
Abstract
Changes in intestinal microecology during acute liver failure (ALF) directly affect the occurrence and development of the disease. The study aimed to investigate the relationship between the intestinal microbiota and the key immune cells. Fecal microbiota transplantation (FMT) was used to determine whether ALF can balance Th17/Treg cytokines. The relationship between gut microbiota and clinical indicators was analyzed. BALB/c mice were treated with d-galactosamine (d-GalN) to induce a murine ALF model. FMT to d-GalN mice was conducted to test for liver function indicators. Results showed that the proportions of Lachnospiraceae, Prevotella, S24-7, Odoribacter and Rikenellaceae in d-GalN mice with intestinal microbiota disorder were restored after FMT. Further, CIA analysis showed that bacteria had a covariant relationship with clinical indicators. Microbiota could account for changes in 49.9% of the overall clinical indicators. Adonis analysis showed that Ruminococcus, and Enterococcus have a greater impact on clinical indicators. FMT down-regulated the expression of IL-17A, TNF-α, and TGF-β, while up-regulated IL-10 and IL-22. Transplantation of feces from Saccharomyces boulardii donor mice improved GalN-induced liver damage. These findings indicate that FMT attenuates d-GalN-induced liver damage in mice, and a clinical trial is required to validate the relevance of our findings in humans, and to test whether this therapeutic approach is effective for patients with ALF.
Similar content being viewed by others
Introduction
Acute liver failure (ALF) is a rare but life-threatening clinical syndrome characterized by the rapid death of large numbers of hepatocytes because of cell necrosis or apoptosis1. This loss of cells ultimately leads to functional impairment of the liver. The most common factors that contribute to the development of ALF are hepatitis, medication overdose, idiosyncratic drug reactions, and metabolic disorders2. Liver transplant remains the most effective solution for treating ALF. However, the scope of this treatment is limited by extremely high cost and shortage of liver donors, and hence the development of alternative therapies is urgently needed3.
The intestinal microbiota is inseparable from the body, whose activities are linked to numerous host functions4,5,6,7. These bacteria contribute to the microenvironment of the intestine via their metabolic activities. Accumulated evidence has demonstrated the important effects of gut microbes on disease, and has provided the rationale behind the hypothesis that managing microbiota may help treat liver diseases8,9,10. Previous studies reported that the gut microbiota play an important role in the pathogenesis of liver diseases. In a BALB/c mouse model, studies have indicated that metabolites produced by gut microbiota are responsible for the diurnal variation of acetaminophen-induced hepatotoxicity11. Studies have also shown that Rhubarb extract prevent alcohol-induced hepatic inflammation by modifying the gut microbiota12. In addition, T helper 17 (Th17) and regulatory T cells (Treg) are key immune cell subsets that link gut microbiota and intestinal immune function. Recent evidence suggests that changes in Th17/Treg balance plays a major role in the development and progression of inflammatory diseases13. Thus, the effects of fecal microbiota transplantation (FMT) on Th17/Treg and gut microbiota need to be investigated.
In recent years, FMT has been widely studied as an alternative treatment. Ferrere et al. showed that fecal transplant from alcohol-resistant donor mice restore gut homeostasis and prevent alcoholic liver disease14. Wang et al. reported that FMT prevents hepatic encephalopathy in a rat model15. Using humanized mice, Duan et al. reported that bacteriophages can specifically target cytolytic E. faecalis, which decrease cytolysin in the liver and abolish ethanol-induced liver disease16. Furthermore, a clinical trial showed that FMT from a rationally selected donor reduced hospitalizations, improved cognition, and dysbiosis in cirrhosis with recurrent hepatic encephalopathy17. However, there is limited evidence about the effects of FMT on ALF. In our previous study, we demonstrated that Saccharomyces boulardii administration can change the gut microbiota and attenuate drug-induced liver injury in a rodent model18. To further investigate whether the regulation of intestinal microbiota by Saccharomyces boulardii alleviate liver failure, we performed this study.We hypothesized that an association exists between gut microbiome and the outcome of d-galactosamine (d-GalN)-induced ALF. To test this hypothesis, we used fecal bacteria transplantation to treat mice with ALF, and analyzed liver function, pathology, and serum cytokines. 16S rDNA sequencing was performed to detect intestinal microbial diversity, and qRT-PCR, western blotting, and immunohistochemistry were performed to verify Th17/Treg cytokines. Overall, we provide an experimental basis for evaluating the potential clinical application of FMT in ALF patients.
Results
Fecal transplantation alters serum AST, ALT, and TBIL levels
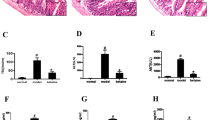
Four groups of mice were studied, including a CTRL group, a d-GalN group with d-GalN-induced liver injury, a d-GalN + SS group and a d-GalN + SB group with FMT treatment at d-GalN challenge. Plasma levels of ALT, AST, and TBIL were measured as an indicator of d-GalN-induced liver injury (Fig. 1). Compared with the CTRL group, there was a significant elevation in the levels of ALT, AST, and TBIL in the d-GalN group (p < 0.05 for ALT, AST), indicating massive abnormality in liver function. However, ALT, AST, and TBIL levels in the d-GalN + SS and d-GalN + SB group were significantly lower than those in the d-GalN group (p < 0.05); the levels decreased significantly in d-GalN + SB group compared with d-GalN + SS group (p < 0.05), suggesting marked attenuation in liver injury after FMT. The d-GalN + SB group was superior to the d-GalN + SS group (p < 0.05).

Effects of fecal microbiota transplantation (FMT) on (A) ALT, (B) AST, and (C) TBIL activities.
Fecal transplantation alters serum IL-10, IL-17A, TNF-α, and TGF-β levels
IL-10, IL-17A, TNF-α, and TGF-β levels in the serum of mice are shown in Table 1. Compared with serum IL-10 and TNF-α levels in d-GalN group, those in the d-GalN + SS and the d-GalN + SB groups were significantly increased (p < 0.05), while IL-17A and TGF-β levels were significantly reduced (p < 0.05). Compared with serum IL-17A and TGF-β levels in d-GalN + SS group, those in d-GalN + SB group were significantly decreased (p < 0.05).
Histopathological analysis of liver tissues after fecal transplantation
Histology of the mice liver sections exhibited a normal lobular liver architecture and integrated cell structure in CTRL group (Fig. 2A). Challenge with d-GalN resulted in acute hepatic injury accompanied by prominent hemorrhage and inflammation, necrosis of hepatocytes, and serious dissolution of the hepatocyte architecture (Fig. 2B). Such liver alterations were alleviated in the d-GalN + SS group and d-GalN + SB group (Fig. 2C,D). Such differences indicated that FMT from proper donors indeed alleviated GalN-induced liver injury and restored hepatocyte structure.

Microscopic examination of liver sections after FMT. (A) Histology of control group with abnormal liver architecture. (B) Histologically, d-GalN group showed obvious hemorrhage and inflammation, necrosis of hepatocytes, and severe dissolution of hepatocyte structure. (C) d-GalN-induced damage was partially restored in d-GalN + SS group. (D) Hepatic injury was markedly attenuated in d-GalN + SB group.
Changes in gut microbiota after fecal transplantation
In order to further study the underlying mechanisms for the protective effects of FMT, we performed 16S rDNA sequencing and examined the temporal changes to the composition of gut microbiota. The metagenomic DNA was quantified and mapped into operational taxonomic units (OTUs) for analysis. A principal coordinate analysis (PCoA) of Bray–Curtis dissimilarities (Fig. 3A) showed clear separation between samples collected after FMT in the d-GalN + SS and d-GalN + SB groups, indicating a change in gut microbiota.

(A) Principal component analysis. (B) Gut microbiota at the pylums level. (C) Gut microbiota at the family level. (D) Results of LEfSe analysis.
At the phylum level, we found that the d-GalN + SB group was associated with a significant decrease in the relative abundance of Bacteroidetes and a significant increase in Firmicutes and Proteobacteria compared to the d-GalN group (p < 0.05, Fig. 3B). These results suggested that FMT from the donor mice gavaged with Saccharomyces boulardii changed the gut microbial community by altering the proportion of three major phyla. At the family level, we also observed several important modifications of the gut microbial composition. Among the major taxonomic families identified, Lachnospiraceae and Prevotella were increased following FMT treatment. Conversely, S24-7, Odoribacter, and Rikenellaceae were significantly decreased (p < 0.05, Fig. 3C).
LEfSe analysis of d-GalN group and d-GalN + SB group revealed that the cladogram and reflecting relative bacterial abundance in the d-GalN + SB group (Fig. 3D) exhibited that Staphylococcus and Parasutterella were overrepresented genera after FMT. Lefse analyses for d-GalN + SS versus d-GalN + SB and d-GalN versus d-GalN + SS were shown in Supplementary Fig. S1. In contrast, a decrease of Anaeroplasma and Ruminococcus genera occurred in response to FMT. Such temporal changes may explain the microbial mechanisms underlying the protective effects of FMT.
Relationship between microbiota and clinical indicators after fecal transplantation
Using co-inertia (CIA) analysis, it was found that bacteria had a covariant relationship with clinical indicators. The influence of bacteria on clinical indicators was further quantified. There was a significant correlation between bacteria and phenotypes (p < 0.01, RV coefficient: 0.238), bacteria could explain 49.9% of changes in overall clinical indicators (Fig. 4A). Using Adonis analysis, we found a complex relationship between the microbiota and clinical indicators. Bacteria from 15 genera such as Ruminococcus, Enterococcus, Veillonellaceae, Klebsiella, Comamonas, Adlercreutzia, Clostridium, Lactobacillus was found to affect the clinical indicators. Further analysis of the first four bacteria in each group of mice showed that compared with the CTRL group, Ruminococcus and Enterococcus in the d-GalN group were significantly lower (p < 0.05), compared with the d-GalN group. The difference of d-GalN + SB group was statistically significant (p < 0.05). There was no significant difference between Veillonellaceae and Klebsiella (p > 0.05) (Fig. 4B–E). Through Random Forest association analysis, bacteria with obvious positive correlation with liver function were found to be Veillonellaceae, Prevotellaceae, Coriobacteriaceae, and F16, whereas those with obvious negative correlation were Lactobacillus, Corynebacterium, Methanobrevibacter, and Megasphaera (Fig. 4F).

Covariant relationship of bacteria with clinical indicators. (A) Co-inertia analysis. (B–E) Adnois analysis. (F) Random forest model.
Fecal transplantation alleviates ALF via Th17/Treg
Immunohistochemical results showed that the expression of TNF-α, TGF-β, and IL-17A in the d-GalN group was higher than that in the CTRL group (Fig. 5A). The expressions of IL-10 and IL-22 in d-GalN + SS group and d-GalN + SB group were lower than that in d-GalN group. The expressions of TNF-α, TGF-β and IL-17A in the d-GalN + SB group were lower than that in the d-GalN + SS group, while the expressions of IL-10 and IL-22 were higher than that in the d-GalN + SS group (Fig. 5B–D). In addition, qRT-PCR and western blot results showed that the mRNA and protein expressions of TNF-α, TGF-β, and IL-17A in the d-GalN group were higher than those in the CTRL group (p < 0.05).The levels of mRNA and protein expressions of IL-10 and IL-22 in d-GalN + SS group and d-GalN + SB group were lower than those in d-GalN group (p < 0.05). In comparison, the mRNA and protein expressions of TNF-α, TGF-β, and IL-17A in the d-GalN + SB group were lower than those in the d-GalN + SS group, and the mRNA and protein expressions of IL-10 and IL-22 were higher than those in d-GalN + SS group, including TGF-β, IL-10, and IL-22 (p < 0.05, Fig. 5B–D).

Expression levels of TNF-α, TGF-β, IL-17A, IL-10, and IL-22 in liver tissues of each group. (A) Immunohistochemical analysis of TNF-α, TGF-β, IL-17A, IL-10, and IL-22 in each group (× 400). (B) The mRNA expression of TNF-α, TGF-β, IL-17A, IL-10, and IL-22 in each group by qPCR. (C) Western blotting for TNF-α, TGF-β, IL-17A, IL-10, and IL-22. (D) Gray ratio by western blotting. *P < 0.05 compared with control group; #P < 0.05 compared with d-GalN group; △P < 0.05 compared with D-GalN + SS group.
Discussion
ALF is a life-threatening condition with high morbidity and mortality rates. However, some patients have been found to spontaneously recover, suggesting that other factors other than the hepatic physiology affect the progression of ALF. The involvement of microbiota in liver diseases was largely described in rodent models or in humans19,20,21. In addition, variability in the susceptibility to develop ALF has also been observed in animal models bearing different microbiota22.
Given the limitations of existing ALF treatments, large efforts are now aimed at developing novel therapies. FMT is rapidly accepted as a viable, safe, and effective treatment for various liver conditions23,24. However, whether FMT has a healing effect on ALF has not been sufficiently investigated. In a previous study, we reported that Saccharomyces boulardii administration could change the gut microbiota in mice and alleviate acute liver failure18. The results of our current study further demonstrate that the protective function of Saccharomyces boulardii can be exerted not only by direct administration, but also by indirect FMT. In our animal model, d-GalN-induced abnormal aminotransferase activities and serious dissolution of the hepatocyte architecture were effectively attenuated by FMT. In addition, serum IL-10, IL-17A, and TNF-αwere significantly reduced and were found to increase after FMT intervention. Previously studies have reported increased level of TGF-β after FMT25,26, while we found that TGF-β was significantly reduced. It is possible that FMT is inducing an anti-inflammatory environment, thus leading to less requirement of TGF-β27.
To elucidate the underlying mechanisms, we examined the changes to the composition and diversity of the gut microbiota. Results showed that the proportion of certain genera in the gut microbiota was significantly changed by FMT, some of which are supported by published research findings. For example, we observed an increase in Parasutterella genus after FMT, which is consistent with the reported low Parasutterella levels in animal models of alcoholic liver disease28 and fatty liver disease29. The decrease in Anaeroplasma after FMT in our model is consistent with reported enrichment of Anaeroplasma in a nonalcoholic steatohepatitis mouse model30. Through CIA analysis, a significant correlation was observed between the micromicrobiota and clinical indicators, among which 49.9% clinical changes could be explained by bacteria. Further, effect-size analyses and Adonis analysis shows that 15 bacterial genera are dominant, and the most influential are Ruminococcus and Enterococcus. Ruminococcus can participate in the biometabolism, and transport and catabolism of the secondary metabolites31. Enterococcus is an intestinal bacterium found under normal physiological conditions. It ferments carbohydrates, produces lactic acid, but does not produce gas, and also ferments lactose. Analysis of Ruminococcus and Enterococcus showed that their proportion in d-GalN + SS and d-GalN + SB groups increased after FMT, and the difference in d-GalN + SB group is statistical significant (p = 0.012), indicating a protective effect of FMT.
In addition, we have confirmed that FMT reduced liver injury by increasing the Lachnospiraceae population and reducing S24-7 population to restore intestinal microbiota composition. Lachnospiraceae is a probiotic that can produce short-chain fatty acids (SCFAs). Studies have shown that SCFAs can regulate Treg cells to reduce inflammation32. Therefore, we used qRT-PCR, western blotting, and immunohistochemistry to investigate the changes in the expression of Th17/Treg cytokines in the liver induced by fecal bacteria transplantation. IL-17A, TNF-α, and TGF-β expression in the d-GalN group was significantly higher than that in the CTRL group (p < 0.05), whereas IL-10 expression was significantly lower than that in the CTRL group (p < 0.05). Compared with that in d-GalN group, IL-17A, TNF-α, and TGF-β expression in d-GalN + BB group was significantly decreased (p < 0.05), whereas IL-10 expression showed a significant increase (p < 0.05). The results are similar to those reported by Zhou et al.33. Recently, a prospective randomized trial reported fecal microbiome transplant in patients with cirrhosis reduced E. feacalis in stool and associated with a decrease in systemic inflammation34. These results suggest that FMT can adjust the balance of Th17/Treg cytokines in the liver tissue to improve d-GalN induced acute liver damage.
Some limitations of this study should be addressed. The impact of FMT cannot be well separated from the impact of antibiotic pretreatment. Detailed mechanisms of FMT on ALF treatment remain a matter of speculation, thus the effects of FMT on intestinal barrier and cells need to further elucidated. Ideally we would like investigate down to a specific bacterial strain. However this is not possible for the present study.
Taken together, this study provided an in-depth analysis of gut microbiota modulations after FMT. Our results demonstrated that FMT from donors could alter the gut microbiota in mice and alleviate d-GalN-induced acute liver injury, indicating a potential therapeutic strategy. Further transcriptomic and clinical research is required to better understand the underlying mechanisms of the hepatic protective effects of FMT.
Materials and methods
Mice treatment and tissue sampling
Male BALB/c mice at 4 weeks of age were purchased from Guizhou Medical University (Guizhou, China) and then individually housed in plastic cages for 2 weeks at room temperature (23 °C) with an artificial cycle of 12-h light and 12-h dark. At the end of each experiment, the mice were sacrificed by cervical dislocation after anaesthetization. Fresh cecal content, plasma, and liver tissue were collected and stored at − 80 °C until analyses. The above procedures were approved by and performed in accordance with the guidelines of the Ethics Committee of the Hospital Affiliated to Guizhou Medical University.
Study design of fecal transplantation
d-GalN was purchased from Sigma Aldrich Corporation (St. Louis, Missouri, USA) to establish a mouse model of acute liver injury. All the mice were randomly divided into the following 4 groups (N = 5 in each group): (1) mice that served as vehicle control (CTRL group); (2) mice that were treated with d-GalN (d-GalN group); (3) mice with d-GalN and fecal transplants from donor mice gavaged with saline solution (d-GalN + SS group); (4) mice that were treated with d-GalN and fecal transplants from donor mice gavaged with Saccharomyces boulardii (d-GalN + SB group). The d-GalN group, d-GalN + SS group, and d-GalN + SBSB group were intraperitoneally (i.p.) injected with 3.0 g/kg d-GalN, while the CTRL group were injected with saline solution.
The feces of donor mice were collected fresh every day and FMT was conducted under the aseptic conditions using a laminar flow hood. The feces from the two groups of donors were collected in cages, and 100 mg (approximately 5–6 fecal particles) was suspended in 1 mL sterile normal saline. The solution was vigorously blended for 10 s before centrifugation at 800×g for 3 min. The supernatant fluid (approximately 500–600 microns) was collected and fed to the mice over 10 times.
According to the established microbial depletion and recolonization protocol35,36,37,38, a combination of ciprofloxacin (0.2 g/L) and metronidazole (1 g/L) were added to the drinking water of FMT recipients for 2 days before FMT. Each recipient was given 100 mL of fecal supernatant orally 48 h after d-GalN injection, and was given FMT once every 24 h, a total of 7 times. The serum, liver and intestinal flora of the two groups were compared. Serum samples, liver tissue specimens, and gut microbiota were collected.
Serum a minotransferase activities and enzyme-linked immunosorbent assay (ELISA)
Fasting blood was collected from each mouse and centrifuged at 1000×g for 5 min at room temperature. Serum sample was extracted and stored at − 20 °C until further analysis. Serum alanine aminotransferase (ALT), aspartate aminotransferase (AST), and total bilirubin (TBIL) activities were quantified with the enzymatic kinetic method by using a HORRON RD171 semi-automatic biochemistry analyzer (HORRON XLH Medical Electronics, China) according to the manufacturer’s protocol.
To evaluate the role of Treg/Th17-driven inflammation, serum cytokines from Treg/Th17 cells, including interleukin IL-10, IL-17A, TNF-α, and TGF-β, were measured by a commercially available ELISA kit (k3605) from Shanghai Hengke Biotechnology Co., Ltd. (Shanghai, China). Data was analyzed by microplate reader (DR-200B).
Histologic examination
Histologic examination was carried out following our previous protocols6. Briefly, a 0.2 cm × 0.2 cm tissue specimen was taken from the right liver lobe from each mouse. All specimens were dehydrated by using graded solutions of alcohol, fixed in pH 7.4 and 10% buffered neutral formalin, and embedded in molten paraffin wax. After hematoxylin and eosin staining, the morphologic evaluation was performed under 10 × with a light microscope (SP2, Leica). Stained tissue sections were blind reviewed by two pathologists based on a double scoring system.
DNA isolation and sequencing
Metagenomic DNA was extracted from the fcecal contents using a QIAamp-DNA stool minikit (Qiagen, Germany) according to the manufacturer’s instructions and previously published protocols39. Real-time qPCR was performed using TaqMan Universal Master Mix (Life Technologies, USA) to examine the quantity and the quality of the DNA extracted from the samples. According to the 16S metagenomic sequencing library preparation guide (Illumina, USA), total DNA was amplified using primers targeting the 16S V3 and V4 regions under the following conditions: 95 °C for 3 min, followed by 30 cycles at 95 °C for 30 s, 54 °C for 30 s, and 72 °C for 30 s, and a final extension at 72 °C for 5 min. Pooled V3-V4 amplicon libraries were sequenced using the Illumina MiSeq platform with a V3 reagent kit.
Fecal microbial analysis
A 16S V3 + V4 region library was constructed by PCR amplification of the 16S rDNA using V3 − V4 specific primers. Sequence data for at least 50,000 paired-end read pairs from each sample were acquired using the Illumina HiSeq sequencing platform. The sequence data were then subjected to filtering for read quality and length, removing primer sequences, and splicing paired-end reads to obtain high quality data for OTU clustering and subsequent analysis. Cluster analysis of species abundance was performed using the QIIME1.91 software package, and phylogenetic groups were classified using RDP and Green gene databases.
Real-time quantitative polymerase chain reaction (qPCR)
Total RNA was extracted from liver tissue and cDNA was synthesized using RNA reverse transcriptase. All primers were provided by Shanghai Sangon biological engineering co., LTD (Shanghai, China). The β-actin housekeeping gene was used as an internal reference to quantitatively determine the Ct value.
Western blotting
Protein concentration in the liver lysates was determined using a BCA kit (Pierce, Rockford, IL). Lysates were electrophoresed on a reducing 12% SDS-PAGE, transferred to an Immobilon-P membrane (Millipore Corp, Bedford, MA), incubated with the antibodies of IL-10 (1:800, Beijing Boosen Biotechnology co., LTD, Beijing, China), IL-17A (1:1000), IL-22 (1:800), TNF-α (1:800), and TNF-β (1:800), and then developed using the enhanced chemiluminescence (ECL) system (Pharmacia Biotech, Piscataway, NJ). The expression of the β-actin protein expression was used as a loading control. Individual band densities were analyzed as follow: We quantified the signal intensity of a specific band on the developed image using Adobe Photoshop and then averaged the intensity readings from three independent experiments. The intensity of the image background was subtracted prior to pooling.
Immunohistochemical staining
Immunohistochemical staining was performed on sections from paraffin embedded liver tissues, according to the method of Martinet et al.40. The IL-10, IL-17A, IL-22, TNF-α, and TGF-β anti-mouse monoclonal antibodies were diluted to 1:600. The sections were stained with DAB and re-dyed with hematoxylin. Images were captured via an Olympus BX51, the optical density was measured using a MIAS (MIAS 2000, Olympus, Tokyo, Japan), and the mean optical density for each section was calculated from five randomly visualized fields, under 400 ×.
Statistical analysis
Data were analyzed via SPSS 19.0 (SPSS Inc., Chicago, IL, USA) and shown as mean ± standard deviation (SD). The groups were compared by one-way ANOVA, and p < 0.05 was considered to indicate a statistically significant difference. Weighted Fast UniFrac principal coordinate analysis (PCoA) based on OTUs was performed to provide an overview of gut microbial dynamics after fecal transplantation. Between-group beta-diversity was examined using LEfSe software41, in order to detect features with significantly different abundances between assigned classes. Analysis of bacterial population and clinical indicators using were performed using effect size analyses analysis, Adonis analysis, co-inertia analysis (CIA) analysis42.
References
Bunchorntavakul, C. & Reddy, K. R. Acute liver failure. Clin. Liver Dis. 21, 769–792. https://doi.org/10.1016/j.cld.2017.06.002 (2017).
Blackmore, L. & Bernal, W. Acute liver failure. Clin. Med. (Lond) 15, 468–472. https://doi.org/10.7861/clinmedicine.15-5-468 (2015).
Bernal, W., Lee, W. M., Wendon, J., Larsen, F. S. & Williams, R. Acute liver failure: A curable disease by 2024?. J. Hepatol. 62, S112-120. https://doi.org/10.1016/j.jhep.2014.12.016 (2015).
Nicholson, J. K. et al. Host-gut microbiota metabolic interactions. Science 336, 1262–1267 (2012).
Kau, A. L., Ahern, P. P., Griffin, N. W., Goodman, A. L. & Gordon, J. I. Human nutrition, the gut microbiome and the immune system. Nature 474, 327–336 (2011).
Jumpertz, R. et al. Energy-balance studies reveal associations between gut microbes, caloric load, and nutrient absorption in humans. Am. J. Clin. Nutr. 94, 58–65 (2011).
Gordon, J. I., Dewey, K. G., Mills, D. A. & Medzhitov, R. M. The human gut microbiota and undernutrition. Sci. Transl. Med. 4, 137 (2012).
Goel, A., Gupta, M. & Aggarwal, R. Gut microbiota and liver disease. J. Gastroenterol. Hepatol. 29, 1139–1148. https://doi.org/10.1111/jgh.12556 (2014).
Marchesi, J. R. et al. The gut microbiota and host health: a new clinical frontier. Gut 65, 330–339. https://doi.org/10.1136/gutjnl-2015-309990 (2016).
Bajaj, J. S. Alcohol, liver disease and the gut microbiota. Nat. Rev. Gastroenterol. Hepatol. 16(4), 235–246. https://doi.org/10.1038/s41575-018-0099-1 (2019).
Gong, S. et al. Gut microbiota mediates diurnal variation of acetaminophen induced acute liver injury in mice. J. Hepatol. 69, 51–59. https://doi.org/10.1016/j.jhep.2018.02.024 (2018).
Neyrinck, A. M. et al. Rhubarb extract prevents hepatic inflammation induced by acute alcohol intake, an effect related to the modulation of the gut microbiota. Mol. Nutr. Food Res. https://doi.org/10.1002/mnfr.201500899 (2017).
Luo, A. et al. The microbiota and epigenetic regulation of T helper 17/regulatory T cells: in search of a balanced immune system. Front. Immunol. 8, 417. https://doi.org/10.3389/fimmu.2017.00417 (2017).
Ferrere, G. et al. Fecal microbiota manipulation prevents dysbiosis and alcohol-induced liver injury in mice. J. Hepatol. 66, 806–815. https://doi.org/10.1016/j.jhep.2016.11.008 (2017).
Wang, W. W. et al. Fecal microbiota transplantation prevents hepatic encephalopathy in rats with carbon tetrachloride-induced acute hepatic dysfunction. World J. Gastroenterol. 23, 6983–6994. https://doi.org/10.3748/wjg.v23.i38.6983 (2017).
Duan, Y. et al. Bacteriophage targeting of gut bacterium attenuates alcoholic liver disease. Nature 575(7783), 505–511. https://doi.org/10.1038/s41586-019-1742-x (2019).
Bajaj, J. S. et al. Fecal microbiota transplant from a rational stool donor improves hepatic encephalopathy: A randomized clinical trial. Hepatology 66(6), 1727–1738. https://doi.org/10.1002/hep.29306 (2017).
Yu, L. et al. Saccharomyces boulardii administration changes gut microbiota and attenuates D-galactosamine-induced liver injury. Sci. Rep. 7, 1359. https://doi.org/10.1038/s41598-017-01271-9 (2017).
Acharya, C. & Bajaj, J. S. Gut microbiota and complications of liver disease. Gastroenterol. Clin. North Am. 46, 155–169. https://doi.org/10.1016/j.gtc.2016.09.013 (2017).
Acharya, C., Sahingur, S. E. & Bajaj, J. S. Microbiota, cirrhosis, and the emerging oral-gut-liver axis. JCI Insight https://doi.org/10.1172/jci.insight.94416 (2017).
Federico, A., Dallio, M., Caprio, G. G., Ormando, V. M. & Loguercio, C. Gut microbiota and the liver. Minerva Gastroenterol. Dietol. 63, 385–398. https://doi.org/10.23736/S1121-421X.17.02375-3 (2017).
Llopis, M. et al. Intestinal microbiota contributes to individual susceptibility to alcoholic liver disease. Gut 65, 830–839. https://doi.org/10.1136/gutjnl-2015-310585 (2016).
Bouri, S. & Hart, A. Fecal microbial transplantation: An update. Curr. Opin. Clin. Nutr. Metab. Care 21, 405–410. https://doi.org/10.1097/MCO.0000000000000488 (2018).
Yan, Z. X. et al. Fecal microbiota transplantation in experimental ulcerative colitis reveals associated gut microbial and host metabolic reprogramming. Appl. Environ. Microbiol. https://doi.org/10.1128/AEM.00434-18 (2018).
Wei, Y. L. et al. Fecal microbiota transplantation ameliorates experimentally induced colitis in mice by upregulating AhR. Front. Microbiol. 9, 1921. https://doi.org/10.3389/fmicb.2018.01921 (2018).
Cheng, S. et al. Fecal microbiota transplantation beneficially regulates intestinal mucosal autophagy and alleviates gut barrier injury. mSystems 3(5), e00137-18. https://doi.org/10.1128/mSystems.00137-18 (2018).
Foligné, B. et al. Contribution of the gut microbiota in P28GST-mediated anti-inflammatory effects: Experimental and clinical insights. Cells 8(6), 577. https://doi.org/10.3390/cells8060577 (2019).
Zhang, X. et al. Flaxseed oil ameliorates alcoholic liver disease via anti-inflammation and modulating gut microbiota in mice. Lipids Health Dis. 16, 44. https://doi.org/10.1186/s12944-017-0431-8 (2017).
Xie, G. et al. Distinctly altered gut microbiota in the progression of liver disease. Oncotarget 7, 19355–19366. https://doi.org/10.18632/oncotarget.8466 (2016).
Ye, J. Z. et al. Dynamic alterations in the gut microbiota and metabolome during the development of methionine-choline-deficient diet-induced nonalcoholic steatohepatitis. World J. Gastroenterol. 24, 2468–2481. https://doi.org/10.3748/wjg.v24.i23.2468 (2018).
Gosalbes, M. J. et al. Metatranscriptomic approach to analyze the functional human gut microbiota. PLoS ONE 6, e17447. https://doi.org/10.1371/journal.pone.0017447 (2011).
Smith, P. M. et al. The microbial metabolites, short-chain fatty acids, regulate colonic Treg cell homeostasis. Science 341, 569–573. https://doi.org/10.1126/science.1241165 (2013).
Zhou, D. et al. Total fecal microbiota transplantation alleviates high-fat diet-induced steatohepatitis in mice via beneficial regulation of gut microbiota. Sci. Rep. 7, 1529. https://doi.org/10.1038/s41598-017-01751-y (2017).
Edwards, L. et al. Faecal microbiota transplantation reduces pathogenic burden and is anti-inflammatory in patients with advanced cirrhosis. J. Hepatol. 73, S77–S78. https://doi.org/10.1016/S0168-8278(20)30770-4 (2020).
Bruce-Keller, A. J. et al. Obese-type gut microbiota induce neurobehavioral changes in the absence of obesity. Biol. Psychiat. 77, 607–615. https://doi.org/10.1016/j.biopsych.2014.07.012 (2015).
McCafferty, J. et al. Stochastic changes over time and not founder effects drive cage effects in microbial community assembly in a mouse model. ISME J. 7, 2116–2125. https://doi.org/10.1038/ismej.2013.106 (2013).
Reikvam, D. H. et al. Depletion of murine intestinal microbiota: Effects on gut mucosa and epithelial gene expression. PLoS ONE 6, e17996. https://doi.org/10.1371/journal.pone.0017996 (2011).
Suez, J. et al. Artificial sweeteners induce glucose intolerance by altering the gut microbiota. Nature 514, 181–186. https://doi.org/10.1038/nature13793 (2014).
Dewulf, E. M. et al. Insight into the prebiotic concept: lessons from an exploratory, double blind intervention study with inulin-type fructans in obese women. Gut 62, 1112–1121. https://doi.org/10.1136/gutjnl-2012-303304 (2013).
Martinet, W., Roth, L. & De Meyer, G. R. Y. Standard immunohistochemical assays to assess autophagy in mammalian tissue. Cells https://doi.org/10.3390/cells6030017 (2017).
Segata, N. et al. Metagenomic biomarker discovery and explanation. Genome Biol. 12, R60. https://doi.org/10.1186/gb-2011-12-6-r60 (2011).
Zhang, C. et al. Dietary modulation of gut microbiota contributes to alleviation of both genetic and simple obesity in children. EBioMedicine 2, 968–984. https://doi.org/10.1016/j.ebiom.2015.07.007 (2015).
Funding
This study was supported by National Nature Science Foundation of China (Nos. 81960118, 81570543 and 81760116).
Author information
Authors and Affiliations
Contributions
Y.M.L., M.L.C., L.Y and X.K.Z. designed all experiments. Y.M.L., L.D.F., S.C., L.L.Z. and B.F.Z. performed the experiments. Y.M.L., Z.C., Y.X.H. and P.L.Z. performed the bioinformatics data analyses. Y.J.C. and L.D.F. provided technical assistance. Y.M.L., Z.C., M.L.C. and L.Y wrote, edited, and proofread the manuscript. All authors read and approved the final manuscript.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Liu, Y., Fan, L., Cheng, Z. et al. Fecal transplantation alleviates acute liver injury in mice through regulating Treg/Th17 cytokines balance. Sci Rep 11, 1611 (2021). https://doi.org/10.1038/s41598-021-81263-y
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-021-81263-y
This article is cited by
-
Impact of the gut microbiota and associated metabolites on cardiometabolic traits, chronic diseases and human longevity: a Mendelian randomization study
Journal of Translational Medicine (2023)
-
Gut microbiota modulates lung fibrosis severity following acute lung injury in mice
Communications Biology (2022)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
