
Abstract
Early Cretaceous floras containing angiosperms were described from several geographic areas, nearly from the Arctic to the Antarctic, and are crucial to understand their evolution and radiation. However, most of these records come from northern mid-latitudes whereas those of lower paleolatitude areas, such as the Crato Fossil Lagerstätte in NE Brazil, are less studied. Here, we describe from this region of northern Gondwanan origin, two fossil-species of eudicots belonging to a new extinct genus Santaniella gen. nov. Together with several vegetative axes and leaves, anatomically well-preserved fruits with seeds and persistent perianth-like organs allowed us to reconstruct its potential affinities with ranunculids, and presumably Ranunculaceae. Previous records putatively assigned to Ranunculales are all from mid-latitudes, and their first unequivocal occurrence in a low-latitude area supports further the hypothesis of a widespread radiation of the earliest diverging eudicot lineage by this early age.
Similar content being viewed by others


Introduction
The apparently rapid diversification of flowering plants in fossil records was famously referred to as an “abominable mystery” and “most perplexing phenomenon” by Darwin, who often advocated for a gradual evolution1. Nowadays, the fossil record demonstrates that the origin and early radiation of angiosperms was indeed less abrupt than previously thought2. Based on extensive evidence, we know now that the first flowering plants appeared at low-latitude areas in Northern Gondwana in the Early Cretaceous (ca. 135 Ma), and then they gradually began to disperse to mid- and high-latitudes in both hemispheres throughout the Cretaceous3. However, the inherently incomplete fossil record still hinders a precise reconstruction of the early evolutionary history of angiosperms and diversification of the main extant lineages, such as the eudicots.
Eudicots are an extremely diverse monophyletic group encompassing about three quarters of all living flowering plants4. They comprise a grade of early-diverging lineages, i.e., Ranunculales, Proteales, Trochodendrales and Buxales, that are successively sisters to the rest5. However, while the resolution of their phylogenetic relationships and systematic affinities have been constantly improved over the past 30 years, their origin and early evolutionary history are still elusive because our knowledge is based mostly on dispersed pollen and mesofossils, whereas the macrofossil record is scarce and mostly fragmented2,6.
The oldest record of eudicots extends back to at least the middle Early Cretaceous mainly in low-latitude areas and based mostly on their distinct tricolpate pollen grains, found in sedimentary rocks since the late Barremian (ca. 125 Ma)2. In addition, their presence in the Barremian and Aptian is strongly supported not only by abundant dispersed pollen grains, but also by the presence of in situ pollen in mesofossils including isolated stamens and carpels2,7. In contrast, unambiguous pre-Albian (before 113 Ma) macrofossils are relatively rare, whereas the diversity of eudicots seems to increase mainly during the Albian and Cenomanian (113–94 Ma) until they eventually start to dominate most plant fossil assemblages from the Late Cretaceous (ca. 100 Ma) onwards2.
The accelerated evolution and diversification of basal eudicots are expected during the late Barremian and early Aptian (ca. 125 Ma)8. In early-diverging eudicots, Ranunculales are sisters to all other lineages and like Proteales, Trochodendrales and Buxales represented by a handful of fossil records in the Early Cretaceous, most of which are not affiliated to extant families2,5. To date, molecular estimates using fossil-based calibrations indicate that the emergence of the crown-group of Ranunculales started only around 123 to 112 Ma4,9,10 and the origin of the crown group of Ranunculaceae at around 108 Ma11. In addition, the few undisputed macrofossils with ranunculid/basal eudicot affinities are derived from mid-paleolatitudes in the Northern Hemisphere12,13,14,15,16,17,18,19,20,21,22,23,24, whereas the low-paleolatitudes, that may have been the cradle for eudicots, are still mostly “terra incognita”.
Northern Gondwanan floras (e.g., Crato Formation flora), which represent the paleoequatorial vegetation during the Early Cretaceous, provide important insights in the evolution and early radiation of angiosperms and their biology. In the Crato Formation from Brazil, one-third of the total angiosperm pollen record is represented by tricolpate grains25, but no unquestionable eudicot megafossil was described yet (see discussion6).
Here, we report for the first time specimens whose exceptional preservation of vegetative and reproductive parts supports affinities with extant Ranunculales, and thus, also represent the most complete ranunculid fossils described to date from the Early Cretaceous tropics.
Details of specimens
Geological setting and age
The study specimens are from the Crato limestones termed Pedra Cariri within a lithostratigraphic unit known as the Crato Formation, belonging to the Santana Group in the Jurassic-Cretaceous Araripe Basin (NE Brazil)26. They have been gathered at open cast pits close to the town of Nova Olinda (Ceará) by local workers who excavate the laminated lacustrine carbonate rock as cut and ornamental stone. Recent dating of other Santana Group formations assumed an early Aptian or even late Barremian age for the Crato Formation27,28. However, this age is not consensual since former estimations suggested a late Aptian/early Albian age of this geologic unit (29 and references therein). Due to the diversity, preservation and abundance of biota, as well as its paleogeographic and temporal context, the Crato limestones contain one of the most important Early Cretaceous fossil lagerstätten in Gondwana30.
Associated plant taxa
In the Crato Formation flora, the numbers of taxa range from more than 40 based on macrofossils and more than 150 based on palynomorphs of spore and seed-bearing plants, and not all were formally described yet30,31. In addition, the angiosperm component of the Crato Formation flora is diverse both taxonomically and ecologically32, and includes several fossil-taxa with clear affinities with early-diverging extant lineages (ANA grade33,34) or mesangiosperms (e.g., magnoliids35,36,37,38, monocots32,39). Several early eudicot taxa were reported earlier but none had been described yet30.
Angiospermae Lindley (P.D. Cantino & M.J. Donoghue)
Eudicotyledoneae M.J. Donoghue, J.A. Doyle & P.D. Cantino
Order Ranunculales Juss. ex Berchtold & J. Presl
Family cf. Ranunculaceae Juss.
Santaniella Gobo, Coiffard, Bachelier, L. Kunzmann et Iannuzzi gen. nov.
Generic diagnosis
Angiosperm with herbaceous to suffruticose growth habit. Thin elongated axes with leaves ranging from sessile and simple distally to increasingly longer, larger, and more divided and lobed towards the base. Proximal leaves pedately compound with typical bifurcation into two (type II) or more leaflets (type III). Leaf and leaflets unlobed to lobed with untoothed margin. Petiole (and petiolules) with marginal attachment. Primary venation pinnate and secondary craspedodromous, branching dichotomously and merging into a perimarginal vein. Anastomoses rare. Fruit a solitary and terminal follicetum derived from an apocarpous, superior gynoecium with 2 to up to at least 11 multiovulate plicate carpels developing into stipitate, reflexed and pendulous follicles, born on a short, flattened to slightly concave receptacle typically surrounded by persistent perianth-like organs. Fruitlet comprises at least up to 9 seeds arranged in a row along a median longitudinal placenta on the abaxial side.
Etymology
The genus name is derived from the lithostratigraphic unit known as the Santana Group where the studied specimens come from.
Plant Fossil Names Registry Number
PFN002910 (for new genus)
Type species
Santaniella lobata sp. nov.
Remarks
Vegetative and reproductive materials of the two fossil-species within the fossil-genus are preserved either as impressions, permineralizations with iron oxide, and/or coalifications, and beside the designation of the holotypes (Figs. 1, 4), four paratypes (Figs. 2, 3, Supplementary Fig. 1) are selected because they exhibit vegetative and/or reproductive diagnostic characters that are not always clearly visible in the holotype of S. lobata. All three leaf type reconstructions and potentially corresponding fossils are illustrated in Fig. 5.
Santaniella lobata Gobo, Coiffard, Bachelier, L.Kunzmann et Iannuzzi sp. nov.

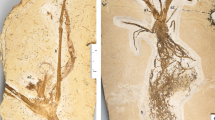
Santaniella lobata gen. nov. et sp. nov. (A–E) holotype (MB. Pb. 1998/691). (A) general view of holotype showing several leaves and a terminal aggregate fruit, with 5–6 stipitate multi-seeded follicles (white arrows point to leaf attachment, gray arrow to a putative opposite leaf, and black arrows to pair of distal most leaves); (B) close-up on follicles attachment to the receptacle, surrounded by 3 outer reflexed (white arrows) and several inner erect and laciniate persistent perianth-like organs (black arrow); (C–E) close-up on leaves showing a gradual acropetal reduction in organization and size from (E) to (C); (C, D) leaf type II. (E) leaf type III with four lobed leaflets (black arrows point to primary veins). Scale bars = 1 cm.

Santaniella lobata gen. nov. et sp. nov. (A–C) paratype (MB. Pb. 1997/1347). (A) general view of paratype showing leaves and a terminal aggregate fruit with 10–11 stipitate follicles (black arrow points to leaf attachment of the distalmost leaf, white arrow to leaf attachment of the basalmost leaf, and gray arrow to a putative sub-opposite leaf); (B) close-up on follicles surrounded by 4 outer (white arrows) and several innermost erect and more or less laciniate (black arrow) persistent perianth-like organs; (C) close-up on leaf type II with two trilobed leaflets; (D) leaf type III (paratype: LP UFC CRT 1559a). Scale bars: 1 cm.

Santaniella lobata gen. nov. et sp. nov. (A–E) paratype (MB. Pb. 1997/1562); (A) fruit with one outer (white arrow) and several inner (black arrow) erect and laciniate persistent perianth-like organs surrounding the stalks of 6 stipitate, reflexed and pendulous follicles; (B) close-up of 3(-4) permineralized seeds; (C–E) SEM images of seeds; (C) whole seed with inserts showing higher magnification of the outer and inner layers of seed coat in the micropylar and chalazal region. Note the jigsaw puzzle-shaped surface cells in chalazal region; (D) close-up of the micropylar region with spiral structures inside the cells of the innermost layer of the seed coat; (E) close-up of the chalazal region with jigsaw puzzle outer layer of the seedcoat. Scale bars: (A, B) = 1 cm; (C) = 500 μm, (C) close-ups = 100 μm; (D) = 100 μm, (D) close-up = 50 μm; (E) and (E) close-up = 100 μm.
Specific diagnosis
Leaves nanophylls and simple to microphylls with at least two to four leaflets. Leaflets usually petiolulate, pinnately lobed and odd-pinnatisect, with acute and decurrent base and obtuse and rounded lobes. Lamina chartaceous. Fruit surrounded by numerous persistent perianth-like organs, typically erect and distally laciniate.
Etymology
The specific epithet refers to the lobed nature of the leaflets.
Holotype
MB.Pb. 1998/691 (repository: Museum für Naturkunde—Leibniz Institute for Evolution and Biodiversity Science, Berlin, Germany), Fig. 1.
Paratypes
MB.Pb. 1997/1347 (Fig. 2A–C), MB.Pb. 1997/1562 (Fig. 3), and MB.Pb 1997/1348 (Supplementary Fig. 1) (repository: Museum für Naturkunde—Leibniz Institute for Evolution and Biodiversity Science, Berlin, Germany); LP UFC CRT 1559a (Fig. 2D) (repository: Federal University of Ceará, Department of Geology, Fortaleza, Brazil).
Plant Fossil Names Registry Number
PFN002911 (for new species)
Type horizon and locality
Open cast pit(s) southwest of Nova Olinda, Ceará, NE Brazil. Lower Cretaceous (uppermost Barremian?/ Aptian), C6 limestone horizon, Crato Formation, Santana Group, Araripe Basin.
Description and remarks
The holotype (MB.Pb. 1998/691) consists of a leafy axis with a terminal fruit (Fig. 1). The axis is about 230 mm long and 2.6 mm wide, and bears several leaves showing a gradual acropetal reduction in both structure and size (Fig. 1A, C–E). The lowest and most proximal leaf is pedately compound and of type III (Fig. 5), with four overlapping lobed and petiolulate leaflets (Fig. 1E), which is better preserved in the second paratype (see below, Fig. 2D). The petiole is 21 mm long and 1.1 mm wide, and the leaf blade is microphyllous, longer and wider than 40 mm. The second and more distal leaf fragments are poorly preserved and can be interpreted as two leaflets of a single leaf of type II, or a pair of leaves of the same type in which the bifurcation is not clearly visible (Figs. 1A,D, 5). The petiole is about 28 mm long and 1.2 mm wide, and the attached leaf fragment is 72 mm long. Individual lobes are 28–35 mm long and 8.5–15 mm wide. The third set of leaf fragments is also poorly preserved and can be interpreted either as a single leaf with two sessile leaflets or a pair of leaves type II (Figs. 1A,C, 5). The petiole is 7.8 mm long and 1.3 mm wide and the attached leaf fragments are 28–31 mm long and 11–13 mm wide. Below the fruit, the distalmost leaves are simple and of type I (Figs. 1A, 5). They are nanophylls, 24 mm long and 2.8 mm wide. L/W ratio is ca. 8:1 and the overall shape is unclear. We do not exclude that they are subtending bracts or a pair of bracteoles.
The aggregate fruit comprises 5–6 stipitate and pendulous follicles. They are 23–30 mm long and 11–12 mm wide, with an 8–8.5 mm long stalk (Fig. 1A). The follicles are connected to a receptacle surrounded by two types of persistent perianth-like organs. The three outermost organs are obovate, with an entire margin and ca. 10 mm long and 6 mm wide (Fig. 1B). Their original organization and number are uncertain but some parts indicate that they were free and reflexed. In contrast, the innermost organs are erect and laciniate (Fig. 1B).
The first paratype (MB.Pb. 1997/1347) is also a leafy axis with a fruit (Fig. 2A). The axis is about 137 mm long and 2.6 mm wide. The most proximal leaf is also pedately compound and of type II (Fig. 5), with two overlapping 3-lobed petiolulate leaflets (Fig. 2C). The petiole is 14 mm long and 1.6 mm wide, and the leaf blade is microphyllous and 36 mm long by ca. 40 mm wide. L/W ratio is about 1:1 and the overall leaf shape is ovate and symmetrical. The leaflets are microphyllous and 36 mm long and 22 mm wide. Their base is acute and decurrent, whereas their lobes are obtuse and rounded. The most distal leaf is simple nanophyll of type I (Figs. 2A, 5) and 18 mm long and 5.2 mm wide. L/W ratio is about 3:1 and the lamina shape is obovate. It is unclear, however, whether there was a second (sub)opposite proximal leaf, and we do not exclude that the distal one is a subtending bract, or one of two bracteoles (Fig. 2A).
The fruit of the first paratype is an aggregate and comprises 10–11 follicles. They are 21–25 mm long and 10–11 mm wide, with a 6–9 mm long stalk (Fig. 2A, B). They are also surrounded at the base by two types of persistent perianth-like organs, comprising 4 outer organs that are very similar to the distalmost leaf, and inner organs with a large base and a more or less laciniate apex (Fig. 2B).
The second paratype (LP UFC CRT 1559a) is a single, well-preserved complete pedately compound leaf of type III (Fig. 5) showing successive dichotomous bifurcations resulting into four lobed asymmetrical leaflets with untoothed margins (Fig. 2D). The complete leaf is reconstructed as being 70 mm long and ca. 50 mm wide. The petiole is about 17 mm long and 1.6 mm wide up to the first bifurcation, and the leaf is a microphyll with a 43 mm long and ca. 50 mm wide blade. L/W ratio is about 1:1 and the lamina shape is ovate and symmetrical. Lamina is chartaceous. The leaflets are sessile and microphyllous and 35–43 mm long and 13–18 mm wide. Their base is acute and decurrent, whereas their lobes are obtuse and rounded. Their blades are also odd-pinnatisect with three to five alternate and asymmetrical lobes, which are obovate and asymmetrical in the middle, and increase in size acropetally. The lobes are proximally decurrent onto the primary vein, but end abruptly distally.
The overall architecture of the leaf (Fig. 5) is derived from a palmate organization with a petiole running straight and then bifurcating. The primary venation of leaflets is pinnate with a single basal vein and without naked basal veins. Agrophic veins and interior secondaries are absent. The major secondaries are craspedodromous and branch dichotomously two to three times before merging into a perimarginal vein. Rare anastomoses occur. The venation of the lobes is mainly composed of secondary veins, except for the distal lobes of each leaflet where their pinnate primary vein ends. Secondary venation is bifurcating and rather thin compared to the primary vein (0.08–0.22 mm), the width ratio secondaries ∕ primary is typically 1:2. The secondaries are regularly spaced (0.17–0.27 mm) and they form uniform moderately acute angles (30–45°) to the primary vein. The attachment of secondaries to the midvein is long decurrent, with the major ones running several millimeters subparallel to the midvein. Intersecondaries and higher order venation are absent. Vein density ranges of 4.5–4.9 (mean of 4.6 mm−1).
The third paratype (MB.Pb. 1997/1562) is a fruit with seeds (Fig. 3). The structure of the fruit is very similar to those of the holotype and first paratype, with 6 stipitate follicles surrounded by several innermost erect laciniate and one outermost obovate (reflexed) persistent perianth-like organs. The follicles are 24–29 mm long and 10–13 mm wide, with a 7–11 mm long stalk (Fig. 3A). The seeds are elliptical and bilaterally symmetrical, and 3–5 mm long and 2 mm wide (Fig. 3A, B). The seed coat comprises two distinct layers, with the outermost composed of elongated cells with straight to undulate walls around the micropyle but forming a jigsaw puzzle-shaped surface over the chalazal region (Fig. 3C–E). A smooth papillate or punctate wall ornamentation with a spongy appearance is also present on the outer layer, especially at the periphery of the seed. In contrast, the inner layer is composed of more or less isodiametric polygonal to irregular cells which form a reticulum, and contain spiral structures which nature remains unclear (Fig. 3D). Below the seed coat, there is no evidence of an embryo, suggesting that it is surrounded by an endosperm and that the seed is albuminous.
The following features were compiled from characters of all specimens of the protologue. The follicles have on average an estimated volume of ca. 1215 mm3 and display numerous transverse veins that are 0.2–1 mm apart. Each follicle contains several seeds (ca. 5–9) with on average an estimated volume of ca. 5.8 mm3, and arranged in a row with the micropyle facing the median longitudinal abaxial placenta.
Santaniella acuta Gobo, Coiffard, Bachelier, L.Kunzmann et Iannuzzi sp. nov.
Specific diagnosis
Leaves nanophylls and simple to microphylls with at least two leaflets. Leaflets usually sessile, with acute and cuneate base and acute and straight to convex apices. Lamina chartaceous. Fruit surrounded by obovate persistent perianth-like organs.
Etymology
The specific epithet refers to the distinctly acute tips of the leaflets.
Holotype
MB.Pb. 2000/71 (Fig. 4) (repository: Museum für Naturkunde—Leibniz Institute for Evolution and Biodiversity Science, Berlin, Germany).

Santaniella acuta gen. nov. et sp. nov. (A, B) holotype (MB. Pb. 2000/71). (A) general view of holotype showing four leaves (white arrow points to leaf attachment of the second proximal leaf of type II and black arrows to distalmost leaves of type I), and a terminal aggregate fruit surrounded by only one persistent perianth-like organ (black triangle). (B) close-up on leaf showing details of venation and apices. Scale bars: 1 cm.
Plant Fossil Names Registry Number
PFN002913 (for new species)
Type horizon and locality
Open cast pit southwest of Nova Olinda, Ceará, NE Brazil. Lower Cretaceous (uppermost Barremian?/ Aptian), C6 limestone horizon, Crato Formation, Santana Group, Araripe Basin.
Description and remarks
The holotype (MB.Pb. 2000/71) and only specimen is a leafy axis with a fruit (Fig. 4). The axis is about 125 mm long and 2.5 mm wide. The two proximal leaves are pedately compound and of type II (Fig. 5). Their petioles are about 6–7 mm long and 1–1.6 mm wide, the leaves are microphylls with ca. 23 mm long and 15 mm wide blade. The L/W ratio is about 1.5:1 and lamina shape is ovate. Lamina is chartaceous. The leaflets are nanophyllous and 16–23 mm long and 5–6 mm wide. They are sessile, lobed to unlobed with untoothed margins, and slightly asymmetrical in the middle of the blade. Their base is acute and cuneate (asymmetrical), and their apices acute and straight to convex. In contrast, the distal-most leaves are simple and of type I (Figs. 4A, 5). One is unlobed and at least 10 mm long and 2.3 mm wide. The L/W ratio is about 4:1. The second distal leaf seems to be bilobed, but it is not well-preserved. However, we do not exclude because of their similarities that the proximal leaves were (sub)opposite and the distal leaves were a pair of bracts or bracteoles.
The overall architecture of the leaf (Fig. 5) is derived from a palmate organization with a petiole running straight and then bifurcating. The primary venation of leaflets is pinnate with a single basal vein and without naked basal veins. Agrophic veins and interior secondaries are absent. The major secondaries are craspedodromous and branch dichotomously two to three times before merging into a perimarginal vein (Fig. 4B). Rare anastomoses occur. Secondary venation is bifurcating and rather thin compared to the primary vein (0.08–0.2 mm), the width ratio between secondaries and the primary vein is typically 1:2 in the distal part of the leaflet. The secondaries are regularly spaced (0.15–0.3 mm) and they form uniform moderately acute angles (30–45°) to the primary vein. The attachment of secondaries to the midvein is long decurrent, with the major ones running several millimeters subparallel to the midvein. Intersecondaries and higher order venation are absent. Vein density is 4.5 mm−1.
The fruit of the holotype is an aggregate and comprises 5–6 follicles surrounded by one obovate, persistent perianth-like organ. The fruitlets are 21–25 mm long and 12 mm wide, with a 7–9 mm long stalk (Fig. 4). They have on average an estimated volume of ca. 1203 mm3 and display numerous transverse veins that are 0.2–0.6 mm apart. The follicles contain at least 5 seeds which are elliptical, bilaterally symmetrical, and 3–5 mm long and 2 mm wide. The seed volumes have on average ca. 5.8 mm3. Their follicles, seeds and perianth-like organ are similar to those of Santaniella lobata. However, there is no evidence of innermost laciniate, persistent perianth-like organs surrounding the follicetum in Santaniella acuta.

Line drawings of leaf type reconstructions of Santaniella gen. nov. and tentative fossil assignments. S. lobata = MB. Pb. 1998/691, MB. Pb. 1997/1347, and LP UFC CRT 1559a; S. acuta = MB. Pb. 2000/71. I, leaf type I: sessile and simple, typically last below the fruit (MB. Pb. 1998/691, MB. Pb. 1997/1347, MB. Pb. 2000/71); II, leaf type II: bifurcate, and can be interpreted as a single leaf with sessile leaflets (MB. Pb. 1998/691), or pedately compound leaves with one bifurcation into two sessile leaflets (MB. Pb. 2000/71). Larger leaves are more clearly pedately compound with distinctly petiolulate and trilobed leaflets (MB. Pb. 1997/1347). Other leaf fragments (MB. Pb. 1998/691) can be interpreted as a single leaf with two leaflets. III, leaf type III: pedately compound leaves with successive bifurcations forming four odd-pinnatisect leaflets (MB. Pb. 1998/691 and LP UFC CRT 1559a). Scale bars = 1 cm.
Cladistic analyses and potential systematics affinities
Our first cladistic analysis (Fig. 6A) was performed using the matrix by Kvaček et al.40 and results in five most parsimonious positions of Santaniella within basal eudicots, either as sister to Glaucideae (Glaucidium), or Hydrastideae (Hydrastis), or Glaucideae + Hydrastideae (successively basal-most lineages of Ranunculaceae), or sister to Nelumbo or Proteaceae + Platanus (basal eudicots). However, if Santaniella is sister to Glaucidium, it would imply changes in distichous phyllotaxis (CH22), marginal teeth (CH35) and carpel number (CH97), and as sister to Hydrastis changes in distichous phyllotaxis (CH22), marginal teeth (CH35) and ovule direction (CH115). Similarly, the Glaucideae + Hydrastideae position implies changes in marginal teeth (CH35), carpel number (CH97) and ovule direction (CH115). If sister to Nelumbo, it would imply changes in leaf dissection (CH34), ovule direction (CH115) and fruit dehiscence (CH126). Also, Nelumbonaceae differ from Santaniella by having large, and centrally peltate leaves, the perianth is caducous and each carpel forms a 1-seeded nutlet embedded in an enlarged and obconic receptacle41. Similarly, the Proteaceae + Platanus position would imply changes in inflorescence structure (CH42), ovule direction (CH115) and fruit dehiscence (CH126). Proteaceae also differ from Santaniella by having a unicarpellate gynoecium42, and Platanaceae by their usually distinct simple, toothed and broadly palmately lobed leaves, and long-pedunculate inflorescence with globular heads, and 1-seeded fruits interpreted as densely hairy achenes or nutlets in a globose head41. Inside Ranunculales, one step less parsimonious positions were recovered as sister to Ranunculaceae as a whole, or as sister to the core of the family. Other ranunculid lineages are two to four steps less parsimonious (see below the second cladistic analysis). One step less parsimonious position outside of the basal eudicots either falls within magnoliids in Piperales and would imply changes in leaf dissection (CH34), inflorescence structure (CH42), carpel number (CH97) and fruit wall (CH124), or within monocots in Dioscoreaceae, but would also imply changes in leaf dissection (CH34), inflorescence structure (CH42), carpel number (CH97) and carpel fusion (CH107). Other positions within other magnoliids or monocots are often less parsimonious and, in magnoliids, fall in lineages which often differ from Santaniella by the absence of several dehiscent follicular fruitlets with many seeds2,43 and, in the case of monocots, can be excluded by their typical linear leaves with parallelodromous or acrodromous to campylodromous venation, and sheathing bases38. Positions within the ANA grade are three to four steps less parsimonious, but in contrast with Santaniella, none of their extant lineages have plicate carpels and/or several ovules, which are synapomorphies of most mesangiosperms44.

Phylogenetic position of Santaniella gen. nov. Most parsimonious positions of Santaniella after its addition to the matrix of Kvaček et al.40 in (A) and Wang et al.45 in (B). Colour scale indicates all most parsimonious positions in red (MP), and those one step less parsimonious in yellow (MP + 1), two steps less parsimonious in green (MP + 2), three steps less parsimonious in cyan (MP + 3), and four steps less parsimonious in dark blue (MP + 4) positions.
Our second cladistic analysis using the dataset of Wang et al.45, which is more centered on Ranunculales, results in the most parsimonious positions of Santaniella within Ranunculales, as sister to Berberidaceae + Ranunculaceae, Berberidaceae, or Ranunculaceae (basal and core lineages), but also among basal eudicots, as sister to Platanus + Proteaceae (Fig. 6B). No changes in any characters are expected in all the most parsimonious positions. However, if Santaniella is within of crown group of Berberidaceae, it would imply changes in carpel number (CH39) and carpel form (CH40), because they fundamentally differ by their single and ascidiate carpel43. The most parsimonious positions within the Ranunculaceae are widespread in basal and core lineages (except Callianthemeae and the last-diverging lineages, i.e., Anemoneae and Ranunculeae). While positions inside Ranunculales (except Ranunculaceae) as sister to Menispermaceaeae or Papaveraceae would be one step less parsimonious, they would imply changes in fruit dehiscence (CH56), or in major venation (CH19), respectively. In addition, Menispermaceae also differ from Santaniella by having simple leaves, and the flowers gathered in inflorescences which develop fruits 1-seeded drupes41, and Papaveraceae by their syncarpous gynoecia with 2, rarely 3 or up to 25 carpels43. Two steps less parsimonious and it could be sister to Circaeasteraceae and Lardizabalaceae, and imply similar changes in major venation (CH19), and fruit dehiscence (CH56). Also, Circaeasteraceae differ from Santaniella by having toothed leaves, achene fruits and 1 to 3 carpels in Circaeaster and 5 to 9 carpels in Kingdonia, but with one ovule in each carpel41,43. In Lardizabalaceae, the flowers are gathered in inflorescences and form fleshy fruits41.
In summary, our cladistic analyses suggest that Santaniella belongs to basal eudicots, most likely to the Ranunculales or the stem lineage of Ranunculaceae. Indeed, the combination of Santaniella features, especially of compound leaves and a superior apocarpous gynoecia with several carpels producing many seeds, is more common in basal eudicots than in any other main lineages of angiosperms46,47. Also, most of traits displayed by Santaniella occur singularly or in various combinations in several extant members of Ranunculales (e.g., in Ranunculaceae), such as the presence of compound and lobed leaves41, craspedodromous secondary venation without intersecondary veins48, bracts (or cauline bract-like leaves) along the stem41, solitary and terminal flowers with short receptacles41, superior apocarpous gynoecium with plicate carpels with transverse veins and several ovules43, aggregate fruits with stalked follicular fruitlets41, persistent floral organs at fruiting stage49, and the range of size and shape of seeds50. Unfortunately, the nature of the persistent perianth-like organs in Santaniella remains unclear, and despite the above similarities the unique combination of its vegetative and reproductive traits does not fit in any extant taxon of Ranunculales.
Comparison to fossil record
Macro- and mesofossil records of putative basal eudicots/ranunculids are widespread in northern mid-latitudes (e.g., China12,13,14, Spain15, Portugal16,17,18, Russia19,20, Kazakhstan21, United States22,23,24). Combined with our findings from southern low-latitudes (Brazil), they shed light on the relatively broad paleogeographical area of this group in the Early Cretaceous.
Some taxa with ranunculidean affinities from Liaoning province, China (late Barremian to early late Aptian age) were reported by Leng and Friis12, Dilcher et al.51, and Sun et al.13. Hyrcantha decussata (Leng & Friis) Dilcher et al. shares with Santaniella terminal fruits with multiple ovules or young seeds, but differs by its syncarpous gynoecium with 2–4 carpels and decussate phyllotaxy of infructescences12,51. Leefructus mirus Sun et al. differs from Santaniella by its simple and trilobate leaves with toothed margins, and syncarpous gynoecium with 5 carpels13. Another eudicot from the Early Cretaceous of China, Gansufructus saligna B. Du., is also preserved at the fruiting stage and bears some resemblances to Santaniella by its persistent perianth, superior gynoecium, and multiple seeds. However, it differs in having simple leaves with poorly organized leaf venation, paniculate infructescences, and tetracarpellate syncarpous gynoecia14.
From the late Barremian of Spain, Iterophyllum lobatum Barral et al., is possibly also a member of Ranunculales and resembles Santaniella lobata by its lobed pinnatipartite leaves with pinnate primary and craspedodromous secondary venation. However, its leaves are always simple with a dentate margin15.
Charcoalified mesofossils with apocarpous gynoecia and superior ovaries containing several ovules with similar affinities to basal eudicots/Ranunculales were also described from the late Barremian to middle Albian of Portugal and the United States. However, all these fossils, e.g., Kajanthus lusitanicus Mendes et al.17, Paisia pantoporata Friis et al.18, and Kenilanthus marylandensis Friis et al.24, are only known from preanthetic or anthetic flowering material which can hardly be compared to the fruits of Santaniella. There is also Teixeira lusitanica von Balthazar et al.16 but it is a unisexual male flower without any evidence of carpels and therefore cannot be compared to Santaniella.
From Aptian to middle Albian Potomac Group sediments in the United States, putative eudicots were described as Potomacapnos apeleutheron Jud & Hickey, which is also characterized by lobed leaflets and veins that ramify dichotomously and fuse into a perimarginal vein like in Santaniella, but differs by its glandular teeth and reticulate venation22. The pinnately lobed leaves of Fairlingtonia thyrsopteroides Jud are also quite similar to those of Santaniella lobata with their pinnate primary veins and a craspedodromous secondary venation branching into perimarginal veins, but they have small petioles, and lobes which decrease in size acropetally and end with a glandular papillate tooth23.
Another potential eudicot, Baderadea pinnatissecta Pessoa et al., was recently described from the Crato Formation flora6. It consists of a single leaf with a pinnatisect dissection similar to that of the leaflets of Santaniella lobata, but the leaf is simple and its lobes are distinctly linear. In addition, its potential affinities with eudicots are only supported by the leaf dissection and primary venation.
Overall importance of our findings
Nowadays, many Ranunculales occur in areas where they could potentially undergo a fossilization process2. However, most of them are typically herbaceous and, in contrast to forest canopy-forming trees or shrubs, are less likely to be preserved as fossils, and also usually produce and shed less vegetative and reproductive parts over their life time52. In addition, they can also occur in environments characterized mostly by erosional rather than depositional processes, and thus, are even less likely to be buried within sediments52.
Fortunately, the depositional lacustrine environment of the Crato Formation along with the microbial mats enables the remarkable preservation potential of various organisms53, including herbaceous and (sub)shrub-like plants less susceptible to be preserved in the fossil record30,54,55. Therefore, the Crato Formation flora allows to track the evolutionary history of plant lineages that are usually not or underrepresented in the fossil record, and is also remarkable by its diversity, especially of early-diverging and basal angiosperm lineages such as Nymphaeales and magnoliids, as well as monocots and eudicots55. It also demonstrates that all major flowering plant lineages were already diversified and lived in low latitudes under hot and (seasonally) dry conditions by the middle Early Cretaceous.
Earlier evidence of eudicots at low paleolatitudes in the Early Cretaceous comes from tricolpate pollen56,57 and is now confirmed by the macrofossils described herein. These new Brazilian fossils are the most complete macrofossils of an early-diverging eudicot with both vegetative and reproductive structures strongly supporting affinities to ranunculids in Early Cretaceous tropics. With all other fossil records linked to Ranunculales from northern mid-latitudes, our results do not only support that the widespread radiation of this clade already started at this early age, but also that they were present in paleoequatorial tropical areas.
Methods
Fossil analysis
The material was studied using a Leica Wild M10 microscope equipped with a Leica DFC 425 camera. Macroscopic images were taken with Canon 250D camera. One specimen exhibits remaining permineralized tissues with cellular structures, especially of in-situ seeds, that were replaced by iron oxide. The complete fossil (MB. Pb. 1997/1562) was directly placed uncoated inside the chamber of a Zeiss EVO 50 Scanning Electron Microscope (SEM) (Carl Zeiss, Germany), and characteristics of the epidermis were observed under a low vacuum. SEM photographs were edited with Photoshop software.
Leaf architecture
The classes of leaf size and part of leaf architecture descriptions follow Ellis et al.58.
Leaf width inferences
The leaf width measurements of leaf types II (Fig. 2C) and III (Fig. 2D) were inferred based on the leaf reconstructions (Fig. 5), assuming that the leaflets were originally not overlapping. They were used to estimate the laminar size, L:W ratio, leaf shape, and medial symmetry of leaf in these specimens.
Vein density measurements
Four lobes of Santaniella fossil-species (S. lobata = LP UFC CRT 1559a; S. acuta = MB. Pb. 2000/71) were used to measure the vein length per area and describe the vein density (mm of vein per mm2), using the program ImageJ (https://imagej.nih.gov/).
Seed and fruitlet volume measurements
The seed and fruitlet volume were estimated based on the formula V = 4/3πab2, where a = L/2, b = (W + T)/4, T = 0.66 W; L = length, W = width and T = thickness, following Eriksson et al.59.
Cladistic analysis
Two cladistic analyses using morphological characters of Santaniella were performed using first the data matrix of Kvaček et al.40 with all major angiosperm lineages except core eudicots, and then using the data matrix of Wang et al.45 which is more specific of Ranunculales. Phylogenetic analyses were conducted using Mesquite 3.6160, where alternative positions were explored and character states were reconstructed. Characters were unordered and equally weighted.
References
Friedman, W. E. The meaning of Darwin’s “abominable mystery”. Am. J. Bot. 96, 5–21 (2009).
Friis, E. M., Crane, P. R. & Pedersen, K. R. Early Flowers and Angiosperm Evolution (Cambridge University Press, 2011).
Kvaček, J. et al. When and why nature gained angiosperms. In Nature Through Time: Virtual Field Trips Through the Nature of the Past (eds Martinetto, E. et al.) 129–158 (Springer, 2020).
Magallón, S., Gómez-Acevedo, S., Sánchez-Reyes, L. L. & Hernández-Hernández, T. A metacalibrated time-tree documents the early rise of flowering plant phylogenetic diversity. New Phyt. 207, 437–453 (2015).
Sun, Y. et al. Phylogenomic and structural analyses of 18 complete plastomes across nearly all families of early-diverging eudicots, including an angiosperm-wide analysis of IR gene content evolution. Mol. Phylo. Evo. 96, 93–101 (2016).
Pessoa, E. M., Ribeiro, A. C. & Jud, N. A. A eudicot leaf from the Lower Cretaceous (Aptian, Araripe Basin) Crato Konservat-Lagerstätte. Am. J. Bot. 108, 2055–2065 (2021).
Friis, E. M., Crane, P. R. & Pedersen, K. R. The Early Cretaceous mesofossil flora of Torres Vedras (NE of Forte da Forca), Portugal: A palaeofloristic analysis of an early angiosperm community. Fossil Imprint 75, 153–257 (2019).
Wang, W., Dilcher, D. L., Sun, G., Wang, H.-S. & Chen, Z.-D. Accelerated evolution of early angiosperms: Evidence from ranunculalean phylogeny by integrating living and fossil data. J. Syst. Evol. 54, 336–341 (2016).
Li, H-T. et al. Origin of angiosperms and the puzzle of the Jurassic gap. Nat. Plants 5, 461–470 (2019).
Sun, Y. et al. Genome sequencing of the endangered Kingdonia uniflora (Circaeasteraceae, Ranunculales) reveals potential mechanisms of evolutionary specialization. iScience 23, 101124 (2020).
Wang, W. et al. The rise of angiosperm-dominated herbaceous floras: Insights from Ranunculaceae. Sci. Rep. 6, 27259 (2016).
Leng, Q. & Friis, E. M. Sinocarpus decussatus gen et sp. Nov., a new angiosperm with basally syncarpous fruits from the Yixian Formation of Northeast China. Plant Syst. Evol. 241, 77–88 (2003).
Sun, G., Dilcher, D. L., Wang, H. & Chen, Z. A eudicot from the Early Cretaceous of China. Nature 471, 625–628 (2011).
Du, B. et al. An exceptionally well-preserved herbaceous eudicot from the Early Cretaceous (late Aptian-early Albian) of Northwest China. Nat. Sci. Rev. 8, nwab084 (2021).
Barral, A., Gomez, B., Feild, T. S., Coiffard, C. & Daviero-Gomez, V. Leaf architecture and ecophysiology of an early basal eudicot from the Early Cretaceous of Spain. Bot. J. Linn. Soc. 173, 594–605 (2013).
von Balthazar, M., Pedersen, K. R. & Friis, E. M. Teixeiraea lusitanica, a new fossil flower from the Early Cretaceous of Portugal with affinities to Ranunculales. Plant Syst. Evol. 255, 55–75 (2005).
Mendes, M. M., Grimm, G. W., Pais, J. & Friis, E. M. Fossil Kajanthus lusitanicus gen et sp. Nov. from Portugal: Floral evidence for Early Cretaceous Lardizabalaceae (Ranunculales, basal eudicot). Grana 53, 283–301 (2014).
Friis, E. M., Mendes, M. M. & Pedersen, K. R. Paisia, an Early Cretaceous eudicot angiosperm flower with pantoporate pollen from Portugal. Grana 57, 1–15 (2018).
Samylina, V. A. Angiosperms from the lower Cretaceous deposits of Kolyma. Bot. Zh. 45, 335–352 (1960).
Krassilov, V. A. & Volynets, Y. Weedy Albian angiosperms. Acta Palaeo 48, 151–169 (2008).
Krassilov, V. A., Shilin, P. V. & Vachrameev, V. A. Cretaceous flowers from Kazakhstan. Rev. Palaeobot. Palynol. 40, 91–113 (1983).
Jud, N. A. & Hickey, L. J. Potomacapnos apeleutheron gen et sp. Nov., a new Early Cretaceous angiosperm from the Potomac Group and its implications for the evolution of eudicot leaf architecture. Am. J. Bot. 100, 2437–2449 (2013).
Jud, N. A. Fossil evidence for a herbaceous diversification of early eudicot angiosperms during the Early Cretaceous. Proc. R. Soc. B. 282, 20151045 (2015).
Friis, E. M., Pedersen, K. R. & Crane, P. R. Kenilanthus, a new eudicot flower with tricolpate pollen from the Early Cretaceous (early-middle Albian) of eastern North America. Grana 56, 161–173 (2017).
Heimhofer, U. & Hochuli, P. A. Early Cretaceous angiosperm pollen from a low latitude succession (Araripe Basin, NE Brazil). Rev. Palaeobot. Palynol. 161, 105126 (2010).
Assine, M. L. et al. Sequências Deposicionais do Andar Alagoas (Aptiano superior) da Bacia do Araripe, Nordeste do Brasil. Bol. Geo. Petro. 22, 3–28 (2014).
Melo, R. M. et al. New marine data and age accuracy of the Romualdo Formation, Araripe Basin, Brazil. Sci. Rep. 10, 15779 (2020).
Lúcio, T., Neto, J. A. S. & Selby, D. Late Barremian/Early Aptian Re–Os age of the Ipubi Formation black shales: Stratigraphic and paleoenvironmental implications for Araripe Basin, northeastern Brazil. J. S. Am. Earth Sci. 102, 102699 (2020).
Coimbra, J. C. & Freire, T. M. Age of the Post-rift Sequence I from the Araripe Basin, Lower Cretaceous, NE Brazil: Implications for spatio-temporal correlation. Rev. Bras. Paleo. 24, 37–46 (2021).
Martill, D. M., Bechly, G. & Loveridge, R. F. The Crato Fossil Beds of Brazil: Window into an Ancient World (Cambridge University Press, 2007).
Kunzmann, L. et al. The Crato flora, a 115 million years old window into the Cretaceous world of Brazil. In Brazilian Paleofloras: From Paleozoic to Holocene (eds Iannuzzi, R. et al.) (Springer, 2022).
Coiffard, C., Kardjiov, N., Manke, I. & Bernardes-de-Oliveira, M. E. C. Fossil evidence of core monocots in the Early Cretaceous. Nature Plants 5, 691–696 (2019).
Mohr, B. A. R., Bernardes-de-Oliveira, M. E. C. & Taylor, D. W. Pluricarpellatia, a nymphaealean angiosperm from the Lower Cretaceous of northern Gondwana (Crato Formation, Brazil). Taxon 57, 1147–1158 (2008).
Coiffard, C., Mohr, B. A. R. & Bernardes-de-Oliveira, M. E. C. Jaguariba wiersemana gen. nov. et sp. Nov., an Early Cretaceous member of crown group Nymphaeales (Nymphaeaceae) from northern Gondwana. Taxon 62, 141–151 (2013).
Mohr, B. A. R. & Eklund, H. Araripia florifera, a magnoliid angiosperm from the Lower Cretaceous Crato Formation (Brazil). Rev. Palaeobot. Palynol. 126, 279–292 (2003).
Mohr, B. A. R. & Bernardes-de-Oliveira, M. E. C. Endressinia brasiliana, a magnolialean angiosperm from the Lower Cretaceous Crato Formation (Brazil). Int. J. Plant Sci. 165, 1121–1133 (2004).
Mohr, B. A. R., Coiffard, C. & Bernardes-de-Oliveira, M. E. C. Schenkeriphyllum glanduliferum, a new magnolialean angiosperm from the Early Cretaceous of northern Gondwana and its relationships to fossil and modern Magnoliales. Rev. Palaeobot. Palynol. 189, 57–72 (2013).
Coiffard, C., Mohr, B. A. & Bernardes-de-Oliveira, M. E. Hexagyne philippiana gen. et sp. Nov., a piperalean angiosperm from the Early Cretaceous of Northern Gondwana (Crato Formation, Brazil). Taxon 63, 1275–1286 (2014).
Coiffard, C., Mohr, B. & Bernardes-de-Oliveira, M. E. C. The Early Cretaceous Aroid, Spixiarum kipea gen. et sp. nov., and implications on early dispersal and ecology of basal monocots. Taxon 62, 997–1008 (2013).
Kvaček, J. et al. Pseudoasterophyllites cretaceus from the Cenomanian (Cretaceous) of the Czech Republic: A possible link between Chloranthaceae and Ceratophyllum. Taxon 65, 1345–1373 (2016).
Kubitzki, K., Rohwer, J. G. & Bittrich, V. The Families and Genera of Vascular Plants. II Flowering Plants Dicotyledons: Magnoliid, Hamamelid and Caryophyllid Families (Springer, 1993).
Weston, P. H. Proteaceae. In The Families and Genera of Vascular Plants IX. Flowering Plants Eudicots: Berberidopsidales, Buxales, Crossosomatales, Fabales p.p., Geraniales, Gunnerales, Myrtales p.p., Proteales, Saxifragales, Vitales, Zygophyllales, Clusiaceae Alliance, Passifloraceae Alliance, Dilleniaceae, Huaceae, Picramniaceae, Sabiaceae (ed. Kubitzki, K.) 364–404 (Springer, 2007).
Endress, P. K. & Igersheim, A. Gynoecium diversity and systematics of the basal eudicots. Bot. J. Linn. Soc. 130, 305–393 (1999).
Endress, P. K. & Doyle, J. A. Reconstructing the ancestral angiosperm flower and its initial specializations. Am. J. Bot. 96, 22–66 (2009).
Wang, W., Lu, A.-M., Ren, Y., Endress, M. E. & Chen, Z.-D. Phylogeny and classification of Ranunculales: Evidence from four molecular loci and morphological data. Plant Ecol. Evol. Syst. 11, 81–110 (2009).
Soltis, D. E., Fishbein, M. & Kuzoff, R. K. Reevaluating the evolution of epigyny: Data from phylogenetics and floral ontogeny. Int. J. Plant Sci. 164, S251–S264 (2003).
Endress, P. K. Evolutionary diversification of the flowers in angiosperms. Am. J. Bot. 98, 370–396 (2011).
Avita, S., Rao, N. V. & Inamdar, J. A. Studies on the leaf architecture of the Ranunculaceae. Flora 171, 280–298 (1981).
Herrera, C. M. Post-floral perianth functionality: Contribution of persistent sepals to seed development in Helleborus foetidus (Ranunculaceae). Amer. J. Bot. 92, 1486–1491 (2005).
Ghimire, B. et al. Seed morphology of the subfamily Helleboroideae (Ranunculaceae) and its systematic implication. Flora 216, 6–25 (2015).
Dilcher, D. L., Sun, G., Ji, Q. & Li, Q. An early infructescence Hyrcantha decussata (comb. Nov.) from the Yixian Formation in northeastern China. PNAS 104, 9370–9374 (2007).
Pigg, K. B. & DeVore, M. L. Paleoactaea gen. nov. (Ranunculaceae) fruits from the Paleogene of North Dakota and the London clay. Am. J. Bot. 92, 1650–1659 (2005).
Varejão, F. G. et al. Exceptional preservation of soft tissues by microbial entombment: Insights into the taphonomy of the Crato Konservat-Lagerstätte. Palaios 34, 331–348 (2019).
Kunzmann, L., Mohr, B. A. R., Wilde, V. & Bernardes-de-Oliveira, M. E. C. A putative gnetalean gymnosperm Cariria orbiculiconiformis gen. nov. et spec. nov. from the Early Cretaceous of northern Gondwana. Rev. Palaeobot. Palynol. 165, 75–95 (2011).
Coiffard, C. & Bernardes-de-Oliveira, M. E. C. Angiosperms in the Early Cretaceous of Northern Gondwana: A track record. In Brazilian Paleofloras: From Paleozoic to Holocene (eds Iannuzzi, R. et al.) (Springer, 2020).
Doyle, J. A., Biens, P., Doerenkamp, A. & Jardiné, S. Angiosperm pollen from the pre-Albian Cretaceous of Equatorial Africa. Bulletin des Centres de Recherches Exploration-Production Elf-Aquitaine 1, 451–473 (1977).
Doyle, J. A. Revised palynological correlations of the lower Potomac Group (USA) and the Cocobeach sequence of Gabon (Barremian-Aptian). Cretac. Res. 13, 337–349 (1992).
Ellis, B. et al. Manual of leaf architecture (Cornell University Press, 2009).
Eriksson, O., Friis, E. M. & Löfgren, P. Seed size, fruit size, and dispersal systems in angiosperms from the Early Cretaceous to the Late Tertiary. Am. Nat. 156, 47–58 (2000).
Maddison, W. P. & Maddison, D. R. Mesquite: A modular system for evolutionary analysis. Version 3.61. http://www.mesquiteproject.org (2019).
Acknowledgements
We are grateful to several staff members of the Museum of Natural History of Berlin (MfN); Cornelia Hiller and Catrin Puffert for their assistance in the curatorial work and Kirsten Born for her support with the SEM. We are also thankful to Dr. Ludwig Luthardt (curator of paleobotanical collection of Museum of Natural History of Berlin) for supporting our work in the collection. We would like to thank Dr. Márcio Mendes for the loan of a specimen of Santaniella from the Laboratório de Paleontologia (UFC, Brazil), and Flávio Lopes from Paleontology and Stratigraphy Department (UFRGS, Brazil) for technical help in the photos and for part of the photography. We also thank Raquel Gewehr de Mello for proofreading an early version of the manuscript. Lastly, we are thankful for the valuable and helpful comments from two anonymous reviewers. This work is part of the PhD thesis of the main author, and was funded by the CAPES-DAAD PROBRAL mobility program (CAPES ID 88881.198776/2018-01; DAAD ID-57446885) and Brazilian National Council for Scientific and Technological Development (Conselho Nacional de Desenvolvimento Científico e Tecnológico—CNPq), as a PhD scholarship for WVG (Process 142135/2018-6), and a fellowship for RI (Processes PQ312747/2017-9). WVG is immensely grateful to the Brazilian and German coordinator of PROBRAL project, Daniel Rodrigues do Nascimento Junior and Lutz Kunzmann, and also to the other project members, Wellington Ferreira da Silva Filho and Márcio Mendes, for all support provided.
Authors statement: The authors are members of a Brazilian-German scientific network initiated for collaborative paleontological-geological investigations in the Brazilian Araripe Basin. In particular, the present paper contains results of the bilateral research project “Towards an integrated analysis of the Early Cretaceous Crato Fossil Lagerstätte” that receives funding support by the CAPES-DAAD PROBRAL Program. The project agenda necessarily implies investigations of specimens in public scientific collections in different countries. We highly acknowledge the friendly invitation from the Museum of Natural History Berlin (Germany) to openly study their collection of Crato plant fossils and to allow publication of the results despite the ongoing and unresolved discussions about the legality of these fossils in German institutional collections. Their support to open science and the doctoral project of the first author are greatly appreciated. Together with the institutional collections, the authors are actively seeking a solution through solid scientific collaboration in the near future that can meet the demands regarding the proper safeguarding of paleontological heritage.
Funding
Open Access funding enabled and organized by Projekt DEAL.
Author information
Authors and Affiliations
Contributions
W.V.G. and C.C. made the design and scope of the research; C.C. conducted phylogenetic analyses; W.V.G., C.C., J.B.B., L.K. and R.I. wrote the manuscript; W.V.G. and C.C. took the photos and with J.B.B., designed the figures.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Gobo, W.V., Kunzmann, L., Iannuzzi, R. et al. First evidence of ranunculids in Early Cretaceous tropics. Sci Rep 12, 5040 (2022). https://doi.org/10.1038/s41598-022-07920-y
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-022-07920-y
This article is cited by
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
