
Abstract
Disorders of consciousness (DOC) are one of the major consequences after anoxic or traumatic brain injury. So far, several studies have described the regaining of consciousness in DOC patients using deep brain stimulation (DBS). However, these studies often lack detailed data on the structural and functional cerebral changes after such treatment. The aim of this study was to conduct a volumetric analysis of specific cortical and subcortical structures to determine the impact of DBS after functional recovery of DOC patients. Five DOC patients underwent unilateral DBS electrode implantation into the centromedian parafascicular complex of the thalamic intralaminar nuclei. Consciousness recovery was confirmed using the Rappaport Disability Rating and the Coma/Near Coma scale. Brain MRI volumetric measurements were done prior to the procedure, then approximately a year after, and finally 7 years after the implementation of the electrode. The volumetric analysis included changes in regional cortical volumes and thickness, as well as in subcortical structures. Limbic cortices (parahippocampal and cingulate gyrus) and paralimbic cortices (insula) regions showed a significant volume increase and presented a trend of regional cortical thickness increase 1 and 7 years after DBS. The volumes of related subcortical structures, namely the caudate, the hippocampus as well as the amygdala, were significantly increased 1 and 7 years after DBS, while the putamen and nucleus accumbens presented with volume increase. Volume increase after DBS could be a result of direct DBS effects, or a result of functional recovery. Our findings are in accordance with the results of very few human studies connecting DBS and brain volume increase. Which mechanisms are behind the observed brain changes and whether structural changes are caused by consciousness recovery or DBS in patients with DOC is still a matter of debate.
Similar content being viewed by others

Introduction
Human consciousness is often described as a complex phenomenon, consisting of two components, wakefulness and awareness1,2. While wakefulness is associated with functional brainstem neurons i.e. the reticular system projecting to both thalamic and cortical neurons, awareness is mostly related to the functional integrity of the thalamus, cerebral cortex, and their connections1,2,3,4.
Disorders of consciousness (DOC) occur as a result of interference with the mentioned systems. DOC can be acute and reversible, as a transient stage in a spontaneous recovery due to trauma, or chronic and irreversible, as in patients with unresponsive wakefulness syndrome (UWS)1,2,3,4. Prolonged DOC is not only long-lasting, serious, and currently untreatable, but also has severe consequences on a patient’s quality of life5,6,7. These conditions impose a serious consideration from both medical and ethical perspectives7,8,9. Clinical conditions such as anoxic or traumatic brain lesions can cause diffuse neuronal damage resulting in serious disorders ranging from minimally conscious state (MCS), UWS, coma, locked-in syndrome, and even brain death1,10. The boundaries separating the mentioned conditions are unclear, with each one including severities1,2. UWS is currently described as wakefulness without consciousness, including a complete lack of reactions. Additionally, it is characterized by variable cycles of sleeping and wakefulness, with preserved spontaneous respiration, digestion, and thermoregulation1,2,10,11,12,13. MCS is on the other hand characterized by inconsistent, but visible evidence of consciousness1,14 with the patient being awake but unaware. A patient in MCS can follow instructions occasionally, give simple gestural or non-verbal yes/no responses, and show a certain level of purposeful movement. The incidence of UWS is roughly 5 to 25 per million, while the prevalence in the adult population in the USA ranges from 40 to 168 per million15. Additionally, various neurological impairments occur, including seizures, movement disorders, myoclonus, focal motor, and sensorimotor deficits, as well as emotional, behavioral, and cognitive disturbances16,17.
Despite advances in diagnosis, improved classification, use of modern technology, and well-known pharmacological treatments alongside non-invasive brain stimulation, the surgical therapeutic approaches are still not significantly advanced18,19. Several studies used deep brain stimulation (DBS) of certain nuclei, such as centromedian parafascicle (CM-pf) complex and brain stem reticular formation, to regain consciousness in UWS and MCS patients5,6,19,20,21,22,23. Research thus often focused on the thalamus as a functionally important thalamocortical link, where stimulation through key thalamocortical and thalamus-forebrain circuitry was shown to be important and caused widespread effect24. The use of the CM-pf complex as a target was based on literature overview and clinical recommendation, as well as due to the connectivity and prospective effect on systems involved in consciousness5,6,19,20,21,22,23,25. Previous studies were done on a small cohort, mainly as case reports, and did not yield a definite answer whether this highly specific and refined method can be therapeutically successful5,6,19,20,21,22,23,26,27,28,29,30. Even though some studies claimed that there was no definitive evidence of the efficacy of this method for treatment of DOC patients, the usefulness of DBS was shown using controlled studies, as well as our previous study6,22,23.
Virtually all of the studies mentioned are missing an explanation of the detailed structural and functional cerebral changes induced by DBS. Recently, renewed interest in the grading of DOC and different trials of therapeutic approaches have been noted. Previous studies observed short-term volume changes in the human brain structure following DBS, potentially indicating restorative possibilities in patients with neurodegenerative diseases31. Modern magnetic resonance imaging (MRI) techniques play an important role in diagnostic classification and evaluation of cerebral damage in order to improve the gradation of different types and levels of DOC32. Moreover, MRI has been shown to be a potential tool for morphometric measurements and changes in a restructured, “reorganized” brain in DOC patients33,34. Modern neuroimaging thus uses different methods to objectively evaluate both structural and functional changes as an effect of different treatments.
Previously, it has been documented that DBS changes function in such structures as the striatum, hippocampus, amygdala, etc. It would thus arguably follow that if electrical stimulation changes the brain’s structure and its function, stimulation of damaged circuitry could then induce both functional and structural changes with an eventual improvement of function35,36,37.
To determine the impact of CM-pf stimulation on a possible brain structural reorganization after functional recovery, we performed a quantitative volumetric MRI analysis in DOC patients who underwent CM-pf DBS. Our study aimed to reveal if CM-pf DBS could induce an impact on brain structural reorganization in DOC patients and lead to a changed volume of specific cortical and subcortical structures.
Methods
Patients
This retrospective study included five patients who underwent CM-pf DBS due to DOC after which they achieved functional recovery (Table 1). Out of the five patients, three were female, average age 16 ± 1.53. The cause of injury in one patient was anoxic due to cardiac arrest, while other two patients experienced traumatic injuries. Two patients were male with an average age of 20 ± 4.24 years. In both of these patients the cause of injury was anoxia due to cardiac arrest. The length between the injury and time to DBS was 2 months in males and 11 ± 5.13 months (range 4–14) in females. The average follow-up duration in female patients was 88 ± 38.17 months (range 30–102), while in male patients it was 84 ± 38.18 months (range 57–111).
In the present study we included patients previously described in Chudy et al.22,23. Forty-nine patients were enrolled in the previous study. Fourteen patients (10 UWS and 4 MCS) fulfilled neurophysiologic, clinical, and neuroimaging criteria and underwent DBS implantation. Four patients (3 MCS and 1 UWS) achieved functional recovery, while other patients were further followed by an attending palliative physician22,23. In this study we included one additional UWS patient who underwent DBS implantation after fulfilling the aforementioned criteria and achieved functional recovery.
Patients selected for DBS were evaluated utilizing the standard Rappaport Disability Rating (RDR) scale and the Coma/Near Coma (C/NC) scale22,23,38. Inconsistently responsive to simple commands patients were classified as MCS patients (C/NC Level 1), while patients who were not able to respond to any command were classified as UWS (C/NC Level 2–4). DOC patients were selected based on three main factors: their neurophysiologic evaluation, 12/24 h electroencephalography (EEG) and neuroimaging (MRI). The neurophysiological criteria we investigated were recordable somatosensory evoked potential (SEP), motor evoked potentials, and brainstem auditory evoked potentials, even with pathological parameters such as prolonged latencies or central conduction time. The entry criterion for SEPs was recordability via stimulation of median nerves, with or without SEPs elicited by tibial nerve stimulation. The second main factor we looked at was EEG. The entry criterion for this method was the presence of periods of desynchronized EEG activity during 12/24 h of monitoring processed EEG22,23. Obtained neuroimaging (MRI) showed absence of a structural defect. After passing extensive selection criteria, the patients underwent DBS electrode implantation into the CM-pf complex of the left thalamic intralaminar nuclei, while in patients with posttraumatic lesions, the electrode was placed in the better-preserved hemisphere22,23. On the third postoperative day, monopolar stimulation was initiated using the contact eliciting the strongest arousal response with minimal current (25 Hz frequency, 90 µs pulse duration, voltage 2.5–3.5 V). Stimulation was applied for a 30-min period every two hours during the daytime. Stimulation duration was up to 10 months after DBS electrode implantation. The informed parental/caregivers consent was obtained for all patients in accordance with the Declaration of Helsinki and ethical approval was obtained from the Institutional Review Board of the Dubrava University Hospital, School of Medicine, University of Zagreb, Croatia.
MRI acquisition
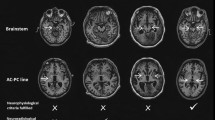
MRI scans were obtained on a 1.5 T MRI scanner (MAGNETOM Aera, Siemens Healthineers, Erlangen) using 24 channeled head coils. Standard clinical sequences were used, as well as high-resolution 3D T1-weighted magnetization-prepared rapid acquisition gradient echo (MPRAGE) sequence with the following scanning parameters: TR = 2300 ms, TE = 3 ms, flip angle = 15°, matrix size = 256 × 256, field of view = 256 mm, voxel size = 1.0 × 1.0 × 1.0 mm. Obtained MRI scans were closely examined showing the absence of any large brain lesion (Fig. 1). MRI was obtained three times; the first one prior to DBS implantation, the second one between 8 and 12 months after surgery (approximately 1 year), and the third time between 1 and 7 years after DBS implementation.

Initial structural MRI scans of included patients showing an absence of any large brain lesion (patients 1–5, from left to right).
Quantitative volumetric methods
Volumetric analysis was conducted using an automated CIVET processing pipeline (version 2.1.0., http://www.bic.mni.mcgill.ca/ServicesSoftware/CIVET) of different inter-dependent algorithms, on a web-based portal, CBRAIN (https://portal.cbrain.mcgill.ca), providing tools for observer-independent corticometric analysis39,40,41. During analysis, the pipeline included the following steps: correction of radiofrequency intensity and nonuniformity artifacts corrected using the N3 algorithm42, linear registration using the 9-parameter affine process of translation, scaling, and rotation, and the 3D T1 volume registration to the ICBM 152 stereotactic space43,44. The brain was masked, including scull stripping and tissue classification as grey matter, cerebrospinal fluid, and white matter, using the discrete classifier with advanced neural network methods45,46. Cortical, grey matter and white matter surfaces extraction using the Laplacian map, partial volume classification47,48 and thickness calculation49 were final steps. The surfaces were smoothed and registered in order to calculate the regional cortical volume and the average regional thickness of the cerebral cortex of automatically parcellated lobes50,51,52. Quantification of regional cortical thickness averages where lobe borders were determined by sulcal landmarks and detected by the CIVET pipeline and regional cortical volume estimates were made for parietal, occipital, frontal, temporal lobes, parahippocampal gyrus, cingulate gyrus, isthmus of the cingulate gyrus, and insula (Fig. 2a). Subcortical structure segmentation (putamen, caudate, thalamus, globus pallidus, hippocampus, amygdala, nucleus accumbens) was performed using an automated MRI brain volumetry system, volBrain53 (Fig. 2b). Volumetric analysis was performed at three measuring points, according to MRI scans: prior to DBS, approximately 1 year after, and 7 years after DBS implementation.

Volumetric analysis was conducted using the CIVET pipeline measuring regional cortical volume and thickness for parietal, occipital, frontal, temporal lobes, isthmus of the cingulate gyrus, parahippocampal, and cingulate gyrus and insula (a), and volBrain software, for subcortical structures segmentation (b). Single-subject volumetric analysis is presented.
Statistical analysis
Data analysis was performed using the MedCalc Statistical Software version 12.5.0 (MedCalc Software, Ostend, Belgium; https://www.medcalc.org). Data were plotted as individual values, horizontal lines and markers represent mean ± SD. Comparison of the regional cortical volumes and thicknesses of parcellated lobes and volumes of subcortical structures were obtained. Depending on the distribution and type of variables, volumetric data in all measuring points were analyzed using one-way ANOVA, with the Student–Newman–Keuls post hoc test, or Kruskal–Wallis test with pairwise comparisons according to Conover54. In order to reduce intersubject variability we have performed the regional volumes normalization by head size. Normalization was done using the subject with lowest head size value as a calibrator value to calculate the coefficient (subject value/calibrated value). Regional volumes were multiplied with the calculated coefficient for each subject and further analyzed using previously mentioned statistical test. We did not correct for multiple comparisons in the present small patient sample. The statistical significance was set at p < 0.05.
Ethics approval and consent to participate
This study was carried out in accordance with the recommendations of the Ethical Board of the Dubrava University Hospital and University of Zagreb, School of Medicine with written informed parents/caregivers consent from all subjects in accordance with the Declaration of Helsinki. The protocol was approved by the Institutional Review Board of the University Hospital Dubrava and University of Zagreb, School of Medicine.
Results
Analysis of segregated volumes of the lobes of the cerebral cortex in three measuring points revealed statistically significant volume increase in right parahippocampal gyrus (↑25.53%) (F(2, 9) = 6.658, p = 0.01), left cingulate gyrus (↑16.65%) (F(2, 9) = 6.679, p = 0.01) and left insula (↑22.74%) (F(2,9) = 6.202, p = 0.04). However, volume increase was observed for left parahippocampal gyrus volume (↑17.63%) (F(2,9) = 1.142, p = 0.36), right cingulate gyrus volume (↑15.92%) (F(2,9) = 1.311, p = 0.31) and right insula volume (↑7.69%) (F(2,9) = 0.871, p = 0.45) (Fig. 3). Volumetric analysis of frontal (L ↓7.57%, R ↑3.22%), parietal (L ↑0.05%, R ↓2.54%), occipital (L ↓9.99%, R ↓15.08%,), and temporal (L ↓3.38%, R ↑2.14%) lobe, as well as isthmus of the cingulate gyrus (L ↑1.53%, R ↑1.52%), bilaterally, did not revealed statistically significant results.

Regional cortical volumetric analysis in three measuring points (prior to DBS, 1 year after DBS, and 7 years after DBS). The regional cortical volumetric analysis revealed significant volume increase in right parahippocampal gyrus, left cingulate gyrus, and left insula, while the trend of volume increase was presented in left parahippocampal gyrus volume, right cingulate gyrus volume, and right insula volume. Vertical bars, standard deviation.
Subcortical structures volumetric analysis revealed statistically significant volume increase in caudate (↑17.7%) (F(2,9) = 4.964, p = 0.04), hippocampus (↑9.07%) (F(2,9) = 4.052, p = 0.02) and amygdala(↑39.53%) (F(2,9) = 4.163, p = 0.02). While putamen (↑10.22%) (F(2,9) = 1.576, p = 0.58) and accumbens (↑28.57%) (F(2,9) = 1.915, p = 0.21) volumes were observed increased, both globus pallidus (↓4.49%) (F(2,9) = 0.054, p = 0.95) and thalamus (↓24.16%) (F(2,9) = 1.104, p = 0.37) volume revealed volume decrease over time, especially prominent in thalamus (Fig. 4). While right parahippocampal gyrus regional cortical thickness increase significantly (↑16.9%) (F(2,9) = 1.937, p = 0.02), regional cortical thickness increase was observed in both right cingulate gyrus and insula bilaterally, as well as in the left parahippocampal gyrus, although these values were not statistically significant. Regional cortical thickness is observed to decrease over time in the parietal, frontal, temporal, and occipital lobe, as well as the isthmus of the cingulate gyrus.

Subcortical structures volumetric analysis in three measuring points (prior to DBS, 1 year after DBS, and 7 years after DBS). Significant volume increase of caudate, hippocampus, and amygdala were presented, while putamen and accumbens volumes presented a trend of volume increase during three measuring points. Both globus pallidus and thalamus volume decreased over time. Vertical bars, standard deviation.
Statistical analysis of normalized segregated volumes revealed significantly higher volume of the left insula (F(2,9) = 6.804, p = 0.01). Volumes of the right insula (F(2,9) = 1.407, p = 0.29), the left parahippocampal gyrus (F(2,9) = 1.142, p = 0.36), the right parahippocampal gyrus (F(2,9) = 1.694, p = 0.23), the left cingulate gyrus (F(2,9) = 3.064, p = 0.09) and the right cingulate gyrus (F(2,9) = 2.423, p = 0.14), as well as volumetric analysis of frontal, parietal, occipital, and temporal lobe, and isthmus of the cingulate gyrus, did not significantly differ among groups. Subcortical normalized volumes of the amygdala (F(2,9) = 4.451, p = 0.04) were significantly higher. Normalized volumes of the caudate (F(2,9) = 2.952, p = 0.22), hippocampus (F(2,9) = 3.390, p = 0.08), putamen (F(2,9) = 1.015, p = 0.40), accumbens (F(2,9) = 2.436, p = 0.14), globus pallidus (F(2,9) = 0.032, p = 0.96) and thalamus (F(2,9) = 0.747, p = 0.50) did not significantly differ.
All five patients included in the study raised to full awareness and regained the ability to communicate and interact. Two male patients (Patient 1 and 2) were graded as MCS using RDR and C/NC scale (Table 1). Both of them are currently able to live independently to a large extent22,23, although both of them are experiencing problems with concentration and memory (Table 2). Out of the three female patients, one was graded as MCS (Patient 3), while two of them were graded as UWS (Patient 4 and 5) using RDR and C/NC scale (Table 1). All female patients regained consciousness, although they are currently still unable to live independently22,23. The main obstacle for further progress is related to motor difficulties, such as spasticity, contractures, inability to move their limbs, to articulate words, etc. For several years, their communication was in a form of non-verbal signs, predominantly as facial gestures and/or limb movement. Today, these patients are able to form simple words (Table 2).
Discussion
Previous studies reported that DBS in DOC is still in its exploratory phase5,6,19,20,21,22,23,26,27,28,29,30. Keeping in mind the ethical criteria and seriousness of DOC, our previous results showed limited but encouraging success22,23. Although brain atrophy was initially observed on MRI prior to DBS, it was difficult and challenging to explain the volume increase of several brain structures and areas after DBS, which was in accordance with previous studies31. Two main aspects are going to be discussed in the following paragraphs. Firstly, a brief overview of consciousness, the role of the thalamus in conscious networks, as well as the general functional effects of stimulation. Secondly, we will discuss the structural changes that occurred after functional improvement, as a result of processes of repair, plasticity, and reorganization.
General effects of deep brain stimulation
All major factors and phenomena of consciousness are not fully understood at the moment. Despite numerous different study explanations and well-elaborated hypotheses, the neurobiological mechanism of consciousness is still largely unexplored37,55,56,57,58. What is known however is that various components of the central nervous systems participate simultaneously in this intriguing phenomenon56,57,58,59,60,61. Alongside the reticular ascending systems projecting from the upper reticular formation of the brainstem, other structures participate significantly in forming consciousness. These structures include monoaminergic pathways, the locus coeruleus, serotonergic and dopaminergic systems, the forebrain cholinergic system, as well as the nucleus basalis62,63,64,65,66. In addition, a special system of neurons connecting the brainstem to the interstitial neurons may also participate67 and form important networks for maintaining and regulating cortical functions68,69. These neurons project towards a number of subcortical structures, in which projections from thalamus nuclei are also received62,63,64,65,66. We propose that the main functional system responsible for the observed structural changes is widespread thalamocortical and thalamosubcortical (feedback) circuitry, modulated by several systems: modulatory monoaminergic systems, the reticular ascending system from the upper brainstem, basal forebrain cholinergic systems, etc.
The thalamus is a crucial relay of the cortico-striatal-thalamo-cortical circuits and its role in arousal, attention to salient stimuli, and processing of information has been documented by both animal models as well as functional connectivity studies in humans24,25,70,71,72,73,74. The CM-pf complex as the main part of the intralaminar nuclei24 provides particularly strong connections to several subcortical structures74. It also provides strong connections to the basal ganglia75, primarily the caudate nucleus76, putamen24, nucleus accumbens77, and pallidum, all of which was demonstrated using both tracing and tractographic studies78. A relatively weaker subcortical connectivity was noticed between CM-pf and the amygdala79, and the hippocampus80,81. On the other hand, excitatory projections from the CM-pf complex toward cortical areas have been reported as much weaker and non-specific24. The cortical projections seem to be limited to the agranular cortex with the only exception being the anterior insular/frontal operculum78. Additionally, the CM-pf complex probably interacts with various cortical areas via the corticostriatal pathways, through strong retrograde projections36.
The selection of the thalamus as a stimulation target thus seems justified because of its potent effect in the thalamocortical circuitry. Our preliminary results showed the thalamic volume decrease less than 1% at the second measuring point, while at the third measuring point thalamic volume decrease of almost 25% was observed (Fig. 4). Since the thalamic volume was slightly decreased at the period when functional recovery was observed in patients, we presume that the constant volume contributed to functional connectivity of the thalamus and previously mentioned structures. Results of positive stimulation effects point to the thalamus and reorganization of functional circuitry as important factors in recovery after DOC. The DBS electrode, positioned deep inside the brain, provides permanent stimulation. This in turn provides new possibilities for integrating function and structure for consciousness recovery. However, despite the evidence that thalamic intralaminar nuclei have an influence on cerebral function and connectivity, it is unlikely that it can explain such major changes in the brain structure volume. In our patients, a larger volume decrease was observed several years after the functional recovery, thus we can only assume it occurred as a further adaptation of the thalamus to the brain reorganization due to the initial brain injury, either trauma or anoxic lesion. Additionally, the thalamus volume could decrease in response to the lack of adequate stimulus. Once the DBS is removed, the stimulus that maintained thalamic volume by providing sufficient stimulation may cease. The variability of structural changes observed after DBS may be partly explained by the great variety of cortical, subcortical, and brainstem functional systems involved. This variability suggests that there is no single functional system affected. Therefore, besides the thalamus and indirectly affected systems, other factors play a possible role and will be discussed below.
Structural effects observed on MRI
The exact underlying mechanisms contributing to volume changes are still unknown. There are several possible explanations about how long functional stimulation could have led to a volume increase. These include synaptogenesis, gliogenesis, axonal remodeling, micro vascularization, neuronal size increase, and extracellular matrix changes. A combination of all these factors is also possible. Of note is that adult neurogenesis has also been described in certain brain regions (i.e. the hippocampus)31,82. It is well known that after various lesions, the brain volume could be changed by different underlying processes of brain tissue repair (apoptosis, microstructural glial reactivity, changes in the extracellular matrix, etc.), plasticity (sprouting, myelination, dendritic plasticity), and reorganization83,84,85,86,87. Therefore, we could expect similar events occurring and leading to the increased brain volume in our patients. Additionally, processes of repair and plasticity probably overlap. The reorganization of pathways is thus expected to be primarily functional (since various pathways transduce electrical information when the brain is stimulated), rather than structural, which is possible primarily during development86,87. Nevertheless, considering how the most voluminous components of the brain tissue are cell bodies of neurons, glia, dendrites, and myelinated axons88,89,90, it is unlikely that these cells experience a significant increase in their size receiving more excitatory input. Moreover, an increase of dendritic branching postsynaptic spines of super excited neurons is possible. The data on glia are less consistent, but it is generally expected that astroglia follows a metabolic increase of its function and become hypertrophic when stimulated91. Volume increase could therefore occur during an increase in synapsis number and size which are more activate during prolonged stimulation of important excitatory glutamatergic neurons92. However, synapsis makes only a small proportion of the brain volume90.
Several examples of increased brain volume during development in abnormal conditions have been previously reported. In such cases, hyper-connectivity within the cortex was explained by the changes in the extracellular substance, which makes approximately 70% of the developing brain, while being scarce in the normal adult brain93,94,95.
The aforementioned structural changes (repair, plasticity, and reorganization) are impossible to follow in patients, even if postmortem material is available. Therefore, MRI is so far the only available method providing general information that something is structurally different. Additionally, it is especially useful to combine different MRI techniques such as tractography with easily performed volumetric measurements, a valuable indicator that structural processes are taking place in the brain after stimulation. Only a few volumetric MRI analyses in patients with DOC were performed so-far33,34. Grey matter volume of the parahippocampal gyrus, thalamus, and caudate were previously presented to be the key features differing healthy subjects and patients with DOC. Additionally, white matter volumes of the parahippocampal gyrus, isthmus of cingulate gyrus, and brainstem were previously described as the most affected white matter regions34. Analysis of the subcortical structures was previously performed only for the thalamus; slower atrophy of the thalamus was observed in MCS than in UWS patients33. These results suggest that patients with better-preserved levels of consciousness are more likely to preserve an increased brain volume over time. Our results indicate that the most informative region is the parahippocampal area. This area is known to be important for the formation of episodic memories as well as permitting rapid processing which in turn enables contextualization of outside events. It is therefore well located to bind associations from other cortical streams, suggesting an important function in consciousness96. Our findings are in the accordance with the results of previous preclinical studies of DBS in rodents which observed structural neuroplasticity (hippocampal neurogenesis97,98), as well as an increased complexity of apical dendrites and the length of basal dendritic trees of pyramidal neurons of the hippocampus99. Furthermore, our findings are in accordance with results showing short-term brain volume increase, namely the hippocampal area, in Alzheimer’s dementia patients following DBS31.
Several limitations of the presented study should be mentioned, such as a small number of patients, their mixed gender, different age and etiology of injury as well as different times for MRI scans from the initial injury and volumetric analysis in included patients. Still, we believe the presented results are valuable and can be a valid starting point for future research.
Conclusions
Even on a small sample size included in the present study, we tried to emphasize the importance of morphometric MRI quantification which can expand our understanding of the relationship between brain structures and consciousness recovery. Our results are in accordance with the results of very few human studies connecting DBS and brain volume changes. They also support the idea that DBS in the CM-pf complex may have a widespread effect on cerebral function and structure.
Structures with altered volumes belong to cortico-thalamo-cortico-basal ganglia circuitry, limbic circuitry and at the same time have strong modulatory monoaminergic brainstem input. The nature of the initial lesion of neural networks, neurons, and glia, is difficult to determine with the current criteria and methodology applied. Which mechanisms are behind the observed brain changes and whether the observed structural changes are caused by consciousness recovery and/or DBS in DOC patients is yet to be elucidated. In future studies, different MRI volumetric analysis of cortical areas or deep brain structures could help to identify cell types and more details contributing to the regional volume change.
Data availability
The datasets generated for this study are available on request to the corresponding author.
References
Plum, F. & Posner, J. B. The Diagnosis of Stupor and Coma 3rd edn. (F A Davis Co, Philadelphia, 1980).
Bernat, J. L. Chronic disorders of consciousness. Lancet 367(9517), 1181–1192. https://doi.org/10.1016/S0140-6736(06)68508-5 (2006).
Schnakers, C. & Laureys, S. Coma and Disorders of Consciousness (Springer, Switzerland, 2017). https://doi.org/10.1007/978-3-319-55964-3.
Zeman, A. Consciousness. Brain 124(Pt 7), 1263–1289. https://doi.org/10.1093/brain/124.7.1263 (2001).
Yamamoto, T. et al. DBS therapy for the vegetative state and minimally conscious state. Acta Neurochir. Suppl. 93, 101–104. https://doi.org/10.1007/3-211-27577-0_17 (2005).
Yamamoto, T. et al. Deep brain stimulation for the treatment of vegetative state. Eur. J. Neurosci. 32, 1145–1151. https://doi.org/10.1111/j.1460-9568.2010.07412.x (2010).
Vanhoecke, J. & Hariz, M. Deep brain stimulation for disorders of consciousness: Systematic review of cases and ethics. Brain Stimul. 10(6), 1013–1023. https://doi.org/10.1016/j.brs.2017.08.006 (2017).
Schiff, N. D., Giacino, J. T. & Fins, J. J. Deep brain stimulation, neuroethics, and the minimally conscious state: Moving beyond proof of principle. Arch. Neurol. 66(6), 697–702. https://doi.org/10.1001/archneurol.2009.79 (2009).
Johnson, L. S. The right to die in the minimally conscious state. J. Med. Ethics 37(3), 175–178. https://doi.org/10.1136/jme.2010.038877 (2011).
Laureys, S. et al. Unresponsive wakefulness syndrome: A new name for the vegetative state or apallic syndrome. BMC Med. 1(8), 68. https://doi.org/10.1186/1741-7015-8-68 (2010).
Calvet, J. & Coll, J. Meningitis of sinusoid origin with the form of coma vigil. Rev. Otoneuroophtalmol. 31, 443–445 (1959).
Kretschmer, E. Das apallische Syndrom. Z. Ges. Neurol. Psychiat. 169, 292–296 (1940).
Jennett, B. & Plum, F. Persistent vegetative state after brain damage. A syndrome in search of a name. Lancet 1(7753), 734–737. https://doi.org/10.1016/s0140-6736(72)90242-5 (1972).
Giacino, J. T. et al. The minimally conscious state: Definition and diagnostic criteria. Neurology 58(3), 349–353. https://doi.org/10.1212/wnl.58.3.349 (2002).
Beaumont, J. G. & Kenealy, P. M. Incidence and prevalence of the vegetative and minimally conscious states. Neuropsychol. Rehabil. 15(3–4), 184–189. https://doi.org/10.1080/09602010443000489 (2005).
Anderson, C. A. & Arciniegas, D. B. Cognitive sequelae of hypoxic-ischemic brain injury: A review. NeuroRehabilitation 26(1), 47–63. https://doi.org/10.3233/NRE-2010-0535 (2010).
Lu-Emerson, C. & Khot, S. Neurological sequelae of hypoxic-ischemic brain injury. NeuroRehabilitation 26(1), 35–45. https://doi.org/10.3233/NRE-2010-0534 (2010).
Thibaut, A., Schiff, N., Giacino, J., Laureys, S. & Gosseries, O. Therapeutic interventions in patients with prolonged disorders of consciousness. Lancet Neurol. 18(6), 600–614. https://doi.org/10.1016/S1474-4422(19)30031-6 (2019).
Tsubokawa, T. et al. Deep-brain stimulation in a persistent vegetative state: Follow-up results and criteria for selection of candidates. Brain Inj. 4(4), 315–327. https://doi.org/10.3109/02699059009026185 (1990).
Cohadon, F. & Richer, E. Deep cerebral stimulation in patients with post-traumatic vegetative state 25 cases. Neurochirurgie 39(5), 281–292 (1993).
Yamamoto, T. et al. Deep brain stimulation and spinal cord stimulation for vegetative state and minimally conscious state. World Neurosurg. 80(3–4), S30.e1-9. https://doi.org/10.1016/j.wneu.2012.04.010 (2013).
Chudy, D. et al. Deep brain stimulation for the early treatment of the minimally conscious state and vegetative state: Experience in 14 patients. J. Neurosurg. 128(4), 1189–1198. https://doi.org/10.3171/2016.10.JNS161071 (2018).
Chudy, D., Raguž, M. & Deletis, V. Deep brain stimulation for treatment patients in vegetative state and minimally conscious state. In Neurophysiology in Neurosurgery: A Modern Approach 2nd edn (eds Deletis, V. et al.) 515–521 (Academic Press, New York, 2020).
Van der Werf, Y. D., Witter, M. P. & Groenewegen, H. J. The intralaminar and midline nuclei of the thalamus. Anatomical and functional evidence for participation in processes of arousal and awareness. Brain Res. Brain Res. Rev. 39(2–3), 107–140. https://doi.org/10.1016/s0165-0173(02)00181-9 (2002).
Metzger, C. D., van der Werf, Y. D. & Walter, M. Functional mapping of thalamic nuclei and their integration into cortico-striatal-thalamo-cortical loops via ultra-high resolution imaging-from animal anatomy to in vivo imaging in humans. Front. Neurosci. 8(7), 24. https://doi.org/10.3389/fnins.2013.00024 (2013).
Hassler, R., Ore, G. D., Dieckmann, G., Bricolo, A. & Dolce, G. Behavioural and EEG arousal induced by stimulation of unspecific projection systems in a patient with post-traumatic apallic syndrome. Electroencephalogr. Clin. Neurophysiol. 27(3), 306–310. https://doi.org/10.1016/0013-4694(69)90060-1 (1969).
McLardy, T., Mark, V., Scoville, W. & Sweet, W. Pathology in diffuse projection system preventing brainstem-electrode arousal from traumatic coma. Confin. Neurol. 31(4), 219–225. https://doi.org/10.1159/000103482 (1969).
Sturm, V., Kühner, A., Schmitt, H. P., Assmus, H. & Stock, G. Chronic electrical stimulation of the thalamic unspecific activating system in a patient with coma due to midbrain and upper brain stem infarction. Acta Neurochir. (Wien) 47(3–4), 235–244. https://doi.org/10.1007/bf01406406 (1979).
Schiff, N. D. et al. Behavioural improvements with thalamic stimulation after severe traumatic brain injury. Nature 448(7153), 600–603. https://doi.org/10.1038/nature06041 (2007).
Magrassi, L. et al. Results of a prospective study (CATS) on the effects of thalamic stimulation in minimally conscious and vegetative state patients. J. Neurosurg. 125(4), 972–981. https://doi.org/10.3171/2015.7.JNS15700 (2016).
Sankar, T. et al. Deep brain stimulation influences brain structure in Alzheimer’s disease. Brain Stimul. 8(3), 645–654. https://doi.org/10.1016/j.brs.2014.11.020 (2015).
Gosseries, O., Di, H., Laureys, S. & Boly, M. Measuring consciousness in severely damaged brains. Annu. Rev. Neurosci. 37, 457–478. https://doi.org/10.1146/annurev-neuro-062012-170339 (2014).
Rubeaux, M., Jayashri Mahalingam, J., Gomez, F., Nelson, M., & Vanhaudenhuyse, A. et al. Thalamic volume as a biomarker for disorders of consciousness. I Proceedings Volume 9287, 10th International Symposium on Medical Information Processing and Analysis. 2015;9287:92870R. https://doi.org/10.1117/12.2071146.
Annen, J. et al. Regional brain volumetry and brain function in severely brain-injured patients. Ann. Neurol. 83(4), 842–853. https://doi.org/10.1002/ana.25214 (2018).
Dandy, W. E. The location of the conscious center in the brain: The corpus striatum. Bull. Johns Hopkins Hosp. 79, 34–58 (1946).
Halgren, E., Walter, R. D., Cherlow, D. G. & Crandall, P. H. Mental phenomena evoked by electrical stimulation of the human hippocampal formation and amygdala. Brain 101(1), 83–117. https://doi.org/10.1093/brain/101.1.83 (1978).
Crick, F., Koch, C., Kreiman, G. & Fried, I. Consciousness and neurosurgery. Neurosurgery 55(2), 273–281. https://doi.org/10.1227/01.neu.0000129279.26534.76 (2004).
Rappaport, M. The disability rating and coma/near-coma scales in evaluating severe head injury. Neuropsychol. Rehabil. 15(3–4), 442–453. https://doi.org/10.1080/09602010443000335 (2005).
Ad-Dab’bagh, Y., Einarson, D., & Lyttelton, O., et al. The CIVET image-processing environment: A fully automated comprehensive pipeline for anatomical neuroimaging research. In: Proceedings of the 12th Annual Meeting of the Organization for Human Brain Mapping (Corbetta, M. eds). Florence: NeuroImage; 2006.
Sherif, T. et al. CBRAIN: A web-based, distributed computing platform for collaborative neuroimaging research. Front. Neuroinform. 21(8), 54. https://doi.org/10.3389/fninf.2014.00054 (2014).
Bregant, T. et al. Region-specific reduction in brain volume in young adults with perinatal hypoxic-ischaemic encephalopathy. Eur. J. Paediatr. Neurol. 17(6), 608–614. https://doi.org/10.1016/j.ejpn.2013.05.005 (2013).
Sled, J. G., Zijdenbos, A. P. & Evans, A. C. A nonparametric method for automatic correction of intensity nonuniformity in MRI data. IEEE Trans. Med. Imaging 17(1), 87–97. https://doi.org/10.1109/42.668698 (1998).
Collin, D. L., Neelin, P., Peters, T. M. & Evans, A. C. Automatic 3D intersubject registration of MR volumetric data in standardized talairach space. J. Comput. Assist. Tomogr. 18(2), 192–205. https://doi.org/10.1097/00004728-199403000-00005 (1994).
Mazziotta, J. et al. A probabilistic atlas and reference system for the human brain: International Consortium for Brain Mapping (ICBM). Philos. Trans. R. Soc. Lond. B Biol. Sci. 356(1412), 1293–1322. https://doi.org/10.1098/rstb.2001.0915 (2001).
Smith, S. M. Fast robust automated brain extraction. Hum. Brain Mapp. 17(3), 143–155. https://doi.org/10.1002/hbm.10062- (2002).
Zijdenbos A, Forghani R, Evans A. Automatic quantification of MS lesions in 3D MRI brain data sets: Validation of INSECT. In Medical Image Computing and Computer-Assisted Intervention (MICCAI98). Lecture Notes in Computer Science (Wells, W. M., Colchester, A., & Delp, S. eds.) Boston (1998).
Kim, J. S. et al. Automated 3-D extraction and evaluation of the inner and outer cortical surfaces using a Laplacian map and partial volume effect classification. Neuroimage 27(1), 210–221. https://doi.org/10.1016/j.neuroimage.2005.03.036 (2005).
MacDonald, D., Kabani, N., Avis, D. & Evans, A. C. Automated 3-D extraction of inner and outer surfaces of cerebral cortex from MRI. Neuroimage 12(3), 340–356. https://doi.org/10.1006/nimg.1999.0534 (2000).
Ad-Dab’bagh Y, Singh V, Robbins S, Lerch P, Lyttelton O, Fombonne E. et al. Native space cortical thickness measurement and the absence of correlation to cerebral volume. In Proceedings of the 11th Annual Meeting of the Organization for Human Brain Mapping (Zilles, K., eds). Toronto: NeuroImage (2005).
Robbins, S. M. Anatomical standardization of the human brain in Euclidean 3-D space and on the cortical 2-Manifold. Ph.D. thesis. School of Computer Science. Montreal, McGill University (2004).
Lyttelton, O., Boucher, M., Robbins, S. & Evans, A. An unbiased iterative group registration template for cortical surface analysis. Neuroimage 34(4), 1535–1544. https://doi.org/10.1016/j.neuroimage.2006.10.041 (2007).
Boucher, M., Whitesides, S. & Evans, A. Depth potential function for folding pattern representation, registration and analysis. Med. Image Anal. 13(2), 203–214. https://doi.org/10.1016/j.media.2008.09.001 (2009).
Manjón, J. V. & Coupé, P. volBrain: An online MRI brain volumetry system. Front. Neuroinform. 27(10), 30. https://doi.org/10.3389/fninf.2016.00030 (2016).
Conover, W. J. Practical Nonparametric Statistics (Wiley, New York, 1999).
Crick, F. & Koch, C. A framework for consciousness. Nat. Neurosci. 6(2), 119–126. https://doi.org/10.1038/nn0203-119 (2003).
Mogensen, J. & Overgaard, M. Reorganization of the connectivity between elementary functions—a model relating conscious states to neural connections. Front Psychol. 20(8), 625. https://doi.org/10.3389/fpsyg.2017.00625 (2017).
Overgaard, M. & Mogensen, J. Visual perception from the perspective of a representational, non-reductionistic, level-dependent account of perception and conscious awareness. Philos. Trans. R. Soc. Lond. B Biol. Sci. 369(1641), 20130209. https://doi.org/10.1098/rstb.2013.0209 (2014).
Overgaard, M. The status and future of consciousness research. Front. Psychol. 10(8), 1719. https://doi.org/10.3389/fpsyg.2017.01719 (2017).
Delacour, J. Neurobiology of consciousness: An overview. Behav. Brain Res. 85(2), 127–141. https://doi.org/10.1016/s0166-4328(96)00161-1 (1997).
Block, N. Consciousness, accessibility, and the mesh between psychology and neuroscience. Behav. Brain Sci. 30(5–6), 481–499. https://doi.org/10.1017/S0140525X07002786 (2007).
Bayne, T., Hohwy, J. & Owen, A. M. Are there levels of consciousness?. Trends Cogn. Sci. 20(6), 405–413. https://doi.org/10.1016/j.tics.2016.03.009 (2016).
Moruzzi, G. & Magoun, H. W. Brain stem reticular formation and activation of the EEG. Electroencephalogr. Clin. Neurophysiol. 1(4), 455–473. https://doi.org/10.1016/0013-4694(49)90219-9 (1949).
Ramón-Moliner, E. & Nauta, W. J. The isodendritic core of the brain stem. J. Comput. Neurol. 126(3), 311–335. https://doi.org/10.1002/cne.901260301 (1966).
Young, G. B. & Pigott, S. E. Neurobiological basis of consciousness. Arch. Neurol. 56(2), 153–157. https://doi.org/10.1001/archneur.56.2.153 (1999).
Parvizi, J. & Damasio, A. R. Neuroanatomical correlates of brainstem coma. Brain 126(Pt 7), 1524–1536. https://doi.org/10.1093/brain/awg166 (2003).
Samuels, E. R. & Szabadi, E. Functional neuroanatomy of the noradrenergic locus coeruleus: Its roles in the regulation of arousal and autonomic function part I: Principles of functional organization. Curr. Neuropharmacol. 6(3), 235–253. https://doi.org/10.2174/157015908785777229 (2008).
Leontovich, T. A. & Zhukova, G. P. The specificity of the neuronal structure and topography of the reticular formation in the brain and spinal cord of carnivora. J. Comp. Neurol. 121, 347–379. https://doi.org/10.1002/cne.901210305 (1963).
Das, G. D. & Kreutzberg, G. W. Evaluation of interstitial nerve cells in the central nervous system. A correlative study using acetylcholinesterase and Golgi techniques. Ergeb. Anat. Entwicklungsgesch 41(1), 3–58 (1969).
Kostovic, I. & Rakic, P. Cytology and time of origin of interstitial neurons in the white matter in infant and adult human and monkey telencephalon. J. Neurocytol. 9(2), 219–242. https://doi.org/10.1007/bf01205159 (1980).
Newman, J. Thalamic contributions to attention and consciousness. Conscious Cogn. 4(2), 172–193. https://doi.org/10.1006/ccog.1995.1024 (1995).
Kinomura, S., Larsson, J., Gulyás, B. & Roland, P. E. Activation by attention of the human reticular formation and thalamic intralaminar nuclei. Science 271(5248), 512–515. https://doi.org/10.1126/science.271.5248.512 (1996).
Minamimoto, T. & Kimura, M. Participation of the thalamic CM-Pf complex in attentional orienting. J. Neurophysiol. 87(6), 3090–3101. https://doi.org/10.1152/jn.2002.87.6.3090 (2002).
Schiff, N. D. Central thalamic contributions to arousal regulation and neurological disorders of consciousness. Ann. N. Y. Acad. Sci. 1129, 105–118. https://doi.org/10.1196/annals.1417.029 (2008).
Haber, S. N. & Calzavara, R. The cortico-basal ganglia integrative network: The role of the thalamus. Brain Res. Bull. 78(2–3), 69–74. https://doi.org/10.1016/j.brainresbull.2008.09.013 (2009).
Jones, E. G. & Leavitt, R. Y. Retrograde axonal transport and the demonstration of non-specific projections to the cerebral cortex and striatum from thalamic intralaminar nuclei in the rat, cat and monkey. J. Comp. Neurol. 154(4), 349–377. https://doi.org/10.1002/cne.901540402 (1974).
Macchi, G., Bentivoglio, M., Molinari, M. & Minciacchi, D. The thalamo-caudate versus thalamo-cortical projections as studied in the cat with fluorescent retrograde double labeling. Exp. Brain. Res. 54(2), 225–239. https://doi.org/10.1007/bf00236222 (1984).
Jayaraman, A. Organization of thalamic projections in the nucleus accumbens and the caudate nucleus in cats and its relation with hippocampal and other subcortical afferents. J. Comp. Neurol. 231(3), 396–420. https://doi.org/10.1002/cne.902310309 (1985).
Eckert, U. et al. Preferential networks of the mediodorsal nucleus and centromedian-parafascicular complex of the thalamus—a DTI tractography study. Hum. Brain Mapp. 33(11), 2627–2637. https://doi.org/10.1002/hbm.21389 (2012).
Su, H. S. & Bentivoglio, M. Thalamic midline cell populations projecting to the nucleus accumbens, amygdala, and hippocampus in the rat. J. Comp. Neurol. 297(4), 582–593. https://doi.org/10.1002/cne.902970410 (1990).
Green, J. D. & Adey, W. R. Electrophysiological studies of hippocampal connections and excitability. Electroencephalogr. Clin. Neurophysiol. 8(2), 245–263. https://doi.org/10.1016/0013-4694(56)90117-1 (1956).
Cavdar, S. et al. The pathways connecting the hippocampal formation, the thalamic reuniens nucleus and the thalamic reticular nucleus in the rat. J. Anat. 212(3), 249–256. https://doi.org/10.1111/j.1469-7580.2008.00858.x (2008).
Fotuhi, M., Do, D. & Jack, C. Modifiable factors that alter the size of the hippocampus with ageing. Nat. Rev. Neurol. 8(4), 189–202. https://doi.org/10.1038/nrneurol.2012.27 (2012).
Jessberger, S. Neural repair in the adult brain. F1000Res 12, 5. https://doi.org/10.12688/f1000research.7459.1 (2016).
Mogensen, J. Reorganization of the injured brain: Implications for studies of the neural substrate of cognition. Front. Psychol. 26(2), 7. https://doi.org/10.3389/fpsyg.2011.00007 (2011).
Chang, Y. Reorganization and plastic changes of the human brain associated with skill learning and expertise. Front. Hum. Neurosci. 4(8), 35. https://doi.org/10.3389/fnhum.2014.00035 (2014).
Kerr, A. L., Cheng, S. Y. & Jones, T. A. Experience-dependent neural plasticity in the adult damaged brain. J. Commun. Disord. 44(5), 538–548. https://doi.org/10.1016/j.jcomdis.2011.04.011 (2011).
Sharma, N., Classen, J. & Cohen, L. G. Neural plasticity and its contribution to functional recovery. Handb. Clin. Neurol. 110, 3–12. https://doi.org/10.1016/B978-0-444-52901-5.00001-0 (2013).
Leise, E. M. Modular construction of nervous systems: A basic principle of design for invertebrates and vertebrates. Brain Res. Brain Res. Rev. 15(1), 1–23. https://doi.org/10.1016/0165-0173(90)90009-d (1990).
Mota, B. & Herculano-Houzel, S. All brains are made of this: A fundamental building block of brain matter with matching neuronal and glial masses. Front. Neuroanat. 8, 127. https://doi.org/10.3389/fnana.2014.00127 (2014).
Santuy, A., Rodríguez, J. R., DeFelipe, J. & Merchán-Pérez, A. Study of the size and shape of synapses in the Juvenile Rat somatosensory cortex with 3D electron microscopy. eNeuro 5, 1. https://doi.org/10.1523/ENEURO.0377-17.2017 (2018).
Turner, D. A. & Adamson, D. C. Neuronal-astrocyte metabolic interactions: Understanding the transition into abnormal astrocytoma metabolism. J. Neuropathol. Exp. Neurol. 70(3), 167–176. https://doi.org/10.1097/NEN.0b013e31820e1152 (2011).
Okubo, Y. & Iino, M. Visualization of glutamate as a volume transmitter. J. Physiol. 589(Pt 3), 481–488. https://doi.org/10.1113/jphysiol.2010.199539 (2011).
Levitt, J. G. et al. Cortical sulcal maps in autism. Cereb. Cortex 13(7), 728–735. https://doi.org/10.1093/cercor/13.7.728 (2003).
Casanova, M. F., Buxhoeveden, D., Switala, A. & Roy, E. Neuronal density and architecture (gray level index) in the brains of autistic patients. J. Child. Neurol. 17(7), 515–521. https://doi.org/10.1177/088307380201700708 (2002).
Casanova, M. F. et al. Reduced gyral window and corpus callosum size in autism: Possible macroscopic correlates of a minicoluminopathy. J. Autism Dev. Disord. 39(5), 751–764. https://doi.org/10.1007/s10803-008-0681-4 (2009).
Behrendt, R. Hippocampus and consciousness. Rev. Neurosci. 24(3), 239–266. https://doi.org/10.1515/revneuro-2012-0088 (2013).
Hamani, C., Stone, S. S., Garten, A., Lozano, A. M. & Winocur, G. Memory rescue and enhanced neurogenesis following electrical stimulation of the anterior thalamus in rats treated with corticosterone. Exp. Neurol. 232(1), 100–104. https://doi.org/10.1016/j.expneurol.2011.08.023 (2011).
Stone, S. S. et al. Stimulation of entorhinal cortex promotes adult neurogenesis and facilitates spatial memory. J. Neurosci. 31(38), 13469–13484. https://doi.org/10.1523/JNEUROSCI.3100-11.2011 (2011).
Bezchlibnyk, Y. B., Stone, S. S. D., Hamani, C. & Lozano, A. M. High frequency stimulation of the infralimbic cortex induces morphological changes in rat hippocampal neurons. Brain Stimul. 10(2), 315–323. https://doi.org/10.1016/j.brs.2016.11.013 (2017).
Funding
This research was supported by the Croatian Science Fundation project CSF-IP-2020-02-4308, the European Union through the European Regional Development Fund, Operational Program Competitiveness and Cohesion, Grant agreement No. KK.01.1.1.01.0007, CoRE-Neuro. In addition, this research was supported by the Adris Foundation Grant “Application analysis of a robotic system for stereotactic neuronavigation”, leader Assistant Professor Darko Chudy, M.D., Ph.D.
Author information
Authors and Affiliations
Contributions
M.R. designed the study, conducted the volumetric analysis, and wrote the first version of the manuscript. N.P. designed the study and contributed to the statistical data analysis and interpretation of results. D.D., D.O. and A.R. conducted literature research, contributed to the data analysis, and designed the figures and tables. D.R., F.A. and P.M. contributed to the volumetric analysis and performed a neurological assessment of included patients. V.D., I.K., and D.C. contributed to the study concept and design, interpreted the results and revised the manuscript. All authors read and approved the final version of the manuscript as submitted.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Raguž, M., Predrijevac, N., Dlaka, D. et al. Structural changes in brains of patients with disorders of consciousness treated with deep brain stimulation. Sci Rep 11, 4401 (2021). https://doi.org/10.1038/s41598-021-83873-y
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-021-83873-y
This article is cited by
-
Deep brain stimulation in disorders of consciousness: 10 years of a single center experience
Scientific Reports (2023)
-
GABAergic Neurons in the Nucleus Accumbens are Involved in the General Anesthesia Effect of Propofol
Molecular Neurobiology (2023)
-
Acute traumatic coma awakening by right median nerve electrical stimulation: a randomised controlled trial
Intensive Care Medicine (2023)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
