical water entering the Caribbean44 and the most easterly of the Caribbean islands, thus among the first to receive Sargassum influxes from the NERR. Therefore, it serves as an ideal site to test our hypothesis regarding seasonal variation and oceanic origins of Sargassum influxes.
Results
Changes in morphotype compositions
Monthly analyses of the relative abundance of S. natans I, S. natans VIII and S. fluitans III revealed temporal differences in the predominant morphotype (Fig. 2, Fig. S1).

Ternary plot illustrating monthly variation in morphotype composition of Sargassum stranding in Barbados over a 12-month period. Sargassum natans I (SNI), S. natans VIII (SNVIII) and S. fluitans III (SFIII). Black lines indicate the average percent contribution of each morphotype in the January samples (67% SNVIII, 25% SFIII, 8% SNI). Quarterly periods are represented by orange (Jan–Mar), yellow (April–Jun), green (Jul–Sept) and blue (Oct–Dec) dots.
While S. fluitans III was the predominant morphotype in 80% of the samples, there were notable exceptions (Fig. 2). For example, in November, January, and February samples were dominated by S. natans VIII, which was generally the least abundant of the three morphotypes.
Linking morphotype compositions to sub-origins
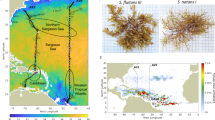
The extent to which the observed variation in the relative abundance of Sargassum morphotypes reflects distinct origins was investigated via sample backtracking for 365 days from the date of stranding using 100 particles to represent each sample. This revealed two distinct sub-origins/transport pathways (Fig. 3). Sargassum stranding in Barbados between March and early August apparently originated close to the equator and travelled along northeast Brazil before arriving in Barbados (Fig. 3a). In contrast, Sargassum arriving between late August and February originated further north and travelled a relatively direct route to Barbados (Fig. 3b).

Backtracks of Sargassum (mixed-morphotype samples) collected from Morgan Lewis beach in Barbados over the period of 1 year (2021–2022). Panel (a) shows sample backtracks from early March to early August and panel (b) shows backtracks from late August through February. All backtracks were run over a 365-day period using 100 particles. White dots indicate the origin of each backtrack particle (i.e., the backtrack location 365 days before arriving to Barbados) and the black square represents the origin of highest probability (mean of all backtrack particles). Dates are presented in year/month/day format. Maps were generated using IDL 8.8.1 (https://www.l3harrisgeospatial.com/Software-Technology/IDL).
The two identified transport pathways differed in the average distance travelled by the particles and average latitude at the origin of each particle backtrack (Fig. 4a). Simulations were attributed to sub-origin/transport pathway A or B based on the route travelled, distance travelled and the approximate location of the origins. Backtracks that originated around the equator (0–7° N; Fig. 4b) and followed a convoluted trajectory along the NE coast of Brazil to the Caribbean (Fig. 3a) are referred to as sub-origin/transport pathway A. These backtracks showed both high average distances travelled (5740 to 8550 km over the 365 days) as well as high variability in the distance travelled among replicate backtracks for any given date (indicated by the relatively large 95% confidence intervals in Fig. 4a). In contrast, backtracks referred to as sub-origin/transport pathway B originated relatively far north (9–18° N; Fig. 4b) and travelled a more direct westerly route covering a much shorter distance over the 365 days (4080–4870 km) than Sargassum from sub-origin/transport pathway A (Fig. 3a,b) and showed greater consistency in the distance travelled among the replicate backtrack particles for any given date (Fig. 4a). This partitioning of the 25 simulations into two “homogenous” groups of sub-origins was supported with the use of K-means partitioning (Fig. S2).

Sargassum sub-origins and transport pathway metrics within the Tropical Atlantic as determined from backtracking monthly Sargassum samples stranded in Barbados. Part (a) shows the average (± 95% CI) distance travelled per track and the average (± 95% CI) latitude of each origin for a given date. Blue dots indicate samples from sub-origin A and orange dots indicate samples from sub-origin B. Part (b) shows the two distinct sub-origins identified. Black square represents the average latitude at the origin of each track for a given date. Map was generated using IDL 8.8.1 (https://www.l3harrisgeospatial.com/Software-Technology/IDL). All backtracks started from the Morgan Lewis beach, located on the east coast of Barbados and were run over a 365-day period using 100 particles. All averages represent mean value of the 100 particles.
A Permutational Multivariate Analysis of Variance (PERMANOVA) indicated that the relative abundance of S. natans I, S. natans VIII and S. fluitans III morphotypes differed significantly between the two sub-origins/transport pathways (p = 0.0131) (Table S1). Sub-origin/transport pathway A (March-early August) showed a predominance of S. fluitans III (Fig. 5a). In contrast, sub-origin/transport pathway B (late August–February) showed higher levels of S. natans VIII, with reduced quantities of S. fluitans III (Fig. 5b).

Ternary plots showing morphotype composition of Sargassum samples stranding in Barbados over a 12-month period. Part (a) shows Sargassum samples from sub-origin/transport pathway A (March–early August). Part (b) shows Sargassum samples from sub-origin/transport pathway B (late August–February). Sargassum natans I (SNI), S. natans VIII (SNVIII) and S. fluitans III (SFIII). Samples are represented by yellow dots and the mean value is represented by a green square.
Oceanographic parameters and sub-origins
We investigated whether environmental conditions, i.e., Sea Surface Temperature (SST) and nutrient load (inferred from Chlorophyll a concentration) derived from satellite imagery, differed between the two sub-origins at the initial locations (‘origins’ (Fig. 4b)) and mid-way (6 months) along the average backtrack trajectory of Sargassum (for a given date). A PERMANOVA indicated that environmental conditions differed significantly between the two sub-origin/transport pathways (p = 0.0039) but not between time periods (origin vs mid-way) (p = 0.3179) (Table S1). Pooling the data across both time periods for each sub-origin supported that environmental differences between the two sub-origins/transport pathways were driven by higher SST at the sub-origin/pathway A (Fig. 6).

Boxplots of sea surface temperature (a) and chlorophyll a concentration (b) for sub-origin/transport pathway A and B (n = 48 in all groups).
Discussion
Since the onset of Sargassum influx events in 2011, there has been notable annual variability in the relative abundance of S. natans I, S. natans VIII, and S. fluitans III, with broadscale spatial and temporal differences observed across the Caribbean29,39,40,41; however, to date there has been little understanding of what drives this. This study supports for the first time that the morphotype composition of Sargassum influxes over the course of a full year varies significantly and appears to be linked to their origin/transport pathway. By using a backtracking algorithm based on ocean drifter data with 0.5% wind, Sargassum from Barbados was traced to two distinct sub-origins/transport pathways within the Tropical Atlantic. Identified pathways align with the findings of Ref.42. In our assessment, Sargassum mats arriving in Barbados between March to early August are likely to take the more southerly transport pathway, linked to the Gulf of Guinea, (arriving from sub-origin A) that passes along the coast of South America. Satellite imagery in Refs.7,45 suggest that Sargassum enters this pathway from south of the equator, which is south of the NERR. Alternatively, Sargassum arriving between late-August and February are likely to take the more northerly transport pathway (arriving from sub-origin B). This matches the seasonal distribution of Sargassum in the NERR as detected by satellites13. Sargassum arriving from sub-origin/transport pathway A were S. fluitans III-dominated while those arriving from sub-origin/transport pathway B had significantly higher amounts of S. natans VIII.
Interestingly, our monthly observations of relative abundance of the three Sargassum morphotypes aligns with morphotype compositions reported in the literature39,40,41. Starting with the November 2014 to May 2015 period41, S. natans VIII was the predominant form observed across the Caribbean. This was especially interesting since no earlier studies had ever reported S. natans VIII-dominated mats throughout the region. Subsequent studies39 in the Mexican Caribbean, also reported large volumes of S. natans VIII during the peak arrival of Sargassum in August 2015. In contrast, recent investigations into the morphotype composition arriving in Jamaica during July and August40 showed a clear dominance of S. fluitans III. Based on the findings of our study, a possible explanation for the large-scale variations observed by Refs.39,40,41 can be linked to the identified sub-origins/transport pathways. It is plausible that 2015 reports of Sargassum39,41 recorded high amounts of S. natans VIII because the majority of their data collection took place during the late August-February (sub-origin/transport pathway B) period, which was associated with significantly higher abundances of S. natans VIII in our analyses. On the other hand, the 2020 summer report40 of Sargassum influx events reported dominance of S. fluitans III; according to the findings of the current study Sargassum mats arriving during this time would have likely arrived from sub-origin/transport pathway A, which shows a significantly higher abundance of S. fluitans III.
While the presence of a southern sub-origin/transport pathway A and a northern sub-origin/transport pathway B provides a plausible explanation for the observed spatial and temporal variations in the relative abundances of S. natans I, S. natans VIII and S. fluitans III, the question remains: why are these sub-origins/transport pathways favouring different morphotypes? In Florida, the two Sargassum species are reported to have different maximum growth rates under optimal conditions, with S. fluitans apparently capable of growing considerably faster than S. natans46. The same study also indicated a difference in thermal tolerances between the two species, stating that S. natans has a broader optimal temperature range (18–30 °C) than S. fluitans. Furthermore, recent studies have also indicated similar differences in growth rates between species, as well as differences among morphotypes from the Tropical Atlantic47. Studies by Lapointe and colleagues comparing growth rates have also indicated that both pelagic species grow significantly faster with nutrient enrichment5,48,49. Therefore, given the two distinct sub-origins/transport pathways, oceanographic conditions between the two areas may have been the proximal cause for the observed morphotype compositions. Our analysis suggests that differing SST within sub-origins/transport pathways influenced the morphotype composition of Sargassum arriving in Barbados. S. fluitans III-dominated mats arriving from the southerly transport pathway A, close to the equator, experienced higher SST when compared to the northerly transport pathway B. Recent studies12 indicate that Sargassum blooms are enhanced by nitrogen-rich neritic waters and that nutrient variability is a key driver of Sargassum variability13. In this study we used chlorophyll a concentration as a direct proxy for nutrient concentration. However, we found high variability in this proxy within each of the two sub-origins/transport pathways, which precluded a clear understanding of its possible role on morphotype composition. Moreover, it is possible that chlorophyll concentration per se might not be a good indicator of nutrient availability for Sargassum. The Equatorial Atlantic has a heterogenous surface environment with contributing nutrient-rich water masses from the Amazon River, the Congo River and equatorial and coastal upwelling50,51. Thus, mats originating close to the equator (sub-origin/transport pathway A) are likely to experience nutrient-rich environments in addition to higher SST than Sargassum from the more northerly sub-origin/transport pathway B, allowing Sargassum to flourish and perhaps amplifying physiological differences among morphotypes that result in differential growth and mortality and thus in different morphotype composition. However, discussions surrounding optimal growth conditions for the three morphotypes remain largely speculative and the extent to which the rate of nutrient uptake varies among morphotypes is not well understood. Interestingly, S. natans VIII and S. fluitans III collected from around 10° N showed different levels (albeit not significant) of %N enrichment; with S. natans VIII having higher %N on average than the other two morphotypes12. If S. natans VIII is indeed capable of absorbing nutrients faster than its counterparts above 10° N, valorisation of this morphotype may be affected. Low phosphate environments increase the uptake of arsenic37, therefore, mats traversing low phosphate environments at and above 10° N may result in S. natans VIII absorbing higher levels of arsenic compared to S. natans I and S. fluitans III, ultimately reducing the applications for which S. natans VIII can be used. Intriguingly, biomass composition of Sargassum arriving in Jamaica during February 2019 had lower quantities of metals in S. natans VIII than S. natans I and S. fluitans III; but when looking at arsenic specifically, S. natans I and S. natans VIII on average contained higher levels of arsenic when compared to S. fluitans III35. Studies on growth and mortality of the three morphotypes in various conditions are required to understand their optimal conditions, the effects of source and dispersal routes on morphotype composition, biomass composition and the potential consequences for valorisation.
The observed seasonality in S. fluitans III and S. natans VIII may also influence the biodiversity associated with Sargassum mats at different times of the year. The greater structural complexity of S. fluitans III supports more organisms when compared to the less foliated S. natans VIII3. This means that S. fluitans III-dominated mats arriving from sub-origin/transport pathway A during March to early August may have greater biodiversity than mats arriving from sub-origin/transport pathway B (late August to February). With more innovative strides being made towards Sargassum valorisation26, in-water harvesting may be the solution to providing large quantities of fresh Sargassum for a variety of uses. However, in-water harvesting may pose a threat to associated biodiversity especially during months where S. fluitans III is the dominant morphotype. To better understand the implication(s) of changing morphotype composition on Sargassum associated biodiversity and valorisation efforts, further research into the seasonality of Sargassum morphotypes is required. Studies should ideally be conducted across the Caribbean to provide a comprehensive understanding of the situation and aid in the region’s continued adaptation to Sargassum influx events.
There are limitations to our study. As such generalizations within or across years or countries should not be made with only one year of data. We cannot say if the results of 2021–2022 reflect a typical year or if the findings were unique to Barbados given that it is one of the first islands to receive Sargassum from the NERR. As Sargassum travels from eastern to western countries across the Caribbean and experiences different environments, differential growth and mortality will undoubtedly play a role in Sargassum quantities and morphotype composition. A further potential limitation of this study arises from the simple classification of simulations into sub-origin/transport pathway A and B. This simple classification into two pathways points to resolvability of the complex problem of blooms; why now, why here, what are the dominant parameters that influence growth and mortality? Nevertheless, the findings of this study are relevant to the developing Sargassum industry by providing insights into the potential causes of variation in morphotype composition arriving in the Caribbean; and by extension provide a baseline for further studies on the predictability of seasonal patterns. The identification of the two sub-origins/transport pathways has implications for advancing the region’s understanding of the factors responsible for the continued proliferation and extensive interannual variability of Sargassum in the Tropical Atlantic since the initial bloom in 2011.
Methods
Sample collection and sorting
For each date of sampling effort, three clumps of newly beached, wet, “fresh gold” Sargassum were collected from Morgan Lewis beach, Barbados (13° 16′ 4.86′′ N–59° 33′ 48.41′′ W) (Fig. S3). Clumps (~ 0.27 kg each) were collected using both hands at approximately 10 m intervals along the shoreline. Using gross morphological features (following52), each of the three clumps was carefully separated into its component morphotypes, Sargassum natans I, S. natans VIII and S. fluitans III (Fig. 1). The displacement volume of each morphotype from each clump was then obtained using a measuring cylinder filled with a known volume of seawater. Sampling occurred opportunistically from February 2021 to March 2022, with at least one sample being collected each month. Sampling efforts resulted in 24 collection days over a 1-year period.
Backtracking of pelagic Sargassum
Satellite tracked mix-layer drifters (drogue element at 15 m) from the Global Drifter Program (GDP)53 have been deployed around the globe since ~ 1979. These drifters provide reliable tracking of water particles at drogue depth and are equipped with batteries that can last in excess of 450 days; however, loss of drogue is common. Using a Surface Velocity Program (SVP), data are provided on position, temperature and drogue on/off54. Current vector components are calculated at 6-h intervals from sequential positions and can be retrieved from https://www.aoml.noaa.gov/phod/gdp/. This study used a compiled file of the GDP data set consisting of: year, day, hour, longitude, latitude, east-current, west-current and drogue-on/drogue-off flag from 1979 to 2020. For tracking, the data were interpolated to a 1/12th degree resolution grid at 365 year-day intervals. The exact steps taken to achieve this dataset can be found in Ref.43.
To determine potential origins of the sampled Sargassum, each of the 24 collection days were backtracked using IDL 8.8.1 programming software (https://www.l3harris.com/all-capabilities/idl). Using a simple backtracking algorithm based on the drifter data set and 0.5% windage, Sargassum was tracked back 365 days from collection points on Morgan Lewis beach to determine the origin of each sample within the Tropical Atlantic. The selected 365-day time for backtracking was based on an experiment that assessed the dispersion of Sargassum from the NERR (Fig. S4). Our results showed that approximately 90% of the Sargassum population within the NERR is dispersed within 365 days. Recent studies44 also found that Sargassum present in the NERR has a high probability of entering the Caribbean within a year’s time.
Backtracks were simulated using 100 particles launched simultaneously from the collection location at Morgan Lewis beach, applying sub grid-scale turbulent motions (Lagrangian Stochastic Model55) to each particle’s current component:
where u′ is an adjusted current component and P(1) is a normal (Gaussian) random distribution with a mean of zero and a standard deviation of one. This simple turbulence addition to each of the east–west (u) and north–south (v) current components acknowledges that the gridded current database is smoother than reality. End points of each particle’s back-trajectory were obtained by center-of-mass calculations of the 100 ending locations.
The tracking methodology used in this study was developed by Ref.43 and is currently used to obtain 3-month Sargassum forecasts for the Lesser Antilles, published in the Sargassum Sub-Regional Outlook Bulletin (https://www.cavehill.uwi.edu/cermes/projects/sargassum/outlook-bulletin.aspx). Forecasts from the Outlook bulletin are well suited for monitoring Sargassum within the Lesser Antilles and are in close agreement with observed influxes56.
Data analysis
Monthly changes in relative abundance of Sargassum morphotypes (volume of morphotype/total volume of Sargassum sample) were calculated by averaging compositional sample data first by date (if multiple Sargassum samples were collected the same date) and then by month (if multiple dates were sampled within the same month). Differences in relative abundance of Sargassum morphotypes between the two sub-origins were tested using a PERMANOVA with the sample compositional data (transformed into a bivariate matrix following57 to address non-independence of the three percent estimates) as response matrix and sub-origin as independent factor, while implementing a constrained nested permutation scheme with sample data nested within dates and dates serving as independent statistical replicates for each sub-origin. This test was conducted using the adonis function of the “vegan” package in R58,59. Data were also checked for homogeneity of dispersion using the betadisper function of the same package (Table S1). Given that Sargassum compositional samples involved three morphotypes, we used ternary plots to display these data; these plots were produced using the “ggplot2”, “ggtern”, “ggpubr”, and “lattice” packages in R59,60,61,62,63.
Environmental data were retrieved using Giovanni (https://giovanni.gsfc.nasa.gov/giovanni/) time-series area-average oceanic data. The data sources were Sea Surface Temperature at 4 microns (Night) 8-daily 4 km (MODIS-Aqua) and Chlorophyll a concentration 8-daily 4 km (MODIS-Aqua MODISA_L3m_CHL_8d_4km). These data were retrieved for each collection date for (1) the location and time of origin (i.e., 365 backtracked days; 1 year) and the location and time of the mid-point (183 backtracked days; 6 months). For example, when assessing the initial SST conditions at time of origin for a sample collected on the 10th of January 2022 in Barbados, we used the 8-day SST average data from the closest available time intervals the year before (i.e., January 2021) at the estimated point of origin. To do this, a box of approximately 93,500 sq km was drawn centred around the point of origin. Then, the SST 8-day average for the entire box area was downloaded. To test for differences in SST and chlorophyll a between sub-origins a PERMANOVA test using SST and chlorophyll as response bivariate matrix data and time period (initial vs mid-way point) and sub-origin location (A vs B) (and their interaction) as independent factors. This test was also conducted using the adonis function of the “vegan” package in R58,59. Chlorophyll a data were square-root transformed to minimize the effect of extreme values. The significance level of 0.05 was used for all analyses.
Data availability
All data generated or analysed during this study are included in this published article [and its Supplementary Information files].
References
Huffard, C. L., von Thun, S., Sherman, A. D., Sealey, K. & Smith, K. L. Pelagic Sargassum community change over a 40-year period: Temporal and spatial variability. Mar. Biol. 161, 2735–2751. https://doi.org/10.1007/s00227-014-2539-y (2014).
Laffoley, D. A. et al. The Protection and Management of the Sargasso Sea: The Golden Floating Rainforest of the Atlantic Ocean (Sargasso Sea Alliance, 2011).
Martin, L. M. et al. Pelagic Sargassum morphotypes support different rafting motile epifauna communities. Mar. Biol. 168, 115. https://doi.org/10.1007/s00227-021-03910-2 (2021).
Butler, J. N. & Stoner, A. W. Pelagic Sargassum: Has its biomass changed in the last 50 years? Deep Sea Res. Part. Oceanogr. Res. Pap. 31, 1259–1264. https://doi.org/10.1016/0198-0149(84)90061-X (1984).
Lapointe, B. E. A comparison of nutrient-limited productivity in Sargassum natans from neritic vs oceanic waters of the western North Atlantic Ocean. Limnol. Oceanogr. 40, 625–633. https://doi.org/10.4319/lo.1995.40.3.0625 (1995).
Gower, J. F. R. & King, S. A. Distribution of floating Sargassum in the Gulf of Mexico and the Atlantic Ocean mapped using MERIS. Int. J. Remote Sens. 32, 1917–1929 (2011).
Gower, J., Young, E. & King, S. Satellite images suggest a new Sargassum source region in 2011. Remote Sens. Lett. 4, 764–773. https://doi.org/10.1080/2150704X.2013.796433 (2013).
Chávez, V. et al. Massive influx of pelagic Sargassum spp. on the coasts of the Mexican Caribbean 2014–2020: Challenges and opportunities. Water 12, 2908. https://doi.org/10.3390/w12102908 (2020).
Franks, J. S., Johnson, D. R. & Ko, D. S. Pelagic Sargassum in the tropical North Atlantic. Gulf Caribb. Res. 27, 6–11. https://doi.org/10.18785/gcr.2701.08 (2016).
Wang, M. et al. The great Atlantic Sargassum belt. Science 365, 83–87. https://doi.org/10.1126/science.aaw7912 (2019).
Johns, E. M. et al. The establishment of a pelagic Sargassum population in the tropical Atlantic: Biological consequences of a basin-scale long distance dispersal event. Prog. Oceanogr. 182, 102269. https://doi.org/10.1016/j.pocean.2020.102269 (2020).
Lapointe, B. E. et al. Nutrient content and stoichiometry of pelagic Sargassum reflects increasing nitrogen availability in the Atlantic Basin. Nat. Commun. 12, 3060. https://doi.org/10.1038/s41467-021-23135-7 (2021).
Skliris, N., Marsh, R., Appeaning Addo, K. & Oxenford, H. Physical drivers of pelagic Sargassum bloom interannual variability in the Central West Atlantic over 2010–2020. Ocean Dyn. https://doi.org/10.1007/s10236-022-01511-1 (2022).
van Tussenbroek, B. I. et al. Severe impacts of brown tides caused by Sargassum spp. on near-shore Caribbean seagrass communities. Mar. Pollut. Bull. 122, 272–281. https://doi.org/10.1016/j.marpolbul.2017.06.057 (2017).
Oxenford, H. A., Johnson, D., Cox, S.-A. & Franks, J. Report on the Relationships Between Sargassum Events, Oceanic Variables, and Dolphinfish and Flyingfish Fisheries 32 (University of the West Indies, 2019).
Ramlogan, N. R., McConney, P. & Oxenford, H. A. Socio-economic Impacts of Sargassum Influx Events on the Fishery Sector of Barbados 86 (University of the West Indies, 2017).
Caribbean Regional Fisheries Mechanism, Japanese International Cooperation Agency. Fact-Finding Survey Regarding the Influx and Impacts of Sargassum Seaweed in the Caribbean Region (CRFM-JICA, 2019).
Louime, C., Fortune, J. & Gervais, G. Sargassum invasion of coastal environments: A growing concern. Am. J. Environ. Sci. 13, 58–64. https://doi.org/10.3844/ajessp.2017.58.64 (2017).
Bartlett, D. & Elmer, F. The impact of Sargassum inundations on the Turks and Caicos islands. Phycology 1, 83–104. https://doi.org/10.3390/phycology1020007 (2021).
Resiere, D., Mehdaoui, H., Nevière, R. & Mégarbane, B. Sargassum invasion in the Caribbean: The role of medical and scientific cooperation. Rev. Panam Salud Pública 43, e52. https://doi.org/10.26633/RPSP.2019.52 (2019).
Resiere, D. et al. Sargassum seaweed health menace in the Caribbean: Clinical characteristics of a population exposed to hydrogen sulfide during the 2018 massive stranding. Clin. Toxicol. 59, 215–223. https://doi.org/10.1080/15563650.2020.1789162 (2021).
Devault, D. A. et al. The silent spring of Sargassum. Environ. Sci. Pollut. Res. 28, 15580–15583. https://doi.org/10.1007/s11356-020-12216-7 (2021).
United Nations Environment Programme - Caribbean Environment Programme. Sargassum White Paper—Turning the Crisis into an Opportunity. Ninth Meeting of the Scientific and Technical Advisory Committee (STAC) to the Protocol Concerning Specially Protected Areas and Wildlife (SPAW) in the Wider Caribbean Region (United Nations Environment Programme, 2021).
Maréchal, J. P., Hellio, C. & Hu, C. A simple, fast, and reliable method to predict Sargassum washing ashore in the Lesser Antilles. Remote Sens. Appl. Soc. Environ. 5, 54–63. https://doi.org/10.1016/j.rsase.2017.01.001 (2017).
Amador-Castro, F., García-Cayuela, T., Alper, H. S., Rodriguez-Martinez, V. & Carrillo-Nieves, D. Valorization of pelagic Sargassum biomass into sustainable applications: Current trends and challenges. J. Environ. Manag. 283, 112013. https://doi.org/10.1016/j.jenvman.2021.112013 (2021).
Desrochers, A., Cox, S.-A., Oxenford, H. A. & van Tussenbroek, B. I. Sargassum Uses Guide: A Resource for Caribbean Researchers (Centre for Resource Management and Environmental Studies, University of the West Indies, 2020).
Robledo, D. et al. Challenges and opportunities in relation to Sargassum events along the Caribbean Sea. Front. Mar. Sci. 8, 699664. https://doi.org/10.3389/fmars.2021.699664 (2021).
Thompson, T. M., Young, B. R. & Baroutian, S. Pelagic Sargassum for energy and fertiliser production in the Caribbean: A case study on Barbados. Renew. Sustain. Energy Rev. 118, 109564. https://doi.org/10.1016/j.rser.2019.109564 (2020).
Oxenford, H. A., Cox, S.-A., van Tussenbroek, B. I. & Desrochers, A. Challenges of turning the Sargassum crisis into gold: Current constraints and implications for the Caribbean. Phycology 1, 27–48. https://doi.org/10.3390/phycology1010003 (2021).
Amaral-Zettler, L. A. et al. Comparative mitochondrial and chloroplast genomics of a genetically distinct form of Sargassum contributing to recent “Golden Tides” in the Western Atlantic. Ecol. Evol. 7, 516–525. https://doi.org/10.1002/ece3.2630 (2017).
Siuda, A. N. S. et al. Genetic variation among morphological forms of pelagic Sargassum and associated hydroids. In 69th Gulf and Caribbean Fisheries Institute 220–221 (GCFI, 2017).
Godínez-Ortega, J. L., Cuatlán-Cortés, J. V., López-Bautista, J. M. & van Tussenbroek, B. I. A natural history of floating Sargassum species (Sargasso) from Mexico. In Natural History and Ecology of Mexico and Central America (ed. Hufnagel, L.) (IntechOpen, 2021).
González-Nieto, D. et al. Molecular assessment of the genus Sargassum (Fucales, Phaeophyceae) from the Mexican coasts of the Gulf of Mexico and Caribbean, with the description of S. xochitlae sp. Nov.. Phytotaxa. 461, 254–274. https://doi.org/10.11646/phytotaxa.461.4.3 (2020).
Wynne, M. J. Checklist of Benthic Marine Algae of the Tropical and Subtropical Western Atlantic: Fifth Revision (Schweizerbart Science Publishers, 2022).
Davis, D. et al. Biomass composition of the golden tide pelagic seaweeds Sargassum fluitans and S. natans (morphotypes I and VIII) to inform valorisation pathways. Sci. Total Environ. 762, 143134. https://doi.org/10.1016/j.scitotenv.2020.143134 (2020).
Davis, T. A., Volesky, B. & Vieira, R. H. S. F. Sargassum seaweed as biosorbent for heavy metals. Water Res. 34, 4270–4278. https://doi.org/10.1016/S0043-1354(00)00177-9 (2000).
Gobert, T. et al. Trace metal content from holopelagic Sargassum spp. sampled in the tropical North Atlantic Ocean: Emphasis on spatial variation of arsenic and phosphorus. Chemosphere 308, 136186. https://doi.org/10.1016/j.chemosphere.2022.136186 (2022).
Alleyne, K. S. T., Neat, F. & Oxenford, H. A. A baseline assessment of the epiphytic community associated with pelagic sargassum in the Tropical Atlantic. Aquat. Bot. 186, 103635. https://doi.org/10.1016/j.aquabot.2023.103635 (2023).
García-Sánchez, M. et al. Temporal changes in the composition and biomass of beached pelagic Sargassum species in the Mexican Caribbean. Aquat. Bot. 167, 103275. https://doi.org/10.1016/j.aquabot.2020.103275 (2020).
Machado, C. B. et al. Pelagic Sargassum events in Jamaica: Provenance, morphotype abundance, and influence of sample processing on biochemical composition of the biomass. Sci. Total Environ. 817, 152761. https://doi.org/10.1016/j.scitotenv.2021.152761 (2022).
Schell, J., Siuda, A. N. S. & Goodwin, D. S. Shipboard observation of pelagic Sargassum spp. reveals proliferation of a rare form and differences in associated mobile fauna community structure. In 68th Gulf and Caribbean Fisheries Institute 421–423 (2016).
Beron-Vera, F. J. et al. Dynamical geography and transition paths of Sargassum in the Tropical Atlantic. AIP Adv. 12, 105107. https://doi.org/10.1063/5.0117623 (2022).
Johnson, D. R., Franks, J. S., Oxenford, H. A. & Cox, S.-A.L. Pelagic Sargassum prediction and marine connectivity in the Tropical Atlantic. Gulf Caribb Res. 31, 20–30. https://doi.org/10.18785/gcr.3101.15 (2020).
Johns, W. E., Townsend, T. L., Fratantoni, D. M. & Wilson, W. D. On the Atlantic inflow to the Caribbean Sea. Deep Sea Res. Part. Oceanogr. Res. Pap. 49, 211–243. https://doi.org/10.1016/S0967-0637(01)00041-3 (2002).
Putman, N. F. et al. Simulating transport pathways of pelagic Sargassum from the Equatorial Atlantic into the Caribbean Sea. Prog Oceanogr. 165, 205–214. https://doi.org/10.1016/j.pocean.2018.06.009 (2018).
Hanisak, M. D. & Samuel, M. A. Growth rates in culture of several species of Sargassum from Florida, USA. Hydrobiologia 151, 399–404. https://doi.org/10.1007/BF00046159 (1987).
Magaña-Gallegos, E. et al. Growth rates of pelagic Sargassum species in the Mexican Caribbean. Aquat. Bot. 185, 103614. https://doi.org/10.1016/j.aquabot.2022.103614 (2023).
Lapointe, B. E., West, L. E., Sutton, T. T. & Hu, C. Ryther revisited: Nutrient excretions by fishes enhance productivity of pelagic Sargassum in the western North Atlantic Ocean. J. Exp. Mar. Biol. Ecol. 458, 46–56. https://doi.org/10.1016/j.jembe.2014.05.002 (2014).
Lapointe, B. E. Phosphorus-limited photosynthesis and growth of Sargassum natans and Sargassum fluitans (Phaeophyceae) in the western North Atlantic. Deep Sea Res. Part Oceanogr. Res. Pap. 33, 391–399. https://doi.org/10.1016/0198-0149(86)90099-3 (1986).
Johnson, D. R., Ko, D. S., Franks, J. S., Moreno, P. & Sanchez-Rubio, G. The Sargassum invasion of the eastern Caribbean and dynamics of the equatorial North Atlantic. In 65th Gulf and Caribbean Fisheries Institute 102–103 (2012).
Oviatt, C. A., Huizenga, K., Rogers, C. S. & Miller, W. J. What nutrient sources support anomalous growth and the recent Sargassum mass stranding on Caribbean beaches? A review. Mar. Pollut. Bull. 145, 517–525. https://doi.org/10.1016/j.marpolbul.2019.06.049 (2019).
Schell, J., Goodwin, D. & Siuda, A. Recent Sargassum inundation events in the Caribbean: Shipboard observations reveal dominance of a previously rare form. Oceanography 28, 8–10. https://doi.org/10.5670/oceanog.2015.70 (2015).
Lumpkin, R. & Pazos, M. Measuring surface currents with surface velocity program drifters: The instrument, its data, and some recent results. In Lagrangian Analysis and Prediction of Coastal and Ocean Dynamics (eds Kirwan, A. D. et al.) 39–67 (Cambridge University Press, 2007).
Johnson, D. R., Oxenford, H. A., Cox, S-A. & Franks, J. S. Holopelagic Sargassum and the complexities of predicting influxes and impacts on pelagic fisheries of the Lesser Antilles. In 71st Gulf and Caribbean Fisheries Institute 300–305 (2018).
Griffa, A. Applications of stochastic particle models to oceanographic problems. In Stochastic Modelling in Physical Oceanography. Progress in Probability Vol. 39 (eds Adler, R. J. et al.) (Birkhäuser, 1996).
Marsh, R., Oxenford, H. A., Cox, S.-A.L., Johnson, D. R. & Bellamy, J. Forecasting seasonal Sargassum events across the Tropical Atlantic: Overview and challenges. Front. Mar. Sci. 9, 914501. https://doi.org/10.3389/fmars.2022.914501 (2022).
Vermeesch, P. Exploratory analysis of provenance data using R and the provenance package. Minerals 9, 193. https://doi.org/10.3390/min9030193 (2019).
Oksanen, J. et al. vegan: Community Ecology Package. R Package Version 2.5-3. https://CRAN.R-project.org/package=vegan (2018).
R Core Team. R: A Language and Environment for Statistical Computing. https://www.R-project.org/ (R Foundation for Statistical Computing, 2022).
Wickham, H. ggplot2: Elegant Graphics for Data Analysis (Springer, 2016).
Hamilton, N. E. & Ferry, M. ggtern: Ternary diagrams using ggplot2. J. Stat. Softw. 87, 1–17. https://doi.org/10.18637/jss.v087.c03 (2018).
Kassambara, A. ggpubr: 'ggplot2' Based Publication Ready Plots. R Package Version 0.4.0. https://CRAN.R-project.org/package=ggpubr (2020).
Sarkar, D. Lattice: Multivariate Data Visualization with R (Springer, 2008).
Acknowledgements
The authors thank Makeda Corbin, Micaela Small and Amy Cox for help in collecting and sorting Sargassum samples and further acknowledges Sarah Mahadeo for contributing the map of the Wider Caribbean Region. This work would not have been possible without the generous funding by The Nippon Foundation of the World Maritime University (WMU)—Sasakawa Global Ocean Institute as well as for The Nippon Foundation & WMU-Sasakawa Global Ocean Institute’s ‘Closing the Circle Programme: Marine Debris, Sargassum and Marine Spatial Planning’ in the Eastern Caribbean.
Funding
This work was supported by The Nippon Foundation of the World Maritime University (WMU)—Sasakawa Global Ocean Institute as well as The Nippon Foundation and WMU-Sasakawa Global Ocean Institute’s ‘Closing the Circle Programme: Marine Debris, Sargassum and Marine Spatial Planning’ in the Eastern Caribbean.
Author information
Authors and Affiliations
Contributions
K.A., H.O. and F.N. conceived the project. K.A. collected the Sargassum compositional data and wrote the first draft. K.A. and D.J. generated the Sargassum backtrack data. K.A. and. H.V. analysed and interpreted the data. All authors contributed to the writing of the article and approved the submitted version.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Alleyne, K.S.T., Johnson, D., Neat, F. et al. Seasonal variation in morphotype composition of pelagic Sargassum influx events is linked to oceanic origin. Sci Rep 13, 3753 (2023). https://doi.org/10.1038/s41598-023-30969-2
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-023-30969-2
This article is cited by
-
In vitro antiherpetic activity of compounds extracted from Sargassum fluitans (Børgesen) Børgesen. Identification and in silico evaluation of potential antiviral molecules in a hydroalcoholic extract
Journal of Applied Phycology (2024)
-
FTIR-ATR chemometric analysis on pelagic Sargassum reveals chemical composition changes induced by cold sample transportation and sunlight radiation
Journal of Applied Phycology (2024)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
