
Abstract
Malaria due to the Plasmodium falciparum parasite remains a threat to human health despite eradication efforts and the development of anti-malarial treatments, such as artemisinin combination therapies. Human movement and migration have been linked to the propagation of malaria on national scales, highlighting the need for the incorporation of human movement in modeling efforts. Spatially couped individual-based models have been used to study how anti-malarial resistance evolves and spreads in response to drug policy changes; however, as the spatial scale of the model increases, the challenges associated with modeling of movement also increase. In this paper we discuss the development, calibration, and validation of a movement model in the context of a national-scale, spatial, individual-based model used to study the evolution of drug resistance in the malaria parasite.
Similar content being viewed by others

Introduction
The promulgation of the Millennium Development Goals, and specifically Goal 6 to combat HIV/AIDS, malaria, and other diseases, by the United Nations has resulted in a renewed global interest in malaria eradication. However, despite the efforts of the past 20 years, malaria caused by the Plasmodium falciparum parasite remains a serious public health concern with 241 million cases and 627 thousand deaths estimated in 20201. One significant barrier to eradication efforts is the evolution of anti-malarial resistance by the parasite, with resistance to the artemisinin components of artemisinin combination therapies (ACTs) being of particular concern1. While the primary driver of anti-malarial resistance is the evolutionary pressure applied on the parasite through the use of anti-malarial treatments (e.g., ACTs)2,3,4, a resistant parasite may also appear in a region due to importation though human movement (e.g., temporary travel for work or leisure) or migration (i.e., permanent relocation)5.
One means of studying the evolution of anti-malarial resistance—and the impact that various drug policies may have upon it—has been through the use of spatially coupled, stochastic, individual-based models (IBMs) that incorporate components such as: transmission of the parasite, immune acquisition and response, genotype evolution, and drug intervention strategies6. By incorporating space and geography in these models, it is possible to evaluate possible drug interventions and observe how geography and population distribution factors in to the development of anti-malarial resistance by the parasite. This insight may allow for new eradication strategies to be developed, as well as proper allocation of resources based upon projected movement patterns. Accordingly, modeling efforts must be accompanied by a model of human movement in order to account for the movement of anti-malarial resistant parasites via infected carriers. However, national scale simulations may incorporate millions of simulated individuals, spread across thousands of cells (representing the simulated space of a region or country), resulting in challenges of model implementation, calibration, and validation.
Since human movement and migration has been linked to malaria transmission and the migration of drug resistant genotypes5,7, incorporation of human movement is a critical component of such IBMs. The two common mathematical models of human movement used in studies of the Sub-Saharan Africa (SSA) region are gravity and radiation models8,9. Gravity models are constructed with the assumption that movement rates between two points (e.g., cities) increase in relation to the size of the point populations and decrease with the square of the distance between term; similar to physical laws of gravitation. Gravity models may also be modified with functions that account for other parameters, such as mode and cost of travel. Likewise, radiation models draw their inspiration from physical processes (i.e., particle diffusion), but differ from gravity models in that the underlying assumption is that individuals move outward from their origin and are “absorbed” by a given destination, within a given distance, with a probability that is proportionate to the population of a given destination.

Prevalence of P. falciparum malaria in Burkina Faso based the Malaria Atlas Project projections for 201710. Map prepared by the authors using ArcGIS Pro (version 2.9.3, https://www.esri.com/en-us/arcgis) using administrative boundaries from the World Bank Group11,12, and prevalence data from the Malaria Atlas Project10.
In this study we examine the data and mathematical models of human movement in Burkina Faso. Burkina Faso is a landlocked country in western SSA with endemic malaria (i.e., persistent transmission of the infection in the population) (Fig. 1), which is the leading cause of hospitalization in the general population (45.8% of cases) as well as for children under five (48.2% of cases)13,14. During periods of seasonal rainfall, the P. falciparum prevalence rate in the two- to ten-year-old population (called PfPR2-10) may increase significantly in relation to the annual mean PfPR2-10 which ranges from 8.0 to 67.6% based upon 2017 estimates10. The primary first-line therapy for uncomplicated malaria are ACTs such as artemether-lumefantrine (AL), which has been recommended since 2005 in accordance with WHO guidelines14. ACTs are used in conjunction with other malaria intervention programs such as: insecticide-treated mosquito nets (ITNs), indoor residual spraying (IRS), intermittent preventative treatment of pregnant women (IPTp), and seasonal malaria chemoprophylaxis15. As a result of ACT usage and these interventions, the general trend since 2005 has been a reduction in the prevalence of malaria16.

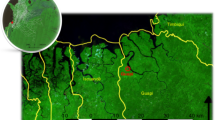
Sites of movement survey (red stars) by Marshall et al.17 overlaid population density from WorldPop18 at a with a resolution of 3 arc. White areas indicate no population density due to water features, while progressively darker shades indicate higher population density. Note the high population density of the region surrounding the survey site in the capital, Ouagadougou in comparison to the agricultural and trade hub Boussé, or agricultural village of Saponé. Map prepared by the authors using ArcGIS Pro (version 2.9.3, https://www.esri.com/en-us/arcgis) using administrative boundaries from the World Bank Group11 and population data from WorldPop18.
Surveys of human movement and migration in Burkina Faso are limited17,19,20,21,22, with Marshall et al.17 being the most recent. In their study, three sites were selected: the capital Ouagadougou in Kadiogo province, the agricultural and trade hub Boussé in Kourweogo province, and the agricultural village Saponé in Bazega province (Fig. 2). All sites were sampled during the rainy season of July 2011, with the number of trips recorded based upon the source and destination. Despite being the most recent quantitative study performed, some limitations are present. First, the section of survey sites is clustered around Ouagadougou, the centrally located capital of Burkina Faso. Second, complete coverage of travel to all provinces in Burkina Faso was not captured. Finally, the timing of the survey during the rainy season likely also introduces biases due to seasonal migration patterns.
However, a sufficient quantity of data was collected by Marshall et al.17 that it was possible for the authors fit the data using a gravity model and radiation model of movements8. Upon completion of model fit, the authors concluded that a gravity model with a power-law distance kernel had the most predictive power in relation to the observed movement patterns. This is consistent with previous studies in SSA that found that radiation models under predict travel compared to gravity models9. However, in the context of SSA it has been suggested that countries may follow unique patterns that are not perfectly explained by either the gravity or radiation model in their unmodified forms9.
Results
Mathematical model
The general success and application of gravity models for human movement and migration in SSA8,9, coupled with the recent work by Marshall et al.8,17, motivated us to start with the modified gravity model suggested:
where \({\text{Pr}}(j|i)\) describes the probability of travel from the source i to the destination j, given the product of the population of j raised to τ (Eq. 1), and the distance kernel (Eq. 2), which takes the form of a power law containing the scale parameter ρ, and the power-law parameter α.
Two limitations are present in the model constructed by Marshall et al.8 First, the model presumes that an entire destination (i.e., destination city) can be treated as a point, resulting in an incongruence when used in the context of a grid-based landscape. Second, is the lack of consideration for the time, distance, or complexity in traveling to a given destination. This is particularly important in the context of malaria interventions since rural communities may lack local medical resources, necessitating travel to seek care5,23. Accordingly, this limitation can be addressed by capturing the difficulty associated with travel through the use of a friction surface, which quantifies the ease or difficulty in traversing surfaces (e.g., road types) or natural barriers such as mountainous terrain24. An alternative is the use of a travel time map (or surface) which estimates the time to reach the nearest city (or high-density urban area) from a given location on the map25. Since travel time maps typically annotate the travel time within an urban environment as zero, this also addresses the incongruence that arises since urban boundaries are delineated travel times of zero.
Accordingly, the probability of movement can be adjusted to use travel times as follows:
where \({\text{Pr}}(j|i)\prime\) describes the new probability of travel from source i to the destination j following the division of the original probability \({\text{Pr}}(j|i)\) by the sum of one plus travel time to the nearest city of the source ti and destination tj. Incorporation of both the source ti and destination tj in the denominator of Eq. (3) ensures accounting for the costs of indirect travel routes. This has the effect of biasing the model towards destination cells that are located in (i.e., travel time of zero) or near (i.e., low travel time to the nearest city) cities, but still allowing travel between rural locations, or from the city to a rural location. Note that since within city travel is unpenalized (i.e., travel time of the source and destination is zero), during model calibration, care must be taken to ensure that the number of trips (e.g., within a city or province, or between cities or provinces) are properly accounted for. If necessary, \({\text{Pr}}(j|i)\prime\) can be adjusted by dividing by a penalty parameter, p, in the event the model is biased towards movement remaining within a given area.
Evaluation of implemented model
The mathematical model developed—that is Eqs. (1–3)—was implemented as part of a stochastic IBM developed for Burkina Faso6,26, and parameterization of the movement equations was performed by first estimating the best fit for the ρ based upon the \(log_{e} \left( \rho \right)\) from the range of values suggested by Marshall et al.8, with \(log_{e} \left( \rho \right) = 0.20\) having the best fit to the survey data17. Next, the inferred values \(\alpha = 1.27\) and \(\tau = 1.342\), as fit by Marshall et al.8, were used in an IBM of Burkina Faso for one month with a population of approximately 19 million individuals, and all travel between cells was logged. The cellular movement of the simulation was aggregated to the province level and compared to the Marshall et al.17 survey data. Initial calibration results revealed a model bias in Kadiogo province—containing the capital Ouagadougou—due to individual movement remaining within the province as a result of the high population relative to the rest of the country. This was corrected by dividing Pr’ by the model penalty \(p = 12\) when the cell is within Kadiogo province, where the penalty value was found though model fitting. As a result, it was found that the movement probability \({\text{Pr}}(j|i)\prime\), coupled with a model penalty applied to Kadiogo Province, produces a reasonable replication of the source to destination movement in regions of Burkina Faso with high, geographically dispersed populations (Fig. 3).

Comparison of the gravity model with kernel function parameterized by Marshall et al.8 versus the mathematical model with travel surface and parameterization described, as implemented in the individual-based model. (a) The mean number of trips for ten replicates to the destination cell in a single month using the parametrization prepared by Marshall et al.8 (b) The mean number of trips for ten replicates to the destination cell in a single month using the mathematical model with travel surface and parametrization described in this manuscript. (c) The population distribution of Burkina Faso from WorldPop18 aggregated into the same 5 km-by-5 km cells used for movement. (d) The raster algebra difference between the mean number of trips to the destination cell from (a,b); note that the Marshall et al.8 model tends to project higher movement to rural areas (negative values in blue tones) compared to (b) which is shifted towards the areas of higher population using the method described here (positive values in red tones). Furthermore, the movement is less diffused compared to (a). However, the lack of diffusion results in movement being better aligned with the concentrations of population and transportation networks compared to (a) despite the total number of trips being similar. Maps prepared by the authors using ArcGIS Pro (version 3.0.3, https://www.esri.com/en-us/arcgis) using administrative boundaries from the World Bank Group11 and data from the simulation described.
Evaluation of the projected movement was performed using the mean of ten replicates for six different configurations (i.e., Marshall et al.8 model with published calibration, biased towards shorter travel, and our model fit; along with the model described here and the three parameterizations), all of which produced a similar number of trips (1,898,288 ± 555) (see Supplemental Information 1, Fig. S1). When comparing the gravity model and parameterization suggested by Marshall et al.8 it is clear that the projected population movement (Fig. 3a) is diffuse with smaller population centers being bypassed by individuals in favor of Ouagadougou. However, the results of the mathematical model with travel surface (Fig. 3b) show trips that are aligned with the population distribution of Burkina Faso (Fig. 3c). Furthermore, the gravity model appears to under project the number of trips to major population centers while over projecting the number of trips to areas of low population. (Fig. 3d).To further contrast the differences in travel, an additional forty replicates (total n = 50) were run for the Marshall et al.8 model with published calibration, and the model described here with the best model fit (see Supplemental Information 2). Both models produce similar results for low population cells (i.e., < 1000); however, as expected, the models diverge significantly as the population increases (see Supplemental Information 1, Fig. S4).
Finally, when the total number of trips is compared to the distance traveled, the results are also comparable to the survey data (Fig. 4). However, given the sparsity of survey data for Kourweogo and Bazega (Fig. 4), the projected short distance travel is consistent with the survey data and long-distance travel—which has a lower overall frequency resulting in fewer survey respondents—is generally consistent with similar travel out of Kadiogo province. To reduce the effects of the long model tails seen in the bottom two panels of Fig. 4, an alternative value of \(log_{e} \left( \rho \right) = 0.45\) was selected. The difference was small with slightly less travel to population centers compared to the parameterization suggested by Marshall et al. or the \(log_{e} \left( \rho \right) = 0.20\) value (see Supplemental Information 1, Figs. S1, S2).

Trips from Kadiogo providence compare favorably with the Marshall et al.17 survey on the basis of distance traveled. However, the limited survey data for Kourweogo and Bazega pose a challenge to model validation, although the simulated results are generally consistent with the survey data.
Discussion
With individually-based models (IBMs) designed for, and calibrated to, epidemiological models of malaria, an appropriate model of movement by individuals is necessary due to the role that human movement and migration plays in the spread of anti-malarial resistance. However, the development, calibration, and validation of movement models, particularly at the national scale, remains a challenge. These challenges can be further compounded by the lack of quantitative data available for a given country of interest. Despite this, IBMs and mathematical modeling more broadly, still play an important role in malaria control with national scale models offering possible insights for surveillance and drug policy response27.
Despite these challenges, geospatially coupled malaria models are adventitious in that they can produce projections for where anti-malarial resistance may arise, allowing surveillance efforts to focused on narrow geographic scopes. An important application of malaria IBMs is evaluation of how changes in drug policy (e.g., changing the partner drug in an ACT or introducing multiple first-line therapies6,26,28,29) impact the emergence of drug resistance by the parasite, and the connection between antimalarial drugs and de novo emergence of resistance has been well established2,3,4, accordingly it is not a matter of if drug resistance will emerge, but rather when. However, the role that human migration plays in introducing an anti-malarial resistant parasite to a region remains unclear.
A standard movement model, or a regionally calibrated movement model such as the one developed by Marshall et al.8, is not guaranteed to meet the needs of a model developer “out of the box.” While a published parameterization may offer a useful starting point for model developers, additional algorithm development, parameterization, and calibration steps are necessary to ensure that the model is appropriate for expected movement in a region, and the goals of the model. As this paper has demonstrated, one approach to improving model fidelity is through the use of a travel time map. Although, as always, model developers should be diligent during the calibration and validation process to ensure that model outputs make sense in the context of quantitative and qualitative data that is available.
While the introduction of an anti-malarial resistant P. falciparum parasites to a region through human movement and migration is just one mechanism by which resistance can appear; it is necessary that IBMs modeling malaria with epidemiological goals properly account for it. As this paper has demonstrated, it is possible to “scale-up” mathematical models to be utilized in national-scale IBMs; however, performance, calibration, and validation are all challenges that require careful investigation. Furthermore, the availability of data for a region of study can place limitations on the extent of validation that is possible, necessitating some caution in the claims that can be made by simulating specific scenarios.
Methods
Implementation
The mathematical model described was implemented using C++, in an IBM previously developed by Nguyen et al.6 During execution of the simulation, individual movement is based upon a decision to (i) leave the current location, and (ii) where to travel to at the end of each timestep—representing one day in this instance—as part of the following operations. First, geographic data (i.e., population, travel times, etc.) is read into the simulation state. On each simulation timestep the population events (e.g., infections, births, deaths, etc.) are processed for each cell in the simulation, and the number of individuals in the cell is determined. Next, the individuals in the cell are selected for travel based upon the overall probability of movement in a given year (see Calibration and Validation). This is followed by the calculation of \({\text{Pr}}(j|i)\prime\) for the current cell i to each other cell in the simulation, resulting in a vector from which a random, multinomial draw is performed for each individual selected for movement, indicating their destination cell. Individuals are then moved to their destination cell at the end of the time step—ensuring that the population of the cells remain synchronized—and a timer is set to indicate if, and when, they will return to their original cell. If no timer is set, then they remain in their current cell.
The incorporation of the movement model resulted in a number of time and space complexity challenges. Analysis of the source code suggests an algorithmic complexity of \(O\left( {n^{3} } \right)\) with a space complexity of \(O\left( n \right)\). In order to ensure efficient use of resources, static memory along with the singleton design pattern was used to eliminate the need to reload or copy geographic data, which is represented in memory as either a matrix or a flat array. While it was originally hypothesized during development that the probability of movement could be calculated once at model initialization based upon the initialization parameters, doing so resulted in artifacts appearing in simulation movement over time. However, when the probability is calculated during each timestep (i.e., in a manner consistent with population growth in the simulation) the model performed as expected suggesting that caching of calculated movement probabilities is unlikely to be possible. This suggests that most optimizations are likely to come from careful programming and code organization to minimize the number of times that movement probabilities need to be calculated per timestep.
Finally, in preparing the spatial data used for the calibration of Burkina Faso, the limiting factor of the spatial resolution is the cellular size of the reference PfPR2-10 values provided by the Malaria Atlas Project10, resulting in a cell size of 25 km2. This results the approximately 273,000 km2 of Burkina Faso being converted into 10,936 pixels (or cells). Initial population data comes from WorldPop18, which was aggregated using ArcMap 10.7.1–25 km2 cells, resulting in pixel level populations ranging from 1 to 206,607 individuals per cell. The travel time are derived from Malaria Atlas Project travel time to cities25, with the original 1 km2 cells aggregated on the basis of the cellular mean to 25 km2.
Calibration and validation
Model calibration begin by fitting the scale parameter ρ, based upon the range of values suggested by Marshall et al.8 who noted that improvements upon it may be possible. To fit ρ, a “synthetic survey” was programmed using MATLAB 2019b. The script approximated the sampling of the Marshall et al.17 by drawing a destination for each trial from a given province (i.e., Kadiogo, Kourweogo, and Bazega) by applying the gravity model fit by Marshal et al.8 (Eqs. 1 and 2). The distance (\(d_{i,j}\)) was based upon the distance between the centroid of each combination of providence, calculated using ArcMap 10.7.1, while the population of the province (\(Pop_{j}\)) was calculated using WorldPop data18. The TLAB script then iterated though the iterating through values 0.05 to 1.8 by steps of 0.05 for \(log_{e} \left( \rho \right)\) taking a random draw from source i to destination j based upon a random draw of samples equal to the number of survey participants traveling from each province (Kadiogo n = 403, Kourweogo n = 503, Bazega n = 387). This was repeated for a total of 1,000 trials for each value to ensure sufficient statistical power.
The results of the synthetic survey were then compared to the survey data from Marshall et al.17 and the number of matches and inter-quartile ranges (IQR) were compared. While Marshall et al.8 suggests \(log_{e} \left( \rho \right) = 0.54\) as the best fit, the value \(log_{e} \left( \rho \right) = 0.20\) was selected since it offered the most IQR matches along with a low mean squared error. As an alternative, \(log_{e} \left( \rho \right) = 0.45\) was noted to evaluate the impact of biasing the model fit towards the longer trips, with minimal impact upon the overall results (see Supplemental Figs. S1 and S2). To further validate the use of \(log_{e} \left( \rho \right) = 0.20\), the parameter was used as a simulation parameter in the IBM, with the frequency of trips and distanced traveled tabulated. The simulation results compared favorably to the survey data, although some gaps in the survey data preclude a complete one-to-one comparison of simulation results to survey data.
Next, the model bias resulting from travel departing from Kadiogo province—containing the capital of Burkina Faso—remaining within the province, was corrected by determining the penalty p to be applied to \({\text{Pr}}(j|i)\prime\). Based upon a parameter space search in which simulation was run with varying values for p a 12-fold intra-province travel penalty was found to be sufficient to eliminate the bias. This was again validated by comparing simulation results against the survey data.
Finally, the precise number of trips was calibrated starting with the finding by Marshall et al.17 that about 29.1% of the population of Burkina Faso travels on a yearly basis with a mean of 3.42 trips. This suggests a daily circulation rate (i.e., the daily probability of an individual traveling) of 0.002727 for travelers leaving their home province. However, since the model (and simulation) allows for intra-province travel, it is necessary to scale this value up to ensure that inter-provincial travel is correct. Using the simulation with the aforementioned calibration parameters, individual movement was tracked in the model and various circulation rates trialed until the national level annual rates (29.1%) were matched by a daily circulation rate of 0.00336. As a result of this rate, approximately 19% of all trips remain within a province (i.e., travel between cells in the same province) while the remainder leave. A final set of trials were performed using the simulation and the calibrated parameters, resulting in simulated national-scale movement that is largely consistent with survey data (Fig. 3).
Data availability
The datasets generated and/or analyzed during the current study are available in the malariaibm-movement-BurkinaFaso-2022 repository, https://github.com/bonilab/malariaibm-movement-BurkinaFaso-2022. The source code for the simulation and/or data analysis can be found under the Source directory. The raster data produced by the simulation and used to produce Fig. 3a,b can be found under the Data/GIS directory while processed survey data from Marshall et al.17 and intermediary simulation results used to produce Fig. 4 is found under Data/Movement.
References
World Health Organization. World Malaria Report 2021 (World Health Organization, 2021).
D’Alessandro, U. & Buttiëns, H. History and importance of antimalarial drug resistance. Trop. Med. Int. Health 6, 845–848 (2001).
Pongtavornpinyo, W. et al. Probability of emergence of antimalarial resistance in different stages of the parasite life cycle. Evol. Appl. 2, 52–61 (2009).
White, N. & Pongtavornpinyo, W. The de novo selection of drug–resistant malaria parasites. Proc. R. Soc. B 270, 545–554 (2003).
Alegana, V. A., Wright, J., Pezzulo, C., Tatem, A. J. & Atkinson, P. M. Treatment-seeking behaviour in low- and middle-income countries estimated using a Bayesian model. BMC Med. Res. Methodol. 17, 67 (2017).
Nguyen, T. D. et al. Optimum population-level use of artemisinin combination therapies: A modelling study. Lancet Glob. Health 3, e758–e766 (2015).
Marshall, J. M., Bennett, A., Kiware, S. S. & Sturrock, H. J. W. The hitchhiking parasite: Why human movement matters to malaria transmission and what we can do about it. Trends Parasitol. 32, 752–755 (2016).
Marshall, J. M. et al. Mathematical models of human mobility of relevance to malaria transmission in Africa. Sci. Rep. 8, 7713 (2018).
Wesolowski, A., O’Meara, W. P., Eagle, N., Tatem, A. J. & Buckee, C. O. Evaluating spatial interaction models for regional mobility in Sub-Saharan Africa. PLoS Comput. Biol. 11, e1004267 (2015).
Weiss, D. J. et al. Mapping the global prevalence, incidence, and mortality of Plasmodium falciparum, 2000–17: A spatial and temporal modelling study. Lancet 394, 322–331 (2019).
World Bank Group. Burkina Faso District Boundary. (2018).
World Bank Group. World Boundaries GeoDatabase. (2020).
World Health Organization. World Malaria Report 2019 (World Health Organization, 2019).
Ministère de la Santé. État de Santé de la Population du Burkina Faso: Rapport 2019. 1–88 https://drive.google.com/uc?export=dowload&id=1K-Aw5Xp5eWH-DSd_VbGT85pi84UWptpC (2020).
Kirakoya-Samadoulougou, F., De Brouwere, V., Fokam, A. F., Ouédraogo, M. & Yé, Y. Assessing the effect of seasonal malaria chemoprevention on malaria burden among children under 5 years in Burkina Faso. Malar. J. 21, 143 (2022).
President’s Malaria Initiative. Burkina Faso Malaria Operational Plan FY 2019. 1–51 https://www.pmi.gov/docs/default-source/default-document-library/malaria-operational-plans/fy19/fy-2019-burkina-faso-malaria-operational-plan.pdf?sfvrsn=3 (2019).
Marshall, J. M. et al. Key traveller groups of relevance to spatial malaria transmission: A survey of movement patterns in four sub-Saharan African countries. Malar. J. 15, 200 (2016).
WorldPop. Global High Resolution Population Denominators Project. (2018).
Henry, S., Boyle, P. & Lambin, E. F. Modelling inter-provincial migration in Burkina Faso, West Africa: The role of socio-demographic and environmental factors. Appl Geogr 23, 115–136 (2003).
Jonas Østergaard Nielsen. I’m Staying! Climate Variability and Circular Migration in Burkina Faso. In Environmental Change and African Societies vol. 5 121–148 (Brill, 2019).
Seid, E. Regional Integration and Trade in Sub-Saharan Africa, 1993–2010: An Augmented Gravity Model. In Regional Integration and Trade in Africa 91–108 (Palgrave Macmillian, 2015).
Sorichetta, A. et al. Mapping internal connectivity through human migration in malaria endemic countries. Sci. Data 3, 160066 (2016).
O’Meara, W. P. et al. The impact of primary health care on malaria morbidity – defining access by disease burden. Trop. Med. Int. Health 14, 29–35 (2009).
Nikolov, M. et al. Malaria elimination campaigns in the lake Kariba region of Zambia: A spatial dynamical model. PLoS Comput. Biol. 12, e1005192 (2016).
Weiss, D. J. et al. A global map of travel time to cities to assess inequalities in accessibility in 2015. Nature 553, 333–336 (2018).
Zupko, R. J. et al. Long-term effects of increased adoption of artemisinin combination therapies in Burkina Faso. PLOS Glob. Public Health 2, e0000111 (2022).
McKenzie, F. E. & Samba, E. M. The role of mathematical modeling in evidence-based malaria control. Am. J. Trop. Med. Hyg. 71, 94–96 (2004).
Boni, M. F., White, N. J. & Baird, J. K. The community as the patient in malaria-endemic areas: Preempting drug resistance with multiple first-line therapies. PLoS Med. 13, e1001984 (2016).
Boni, M. F., Smith, D. L. & Laxminarayan, R. Benefits of using multiple first-line therapies against malaria. Proc. Natl. Acad. Sci. USA 105, 14216 (2008).
Acknowledgements
This work was supported by the National Institutes of Health (R01AI153355) and the Bill & Melinda Gates Foundation Grant OPP159934 to the University of Washington and INV-005517. Simulations described in this study were performed on the Pennsylvania State University’s Institute for Computational and Data Sciences’ Roar supercomputer. AW is funded by a Career Award at the Scientific Interface by the Burroughs Wellcome Fund, the National Library of Medicine (DP2LM013102), and NIAID/NIH Grant 1R01AI160780-01. JG received funding from the Gates Foundation (INV-002092).
Author information
Authors and Affiliations
Contributions
R.J.Z. and M.F.B. designed the study. R.J.Z. and T.D.N. implemented the model. R.J.Z. calibrated and validated the model. R.J.Z. wrote the first draft of the paper and prepared all figures. J.G. and A.W. assisted with the original model development and validation of the model results. All authors contributed to the editing of the manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Zupko, R.J., Nguyen, T.D., Wesolowski, A. et al. National-scale simulation of human movement in a spatially coupled individual-based model of malaria in Burkina Faso. Sci Rep 13, 321 (2023). https://doi.org/10.1038/s41598-022-26878-5
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-022-26878-5
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
