
Abstract
First record of the genus Helius—long-rostrum cranefly from Maestrazgo Basin (eastern Spain, Iberian Penisula) is documented. Two new fossil species of the genus Helius are described from Cretaceous Spanish amber and compared with other species of the genus known from fossil record with particular references to these known from Cretaceous period. Helius turolensis sp. nov. is described from San Just amber (Lower Cretaceous, upper Albian) Maestrazgo Basin, eastern Spain, and Helius hispanicus sp. nov. is described from Álava amber (Lower Cretaceous, upper Albian), Basque-Cantabrian Basin, northern Spain. The specific body morphology of representatives of the genus Helius preserved in Spanish amber was discussed in relation to the environmental conditions of the Maestrazgo Basin and Basque-Cantabrian Basin in Cretaceous.
Similar content being viewed by others

Introduction
The family Limoniidae was previously recorded from Spanish amber. Three species were described: Alavia neli Krzemiński et Arillo, 20071, Helius alavensis Kania, Krzemiński et Arillo, 20162, and Helius spiralensis Kania, Krzemiński et Arillo, 20173, all from Peñacerrada I outcrop1,2,3.
In Cretaceous Spanish amber, we find some of the oldest representatives of the genus Helius, evidence of the beginnings of the evolution of this group of insects. From the Cretaceous period only six species are known, four from Early Cretaceous2,3,4,5, and two from Late Cretaceous6,7. The oldest representatives of the genus were described based on lower Barremian8 inclusions in Lebanese amber, from Hammana-Mdeyrij4,5 and Tannourine (Lebanon4 and two a little younger from upper Albian of Peñacerrada I (Spain)2,3. Two other species were also described from Late Cretaceous, from lower Cenomanian (Tannai village, Myanmar)7 and from Turonian (Orapa Diamond Mine (Botswana)6. While from younger periods 23 species are known, 13 from Eocene9,10,11,12,13, three from Oligocene14,15,16 and seven from Miocene17,18,19,20 (Table 1). In extant fauna the genus Helius is speciose, over 230 extant species of this genus are known and worldwide distributed21. It is interesting and still enigmatic that at the beginning of Cretaceous we find species of the genus Helius with very elongate rostrum, e.g., Late Cretaceous Helius botswanensis Rayner et Waters, 19906 or much older, Early Cretaceous Helius ewa Krzemiński, Kania, Azar, 20145 (the oldest known representative of the genus), suggesting on rapid parallel evolution of this insects and Angiospermae at the beginning of Cretaceous period5, but additional material is needed to support this hypothesis or finally is needed to explain this problem. Herein, two new peculiar species of Helius are described and figured from Spanish amber and characterized by relatively short rostrum.
Material and methods
The study was based on two inclusions in Cretaceous amber of Spain. One of these specimens comes from the Upper Albian amber-bearing deposit of Peñacerrada I (Basque-Cantabrian Basin, near the village of Moraza, Province of Burgos). The second one comes from the upper Albian amber-bearing deposit of San Just (Maestrazgo Basin, Utrillas municipality, Province of Teruel).
The first specimen is deposited at the Museo de Ciencias Naturales de Álava, (Vitoria, Spain) and the second one is housed at the Fundación Conjunto Paleontológico de Teruel-Dinópolis (Teruel, Spain).
The specimens were embedded in epoxy resin (EPO-TEK 301) as described24,25, which allowed physical protection and optimal study in ventral, lateral and dorsal views.
Both specimens were examined with a Nikon (SMZ25) stereomicroscope Nikon SMZ 1500 equipped with a Nikon DS–Fi1 camera. The measurements were taken with NIS–Elements D 3.0 software. The length of head was measured as length of head capsule excluding rostrum. The length of the discal cell—measurements were given from its posterior edge to the point of connection of vein m-m with vein M3. The length of hypopygium was measured from the posterior margin of tergite IX to the tip of gonocoxite. The measurements were given only for undamaged structures. Drawings were completed by tracing the photographs. Drawings (Figs. 1A,B,D and 3B) and photographs were made by Iwona Kania-Kłosok. Map was built using the map Maps-For-Free (https://maps-for-free.com) and modified with the software programs Corel Draw and Corel Photopaint X7. The wing venation and male genitalia nomenclature follows that of17,26,27.

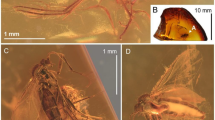
Helius turolensis sp. nov. No. 53–10-54 (male), holotype. (A) head (lateral view), reconstruction; (B) wing; (C) the diagram illustrating the relationship between the length of rostrum (r), antenna (a); palpus (p) and head (h); D. hypopygium (dorsal view), reconstruction; Abbreviation of head (h): a—antenna; fl—flagellomeres; pl—palpus; ped—pedicel; r—rostrum; scp—scape.
Systematic palaeontology
Order Diptera Linnaeus, 175828.
Infraorder Tipulomorpha Rohdendorf, 196129.
Family Limoniidae Speiser, 190930.
Genus Helius Lepeletiere & Serville, 182831.
Type species: Helius longirostris (Meigen, 1818)32.
Key to extinct species of the genus Helius known from Cretaceous
-
1.
Rostrum very elongate reaching almost 0.3 the body length ……………………………………………………………… Helius (Helius) ewa Krzemiński, Kania & Azar, 20145
− Rostrum distincly shorter than 0.3 the body length … 2.
-
2.
Rostrum elongate, but as long as or shorter than head …Helius krzeminskii Ribeiro, 20027.
……………………………………………………………… Rostrum longer than head … 3.
-
3.
Palpus shorter than rostrum … Helius botswanensis Rayner & Waters, 19906.
……………………………………………………………… Palpus as long as or longer than rostrum … 4.
-
4.
Palpus as long as rostrum, last palpomere not very elongate approximately as long as second and third, penultimate palpomere very small and short, about 0.25 the length of the fourth palpomere; antennomeres 1–4 relatively massive … Helius hispanicus sp. nov.
− Palpus longer than rostrum, last palpomere elongate longer second and third; penultimate palpomere elongate, palpomeres 3 elongate, at least 0.3 the length of the fourth palpomere; antennomeres 1–4 elongate, cylindrical ……………………………………………………………… 5.
-
5.
Last palpomere as long as the preceding all taken together …6.
− Last palpomere longer than the preceding all taken together ………………………………………………………………7.
-
6.
Flagellomeres elongate and tiny, slender, last flagellomere distinctly narrow, as long as penultimate one; palpus almost equal in length to rostrum … Helius (Helius) alavensis Kania, Krzemiński & Arillo, 20162.
− Flagellomeres cylindrical, last flagellomere longer than penultimate one; palpus almost two times longer than the length of rostrum Helius lebanensis Kania, Krzemiński & Azar, 20134.
-
7.
Gonocoxite with extra lobe at the apex on dorsal surface, relatively wide and elongated, of comparable size to outer and inner gonostylus, only slightly shorter than outer and inner gonostylus … Helius (Helius) spiralensis Kania, Krzemiński & Arillo, 20163.
− Lacks extra lobe of gonocoxite … Helius turolensis sp. nov.
Helius turolensis sp. nov.

Helius turolensis sp. nov. No. 53–10-54 (male), holotype. (A) body, latero-ventral view; (B) hypopygium; (C) wing.
Diagnosis
Rostrum only slightly longer than head, constitute 0.5 the length of head; antenna relatively slender and short, terminate just beyond the head, each flagellomere with two very elongate setae, approximately 6.7 times longer than length of segments bearing them; palpus longer than rostrum; last palpomere elongate, slender, longer than palpomeres 1–3 combined; third palpomere widened in basal part, about 0.3 × the length of the fourth palpomere.
Etymology
The specific epithet is derived from Teruel, where the specimen was been found.
Material examined
Holotype No. SJ-10-54 (male), housed at the Fundación Conjunto Paleontológico de Teruel-Dinópolis (Teruel, Spain).
Horizon and locality
The specimen was found in amber from gray-black claystones with abundant plant remains which corresponds to a deposit of a fluvial deltaic swamp, in the Utrillas Group33, Lower Cretaceous, upper Albian.
The outcrop of San Just34 is located in the Maestrazgo Basin, municipality of Utrillas (Province of Teruel, Aragón Autonomous Community, eastern Spain).
Description
Body (Fig. 2A) pale brown, 2.97 mm long.
Head (Figs. 1A, 2A): 0.36 mm wide, 0.30 mm high; rostrum 0.38 mm long; antenna (Figs. 1A, 2A) 16-segmented, about 0.67 mm long (Fig. 1C); scape cylindrical and not very elongate, widened in apical part, pedicel short and rather wide, approximately as long as wide; first flagellomere cylindrical, fraction longer than the next one, not extended at the base, three basal segments of antenna with not very elongate a few setae, longer than width and length of segments bearing them, flagellomeres became more slender to the apex of antenna; flagellomeres 2–14 with very long setae, elongate setae 0.51 mm long. Maxillary palp (Figs. 1A, 2A) four segmented, 0.51 mm long (1/0.05 mm; 2/0.07 mm; 3/0.12 mm; 4/0.27 mm) slender, two basal segments almost equal in length, only slightly widened at distal part, third palpomere massive, longer than first and second palpomeres, but shorter than first and second palpomeres combined, widened at basal part and narrowed at distal part, fourth palpomere very elongate, longer than all other palpomeres arranged together, tiny.
Thorax (Fig. 2A): wing (Figs. 1B, 2A,C) 3.65 mm long, 0.93 mm wide; pterostigma present, oval, pale-brown; Sc relatively short, ends well before fork of Rs; Rs relatively short, R2+3+4 almost 1.5 × the length of Rs; R1 ends opposite 0.60 × length of R2+3+4 level; R5 longer than R2+3+4; cross-vein m-cu connected with M3+4 behind half of its length measured from fork of Mb; d-cell, twice longer than wide; M3 slightly waved, 1.5 × as long as d-cell; A1 and A2 slightly waved, arched at the margin of wing, A1 elongate. Tip of A1 behind Rb bifurcation level.
Abdomen: hypopygium (Figs. 1D, 2A,B): 0.66 mm long, with gonocoxite relatively narrow and elongate, approximately 3 × as long as wide; at the apex of gonocoxite bunch of very elongate setae arranged around the tip; outer and inner gonostyles of comparable size; outer gonostylus elongate, slightly widened in the distal part.
Comparison
Helius turolensis sp. nov. differs from all other Cretaceous representatives of genus by the ratio between the length of rostrum, antenna, palpus and head.
This species differs from species known from the Eocene and the Miocene periods by tiny, slender antennae with characteristic, very elongate setae on each flagellomere, 6.7 × longer than length of segments bearing them.
Moreover, in contrast to H. hispanicus sp. nov. third palpomere is massive and widened at the base, in H. turolensis sp. nov. this palpomere is very small, not longer than 1.5 × its width.
The antennae of H. turolensis sp. nov. bears two very elongate, symmetrically arranged setae, similarly to H. hispanicus sp. nov. In contrast to H. hispanicus sp. nov. in H. turolensis sp. nov. palpus is more slender, last palpomere is elongate and tiny, longer than the length of all other palpomeres combined in H. hispanicus sp. nov. last palpomere is not very elongate approximately as long as second and third combined. Differences are also well visible in ratio between the length of rostrum, antenna, palpus and head between these two species.
In H. turolensis sp. nov. palpus is longer than rostrum, antenna is almost 0.25 × longer than palpus, while in H. hispanicus sp. nov. palpus and rostrum are in length, antenna is almost 0.14 × longer than palpus.
In H. alavensis, antennae are slender, without very elongate setae as in H. turolensis. Palpus in H. alavensis is only slightly longer than rostrum, approximately 1.14 × the length of rostrum, in H. turolensis 1.25 × . In contrast to H. spiralensis, gonocoxite in H. turolensis lacks extra lobe, in H. spiralensis the extra lobe is present and bears elongate, strong setae at apex.
In contrast to other Cretaceous species H. turolensis differs especially by morphology of the head, the wing venation and the male genitalia.
In H. ewa, rostrum is very elongate, reaching about the third of body length, in H. turolensis rostrum is only slightly longer than head, head ca. 0.36 mm long; rostrum ca. 0.38 mm long. In H. lebanensis, discal cell is opened, in contrast to H. turolensis where discall cell is closed. In H. krzeminskii, rostrum is relatively short and the differences between these two species are well visible comparing the ratio of the length of rostrum, the length of head and head appendages. In H. botswanensis palpus, is shorter than rostrum, rostrum is approximately twice as long as head, while in H. turolensis, palpus is longer than rostrum, rostrum is approximately as long as the head.
Helius hispanicus sp. nov.

Helius hispanicus sp. nov. No. MCNA 9946 (sex undefinied), holotype. (A) the diagram illustrating the relationship between the length of rostrum (r), antenna (a); palpus (p) and head (h); (B) head. Abbreviation of head (h): as in Fig. 2.

Helius hispanicus sp. nov. No. MCNA 9946 (sex undefinied), holotype. (A) fore part of the body (lateral view); (B) last palpomeres; (C) head (lateral view); Abbreviation of head (h): as in Fig. 2.
Diagnosis
Rostrum only slightly longer than head; antennomeres 1–4 relatively massive and short, terminate just beyond the head; each flagellomere with two very elongate setae, approximately 4.5 × longer than segments bearing them; palpus as long as rostrum; last palpomere not very elongate approximately as long as second and third; penultimate palpomere very small and short, about 0.25 × the length of the fourth palpomere.
Etymology
The specific epithet is derived from Hispania, Latin name for Spain.
Material examined
Holotype No. MCNA 9946 (sex unidentified), Peñacerrada, Álava, Spain, housed at the Museo de Ciencias Naturales de Álava, Vitoria, Spain.
Horizon and locality
The type specimen was found in amber from coal levels with abundant plant remains developed in delta plain areas which corresponds to the top of filling sequences of interdistributary bays but it is also found in filling deposits of abandoned fluvial channels or crevasse splay, in the Utrillas Group33, Lower Cretaceous, upper Albian.
The outcrop of Peñacerrada I34 is located in the Basque-Cantabrian Basin, municipality of Moraza (Province of Burgos, Castilla y León Autonomous Community, northern Spain).
Description
The head and thorax (Figs. 3B, 4A) brown.
Head (Figs. 3, 4A,C): 0.30 mm wide, 0.33 mm high; rostrum 0.59 mm long; antenna (Figs. 3B, 4A,C) 16-segmented, about 0.69 mm long (Fig. 3A) (1/0.43 mm; 2/0.22 mm; 16/0.10 mm); scape cylindrical, but wide and massive, elongate, 2.5 × longer than wide, with a few not very elongate setae; pedicel relatively elongate and wide, massive, widened in distal part, longer than wide; first flagellomere wide, relatively short, flagellomeres 1–14 tapering toward apex; each flagellomere with two symmetrically arranged very long setae, much longer than segments bearing them, last flagellomere with two very elongate setae arranged at the tip since the last it’s the one at the tip. Maxillary palp (Figs. 3B, 4A–C) four-segmented, two basal segments massive, relatively elongate and almost equal in length (length of palpomeres 0.59 mm: 1/0.14; 2/0.14 mm; 3/0.07 mm; 4/0.24 mm), third palpomere small, 1.5 × longer than wide, about half the length of first and second palpomeres; last palpomere not very elongate, shorter than all other palpomeres combined, only 4 × as long as wide.
Remarks
The specimen is poorly preserved. Abdomen not preserved, wings and legs are only partly preserved. But, the relation between the length of rostrum, antennae and palpus, and other morphological features like morphology of antenna or palpus allow to classify this specimen to the genus Helius and to describe it as a new species.
Comparison
Helius hispanicus sp. nov. differs from all other representatives of genus by the by the occurrence of very small and short third palpomere, which constitute only 0.25 × the last one. Third palpomere is very small, not longer than 1.5 × of its width, while in all other extinct species known from Cretaceous period this segment of maxillary palpus is rather elongate and widened. Moreover, Helius hispanicus sp. nov. differs from Cretaceous representatives of genus by ratio between the length of rostrum, antenna, palpus and head. In H. spiralensis and H. alavensis, palpus is longer than rostrum, in H. hispanicus is as long as rostrum, in H. botswanensis is distinctly shorter than rostrum. Rostrum as long as or shorter than head occur in H. krzeminskii, in H. (H.) ewa rostrum is very elongate reaching almost 0.3 the body length, while in H. hispanicus rostrum is longer than head, but and shorter than in H. (H.) ewa.
On the antennae of this species occur two very elongate, symmetrically arranged setae, these very elongate setae does not occur in other species of Helius known from fossil record of younger periods like Eocene.
Discussion
Six species of flies belonging to the genus Helius are known from the cretaceous period and only three are known from Spanish amber1,2,3 (Fig. 5). All the species of the genus Helius described so far on the basis of inclusions in Cretaceous Spanish amber came from the same outcrop Peñacerrada I, Álava, Basque-Cantabrian Basin, Spain.

Maps with enlarged view of the distribution of know Cretaceous localities of the representatives of the genus Helius. Map was built using the map Maps-For-Free (https://maps-for-free.com) and modified with the software programs Corel Draw and Corel Photopaint X7.
One of the species new to science included in this work, H. hispanicus sp. nov., was described on the basis of inclusions also originating from the outcrop Peñacerrada I, Álava, while the other fossil representative H. turolensis sp. nov. was found at the outcrop of Sun Just located in the Maestrazgo Basin. Both the first and second positions are dated to upper Albian. It should be noted that this is the first record of the genus Helius from fossil record of the Maestrazgo Basin.
Most of the 150 Iberian Penisula amber deposits are dated to Albian (Early Cretaceous), only a few of these localities are dated to Late Cretaceous e.g. in Asturias or Catalonia, and only two localities with amber are dated to the late Triassic (both in Alicante)35.
Both Basque-Cantabrian Basin and Maestrazgo Basin were formed during significant changes in fauna and flora on earth. The large sedimentary Basque-Cantabrian Basin has developed between the Iberian and European tectonic plates, the Maestrazgo basin is an intercontinental basin located in the Iberian Range35. In the Basque-Cantabrian Basin amber is preserved in rocks rich in organic matter from sediments, is large quantity of plants remains, coal and other continental organic material which was transported by rivers. Largest deposits of Maestrazgo Basin is San Just with abundant plant remains and fusinished wood, were probably formed during subtropical hot-humid environment with hot and dry areas35.
It is in such an area with a subtropical, hot and humid environment and dry areas that the representatives of the genus Helius occurred in Cretaceous period.
Recent species of the genus Helius occur mainly in Oriental, Australian and Oceanian or Neotropic regions21 and many species of this genus prefer rather hot climates.
Based on geological and biotic evidence, a sharp distinction between the northern territories of Laurasia and the southern landmasses of Gondwana had happened in the earliest late Mesozoic36. According to Eurogondwana model, which explains the migration of Eurogondwanan lineages to Laurasia36, European region were probably connected to Africa during the Early Cretaceous through the Apulian microplate and was served 130 Ma37,38. The research conduct that e.g. the hormurid scorpions colonized Laurasia from Africa via the Apulia microplate (Europa terrane) in the Cretaceous. The migrations of some taxa into Asia and North America were possible through Europe, on the other hand, Europe was preventing Laurasia taxa from migrating to Africa38.
Most species of the genus Helius, whose bodies have been preserved in Spanish amber, have certain common features. Antennae and palpi are usually thin, very narrow, slender, the flagellomeres are small and the boundaries between them are often difficult to distinguish. The antennae of species known from Spanish amber are also relatively short and among the species of the genus Helius we do not find representatives with a very elongate rostrum, the rostrum is usually only slightly longer than the head, as in Helius alavensis or Helius spiralensis. These set of characters distinguish the group of species known from Spanish amber from other known fossil species. A comparable long rostrum also occurs in a bizarre stem scydmaenine beetle from the mid-Cretaceous Burmese amber39, which represents a case of convergent evolution in Cretaceous insects.
These insects rapidly evolved during the Cretaceous period and adapted to the new food spectrum offered by angiosperms (flowering plants) that becoming abundant at that time2,3,4,5,7,40,41, but as it is suggested42, some of them could drops of extinct gymnosperms to feed on floral nectar and pollination.
References
Krzemiński, W. & Arillo, A. Alavia neli, n. gen. and n. sp.: The first Limoniidae (Diptera) from the Lower Cretaceous amber of Álava (Spain). Alavesia 1, 11–13 (2007).
Kania, I., Krzemiński, W. & Arillo, A. First representative of the genus Helius Lepeletier and Serville, 1828 (Diptera, Limoniidae) from the Lower Cretaceous Álava amber (Spain). Cretac. Res. 63, 33–38 (2016).
Kania, I., Krzemiński, W. & Arillo, A. A new peculiar species of the genus Helius Lepeletier & Serville, 1828 (Diptera, Limoniidae) from Cretaceous Álava amber (Spain). Earth Environ. Sci. Trans. R. Soc. Edinb. 107, 1–7 (2017).
Kania, I., Krzemiński, W. & Azar, D. The oldest representative of Helius Lepeletier & Serville 1828 (Limoniidae, Diptera) from Lebanese amber (Early Cretaceous). Insect Syst. Evol. 44, 1–8 (2013).
Krzemiński, W., Kania, I. & Azar, D. The Early Cretaceous evidence of rapid evolution of the genus Helius Lepeletier and Serville, 1828 (Limoniidae, Diptera). Cretac. Res. 48, 96–101 (2014).
Rayner, R. J. & Waters, S. B. A Cretaceous crane-fly (Diptera: Tipulidae): 93 million years of stasis. Zool. J. Linn. Soc. 99, 309–318 (1990).
Ribeiro, G. C. A new fossil Helius (Diptera, Limoniidae) from Burmese amber. Stud. Dipterol. 9(2), 403–408 (2002).
Maksoud, S. & Azar, D. Lebanese amber: Latest updates. Palaeoentomology 3(2), 125–155. https://doi.org/10.11646/palaeoentomology.3.2.2 (2020).
Loew, H. Über den Bernstein Und die Bernsteinfauna. Program der Keiserischen Realschule Meseritz, 1–44 (1850).
Krzemiński, W. Fossil Tipulomorpha (Diptera, Nematocera) from Baltic amber (Upper Eocene): Revision of the genus Helius Lepeletier et Serville (Limoniidae). Acta Zool. Cracov. 35, 597–601 (1993).
Podenas, S. New species of Helius crane flies (Diptera: Limoniidae) from Baltic amber (Eocene). Mitteil. Geol. 86, 229–238 (2002).
Kania, I. Subfamily Limoniinae Speiser, 1909 (Diptera, Limoniidae) from Baltic amber (Eocene): The genus Helius Lepeletier & Serville, 1828. Zootaxa 3814(3), 333–352 (2014).
Krzemiński, W. et al. True flies (Insecta: Diptera) from the late Eocene insect limestone (Bembridge Marls) of the Isle of Wight, England, UK. Earth Environ. Sci. Trans. R. Soc. Edinb. 110(3–4), 495–554 (2019).
Statz, G. Neue Tipulidenfunde aus den Braunkohlen-Schiefern von Rott am Siebengebirge. Wissensch. Mitteil. Verein Nat. Heimatkunde Köln 1(3), 90–106 (1934).
Statz, G. New Dipteren (Nematocera) aus dem Oberoligocän von Rott. III. Familie Limnobiidae (Stelzmücken). IV. Familie: Tipulidae (Schnaken). V. Familie: Culicidae (Stechmücken). Palaeontographica 95, 93–120 (1944).
Krzemiński, W. A first fossil Helius (Diptera, Limoniidae) from North America. Acta Zool. Cracov. 34, 311–313 (1991).
Krzemiński, W. Three new species of the genus Helius Lepeletier & Serville (Diptera, Limoniidae) from the Middle Miocene of Stavropol (northern Caucasus, Russia). Acta Zool. Cracov. 45(4), 317–320 (2002).
Podenas, S. & Poinar, G. O. Jr. New short-palped crane flies (Diptera-Limoniidae) from Mexican amber. Proc. Entomol. Soc. Wash. 114, 347–371 (2012).
Wu, S., Krzemiński, W., Soszyńska-Maj, A. & Ren, D. New fossil representative of the Helius (Diptera, Limoniidae) from the little known and newly discovered locality Caergen Village of northeastern Tibetan Plateau (China). Palaeontol. Electron. 22, 1–8 (2019).
Kopeć, K., Kania, I. & Krzemiński, W. New and little known crane-fly species of the genera Helius, Elephantomyia and Toxorhina (Diptera, Limoniidae) from Dominican and Mexican amber. Palaeontol. Electron. 19, 1–14 (2016).
Oosterbroek, P. Catalogue of the Crane-flies of the World. (Diptera, Tipuloidea: Pediciidae, Limoniidae, Cylindrotomidae, Tipulidae). http://nlbif.eti.uva.nl/ccw/index.php. Last updated 28 December 2012. Accessed 1 February 2021 (2021).
Kania, I., Wojtoń, M., Kopeć, K., Owsiak, A. & Jordan, W. Helius anetae sp. nov. (Diptera: Limoniidae), a new representative of the genus from the Eocene Baltic amber. Geol. Paläontol. 281, 101–109 (2016).
Cockerell, T. D. A. British fossil insects. Proc. U.S. Natl. Mus. 49, 469–499 (1915).
Corral, J.C., López del Valle, R. & Alonso, J. El ámbar cretácico de Álava (Cuenca Vasco-Cantábrica, norte de España). Su colecta y preparación. Estudios del Museo de Ciencias Naturales de Álava 14 (Special Publication, number 2), 7–21 (1999).
Nascimbene, P. & Silverstein, H. The preparation of fragile Cretaceous ambers for conservation and study of organismal inclusions. In Studies on Fossils in Amber, with Particular Reference to the Cretaceous of New Jersey (ed. Grimaldi, D.) 93–102 (Backhuys Publishers, 2000).
McAlpine, J. F. B., Peterson, B., Shewell, G., Teskey H., Vockeroth, J. & Wood, D. Manual of Nearctic Diptera. Vol. 1. Agriculture Canada Research Branch Monograph, 27 (1981).
Ribeiro, G. C. Phylogeny of the Limnophilinae (Limoniidae) and early evolution of the Tipulomorpha (Diptera). Invertebr. Syst. 22, 627–694 (2008).
Linnaeus, C. Systema nature per regna tria naturae, secundum classes, ordines, genera, species, cum caracteribus, differentiis, synonymi, locis. Tomus I. Editio decima, reformata. L. Salvii, Holmiae [= Stockholm], 824 (1758).
Rohdendorf, B. B. The oldest infraorders of Diptera from the Triassic of Middle Asia. Palaeontol. Zhurnal 2, 90–100 (1961) (In Russian).
Speiser, P. 4 Orthoptera. Orthoptera Nematocera. Wissenschaftliche Ergebnisse der Schwedischen Zoologische Expededition nach Kilimandjaro, Meru 10 (Diptera), 31–65 (1909).
Le Peletier, A. L. M. & Serville, J. G. A. Entomologie, ou histoire naturelle des crustacés, des arachnides et des insectes. Encycloped. Method. Hist. Nat. 10, 345–833 (1828).
Meigen, J. W. Systematische Beschreibung der bekannten europäischen zweiflügeligen Insecten. Erster Theil. F.W. Forstmann, Aachen. xxxvi + 332 + [1] p. (1818).
Barrón, E. et al. Palynology of Aptian and upper Albian (Lower Cretaceous) amber-bearing outcrops of the southern margin of the Basque-Cantabrian Basin (northern Spain). Cretaceous Res. 52, 292–312 (2015).
Peñalver, E. & Delclòs, X. Spanish Amber. In Biodiversity of Fossils in Amber from the Major World Deposits (ed. Penney, D.) 236–270 (Siri Sci. Press, 2010).
Murillo-Barroso, M. et al. Amber in prehistoric Iberia: New data and a review. PLoS ONE 1, 1–36 (2018).
Ezcurra, M. & Agnolín, F. L. A new global paleobiogeographical model for the late Mesozoic and early Tertary. Syst. Biol. 61(4), 553–566 (2012).
Bosellini, A. Dinosaurs, “re-write” the geodynamics of the eastern Mediterranean and the paleogeography of the Apulia Platform. Earth Sci. Rev. 59, 211–234 (2002).
Monod, L. & Prendini, L. Evidence for Eurogondwana: The roles of dispersal, extinction and vicariance in the evolution and biogeography of Indo-Pacific Hormuridae (Scorpiones: Scorpionoidea). Cladistics 31, 71–111 (2015).
Yin, Z., Cai, C. & Newton, A. F. Beetle with long ‘nose’: A bizarre stem scydmaenine in amber from Myanmar (Coleoptera: Staphylinidae: Scydmaeninae). Cretac. Res. 89, 98–106 (2018).
Silvestro, D. et al. Fossil data support a pre-Cretaceous origin of flowering plants. Ecol. Evol. https://doi.org/10.1038/s41559-020-01387-8 (2021).
Li, H. et al. Origin of angiosperms and the puzzle of the Jurassic gap. Nat. Plants 5, 461–470 (2019).
Khramov, A. V., Bashkuev, A. S. & Lukashevich, E. D. The fossil record of long-proboscid nectarivorous insects. Entomol. Rev. 100(7), 881–968 (2020).
Acknowledgements
We would like to thanks Dr. Wiktoria Jordan-Stasiło (University of Rzeszów) for making the maps of the distribution of fossil sites of the representatives of the genus Helius. This project was supported the project CRE: Global bioevent of massive resin production at the initial diversification of modern forest ecosystems, funded by the Spanish AEI/FEDER, UE Grant CGL2017-84419 and the project by the National Science Centre, Poland, Grant No. 2016/23/B/NZ8/00936.
Funding
Open Access funding enabled and organized by the University of Rzeszów.
Author information
Authors and Affiliations
Contributions
I.K. conceived and designed the study, lead and performed the data analysis, interpretations and writing, making photographs, drawings, graphical figures, analysis, writing and corrections of the manuscript; correspondence. A.A. corrections of the manuscript. W.K. writing and corrections of the manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Kania-Kłosok, I., Krzemiński, W. & Arillo, A. Two new long-rostrum cranefly species from the Cretaceous Iberian amber (Diptera, Limoniidae, Helius). Sci Rep 11, 12851 (2021). https://doi.org/10.1038/s41598-021-91803-1
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-021-91803-1
This article is cited by
-
Cretaceous Antodicranomyia (Diptera: Limoniidae) and their paleohabitat
Scientific Reports (2022)
-
New finding of Toxorhina (Ceratocheilus) limoniid fly in Eocene Baltic amber and the biogeographical context of the genus
Scientific Reports (2022)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
