
Abstract
The potential for a Trichoderma-based compost activator was tested for in-situ rice straw decomposition, under both laboratory and field conditions. Inoculation of Trichoderma caused a 50% reduction in the indigenous fungal population after 2 weeks of incubation for both laboratory and field experiments. However, the Trichoderma population declined during the latter part of the incubation. Despite the significant reduction in fungal population during the first 2 weeks of incubation, inoculated samples were found to have higher indigenous and total fungal population at the end of the experiments with as much as a 300% increase in the laboratory experiment and 50% during day-21 and day-28 samplings in the field experiment. The laboratory incubation experiment revealed that inoculated samples released an average of 16% higher amounts of CO2 compared to uninoculated straw in sterile soil samples. Unsterile soil inoculated with Trichoderma released the highest amount of CO2 in the laboratory experiment. In the field experiment, improved decomposition was observed in samples inoculated with Trichoderma and placed below ground (WTBG). From the initial value of around 35%, the C content in WTBG was down to 28.63% after 42 days of incubation and was the lowest among treatments. This is significantly lower compared with NTBG (No Trichoderma placed below ground, 31.1% C), WTSS (With Trichoderma placed on soil surface, 33.83% C), and NTSS (No Trichoderma placed on soil surface, 34.30% carbon). The WTBG treatment also had the highest N content of 1.1%. The C:N ratio of WTBG was only 26.27, 39.51% lower than the C:N ratio of NTBG, which is 43.43. These results prove that the Trichoderma-based inoculant has the potential to hasten the decomposition of incorporated rice straw.
Similar content being viewed by others

Introduction
Rice is one of the most important agricultural commodities in the Philippines and other parts of the world, particularly Asia1. However, the production of this staple grain to feed the growing population also produces many tons of waste. In 2009, it was reported that 242, 97, 22, and 11 MT of rice straw residue were produced annually in China, India, Thailand, and the Philippines, respectively2. To eliminate this residue, one cost-effective method that is widely practiced is burning rice straw in the field2. In the Philippines alone, 95% of the residue undergoes open-field burning2. Rice straw burning is widely employed because it is less laborious compared to other practices such as straw incorporation into the soil3. Burning can also eliminate many pathogens4, and, most importantly, it results in rapid and complete residue removal5. While burning seems to be the most practical way of removing the straw, this practice has several undesirable effects including loss of major nutrients6. The nutrient loss caused by straw burning can reach 80% for nitrogen (N)7, 25% for phosphorus (P)8, 21% for 33 potassium (K)9, and 4–60% for sulfur (S)10. In addition, biomass burning is the second-largest source of trace gases and the largest source of primary fine carbonaceous particles in the global troposphere11,12,13,14. A recent Life Cycle Assessment (LCA) of existing in-field and off-field rice straw management practices reported that rice cultivation with in-field burning is the worst option with the lowest energy efficiency and highest air pollution emission15.
The use of modern farm equipment enables farmers to easily manage farm operations. For instance, the combine harvester combines harvesting operations—reaping, threshing, and winnowing into a single process. The use of combine harvesters has therefore been used for rice and has expanded rapidly worldwide and major producers, such as Vietnam and Cambodia, almost exclusively rely on them16. Because of this technology, harvesting can be performed even with minimal manpower and the chopped rice straw can be incorporated into the soil for decomposition.
Rice straw incorporation can solve environmental pollution caused by straw burning. In a meta-analysis in China which compared the effects of straw burning and incorporation on the Net Global Warming Potential (NGWP) to include C sequestration, it was found that switching from burning to straw incorporation could mitigate 34.18 Mt. CO2 eq year−1 or 31% of total emissions in the country17,18,19. Studies show that early incorporation is one the most cost-effective, climate-smart rice straw management options19, 20. High rates of straw incorporation under aerobic conditions can sequester soil organic carbon (SOC) with a minimal increase in emissions compared to incorporation under flooded conditions. Practices that optimize SOC sequestration while minimizing emissions, such as early straw incorporation with alternate wetting and drying water management, could be an important step towards carbon–neutral rice systems19. Straw incorporation has been shown to enhance nutrient recycling and provide soil fertility benefits by increasing SOC and yields of subsequent crops8, 21,22,23,24. The incorporated straw can also provide substrates to promote biodiversity through flourishing of invertebrates that decompose the straw, which in turn enhances nutrient cycling in paddy soils24, 25. A recent study in Vietnam showed that the addition of rice straw improved soil pH, soil organic C, and nutrient content compared to addition of ash from burned straw26. Due to the low N content in straw, however, large quantities would be needed to supply adequate amounts of N. In addition, the straw has to decompose before the nutrients can become available for uptake and the rate of decomposition and supply of nutrients depend on soil type and season. A sufficient level of decomposition is also needed for mechanized seedbed preparation to prevent clogging of seeders.
Unfortunately, rice straw decomposition takes time. Rice straw is composed mainly of cellulose, hemicellulose, and lignin27, 28. Silica, polysaccharides, and lignin were reported to form complex rigid structures that inhibit the decomposition of straw lignin28,29,30. In a recently reported 5-year experiment, the release of C, N, cellulose, and hemicellulose was found to occur mainly during the first and second years after straw incorporation. Moreover, the release of P and K occurred mainly during the first month, while lignin was released at various rates throughout the entire study period31. Long-term incorporation of crop residues in flooded rice soil can increase soil organic matter, total N, and soil biological activity22. Continuous incorporation of crop residues after each crop can eventually increase the N-supplying capacity of rice soils32. In a study in Vietnam, soil N increased from 0.65 to 0.85% following 9 years of cropping with incorporation of rice straw while straw removal caused a decline in soil N33. However, the benefits of incorporated residues on soil organic matter and soil N supply seldom translate into increased yield or profit for flooded rice5. The production of two or three rice crops annually also results in the production of large quantities of straw, with little turnaround time between crops. This results in limited decomposition of the straw when incorporated, with potential negative effects on nutrient availability and use efficiency of applied fertilizers for the subsequent crop21, 22, 24.
Because of the environmental impacts of straw burning and the importance of quick turnaround times between crops, there is a need to hasten the decomposition of rice straw that is incorporated into the field. Accelerating the decomposition process can promote rice straw incorporation as a preferred practice compared to burning and the application of microbial inoculants could possibly help in doing so.
Microorganisms play a role in the decomposition process as they are the ones responsible for the conversion of crop residues to soil organic matter. Rice straw decomposition is a complex process made up of several subprocesses and involves a diversity of organisms. One of the most common rice straw-degrading microorganisms are the fungi Trichoderma34. Trichoderma spp. play an important role in biological decomposition and are also known to produce cellulolytic and hemicellulolytic enzymes. Trichoderma spp. are filamentous soil fungi that secrete a well-balanced cellulolytic complex, which efficiently hydrolyzes cellulosic substrates into monomeric glucose chain. The potential of Trichoderma spp. in hastening the decomposition of straw has been previously reported35. In the Philippines, the inoculation of a straw-based compost pile with Trichoderma harzianum was reported to have a composting time of less than half of the conventional methods of composting36, 37. Trichoderma spp. are also reported to be able to degrade agricultural and domestic waste relatively quickly without emitting bad odors38. Among the many species of Trichoderma, T. harzianum was reported to produce the highest amounts of cellulolytic enzymes that are useful in degrading straw39. T. viride was also reported to produce ligninolytic enzymes that were able to reduce the lignin content of rice straw40. The use of Trichoderma-based composting technologies has been widely accepted and may have a potential to enhance in situ rice straw decomposition. However, there is limited knowledge on the potential application of Trichoderma spp. to enhance the decomposition of rice straw that is incorporated into the field.
This study aimed to determine if a commercially available Trichoderma-based compost activator, which is usually applied in composing piles, can be used to hasten the decomposition of rice straw that is incorporated into the field.
Materials and methods
Two sets of experiments were performed to determine the potential of a Trichoderma inoculant for in situ rice straw decomposition. The first one is a laboratory incubation experiment that determines the effect of Trichoderma on the decomposition rate of rice straw in sterile and unsterile soils. The second is a field experiment that analyzed the effect of Trichoderma application and straw placement on in situ rice straw decomposition using the litterbag technique. The study complies with local and national guidelines.
Laboratory incubation experiment
The incubation experiment was conducted at the Soil Microbiology Laboratory, Division of Soil Science, Agricultural Systems Institute, College of Agriculture and Food Science of the University of the Philippines Los Baños (UPLB) to determine the effect of Trichoderma on the decomposition of rice straw mixed with sterile and unsterile soil. The experiment was laid in a completely randomized design with five treatments, three replicates per treatment, and nine sampling periods.
Freshly harvested rice straw was collected from the Zeigler Experimental Station (ZES), IRRI, Los Baños, Laguna. It was air-dried and chopped prior to the set-up of the experiment. The treatments included: unsterile soil only (USO), unsterile soil plus rice straw (USRS), unsterile soil plus rice straw plus Trichoderma (USRST), sterile soil plus rice straw (SSRS), and sterile soil plus rice straw plus Trichoderma (SSRST). Fifty grams of soil, 5 g of rice straw, and 500 mg of Trichoderma were used in the designated treatments. The treatments were placed in a wide-mouth incubation jar and mixed thoroughly. Water was added to keep the rice straw moist, but not saturated. A CO2 trap consisting of 30 mL of 0.3 N NaOH was placed and the jar was screwed tightly and incubated at room temperature. Three jars were taken from each treatment at 1, 3, 7, 14, 21, 28, and 42 days after the incubation for the determination of CO2 evolved. The Trichoderma and fungal population were also determined for samples taken at 7, 14, 28, and 42 days after incubation.
Field incubation experiment
The field incubation experiment tested the effect of Trichoderma application and straw placement on the decomposition of rice straw under field conditions. The experiment was laid out in a completely randomized design with two factors, four treatments with three replicates per treatment, for five sampling periods. The two factors considered are presence or absence of the Trichoderma activator and placement of the litterbag (soil surface or below ground). Hence, the treatments are: no Trichoderma on soil surface (NTSS), no Trichoderma below ground (NTBG), with Trichoderma on soil surface (WTSS), and with Trichoderma below ground (WTBG).
The field experiment used the litter bag technique in analyzing the effect of treatments on rice straw decomposition. To perform this, freshly harvested rice straw was collected from the ZES. A total of 100 g of air-dried and chopped rice straw were placed in nylon bags with a mesh size of 20 × 20 µm each. The mesh size selected for this experiment was very small to ensure that soil animals did not enter the litterbag and affect the decomposition as previously reported25. One gram of Trichoderma activator was added to the litter bags with the Trichoderma treatment. A total of 96 litterbags were placed in the field, half of which were inoculated with Trichoderma. One set of the treatments was placed at the soil surface secured with steel wire while the other set was buried 10 cm below ground. Six litterbags per treatment were sampled at 7, 14, 21, 28 and 42 days after the field set-up. One set of three replicates was subjected to weight loss determination, while the other three replicates were used for the determination of fungal populations (Trichoderma, other fungi, and total fungal populations) and analysis for total carbon and total nitrogen.
Data collection
The analyses performed on the rice straw and soil samples are discussed in detail below.
Determination of CO2 evolved using the titration method
The contents of the CO2 trap were transferred to a 125-mL Erlenmeyer flask. Two to three drops of phenolphthalein indicator and 1.0 mL of 50% BaCl2 were added and titration was performed using 0.2 N HCl until the pink color just disappeared. The amount of acid used to neutralize the base was recorded. Likewise, 30 mL of a fresh sample of 0.3 N NaOH were titrated. The amount of CO2 evolved per 100 g rice straw was calculated using the following formula:
where:
V = Volume (mL) of acid used to titrate the alkali in the CO2 trap to the end point.
B = Volume (mL) of acid used to titrate the fresh sample of alkali to the end point.
N = Normality of the acid.
22 = Weight of 1 meq CO2 in mg.
Determination of fungal populations
The fungal populations (Trichoderma, other fungi, and total fungal population) were determined using the spread-plate method. Ten grams of sample from the incubation experiment were placed in 95 mL of sterile distilled water and were serially diluted. One hundred microliters of liquid from dilutions 10–3 to 10–5 were spread-plated on Potato Dextrose Agar (PDA) with Rose Bengal supplemented with 50 mg L−1 streptomycin. Potato dextrose agar (PDA) is an enrichment medium that enables the growth of all fungal species present from the sample. Trichoderma colonies can be differentiated from other fungi because they have irregular shape, with cottony and yellowish green conidia with white mycelia towards the margin41. They also have a yellow irregular zone surrounding the colony at the bottom side of the glass plates. Similarly, 10 g of sample were oven-dried at 105 °C for 72 h. The colonies that formed were counted and the populations were calculated using the formula:
Determination of total C and total N (combustion method)
Rice straw samples were ground twice using Wiley and Ball Mill grinders until the straw had the desired talc-like consistency. A small amount (1.5 to 2.0 g) of the powdered samples was submitted to the IRRI Analytical Service Laboratory for total C analysis using a dynamic combustion system coupled with a gas chromatograph equipped with a thermal conductivity detector (GC-TCD).
Determination of weight loss and decomposition rate
The retrieved litterbags were separated from the adhering soil particles by washing them carefully with tap water and then pat dried using a clean cloth. The straw was removed from the litterbags and weighed to determine its fresh weight. Next, the straw was oven-dried at 70 °C for 72 h and then weighed again for its oven-dried weight. The decomposition rate of the straw was determined based on mass loss using the exponential decay model42:
where:
k = rate of decomposition.
Mo = initial dry mass of rice straw.
Mt = dry mass of rice straw after t time.
t = time (days) the straw is in the soil.
Statistical analysis
The analysis of variance (ANOVA) for completely randomized design (CRD) was performed on the collected data per sampling period. Least Significant Difference (LSD) was used in comparing the means of all parameters measured using the Statistical Tool for Agricultural Research (STAR) software.
Results
Characterization of the soil and environment
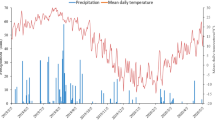
ZES at IRRI has 200 ha of research fields dedicated to different rice research activities. One of the fields (plot UJ) was selected as the study site due to its availability during the time of the study. The site was dried during its fallow period and was previously planted with lowland rice. Plot UJ is situated at 14° 08′ 40.48″ N and 121° 15′ 53.21. Soil samples collected from plot UJ were found to have a loam texture, with 23.33% clay, 43.33% silt, and 33.33% sand. The soil also has a neutral pH of 7.04. The laboratory incubation experiment was exposed at an ambient temperature of around 27 °C. The average daily temperature during the field incubation was a bit higher and ranged from 29.1 to 29.6 °C (Fig. 1). Higher precipitation was observed during the start of the field incubation experiment, with daily rainfall of around 20 to 30 mm. Lesser amounts of rainfall were observed at the end of the first week, while the second week of incubation was characterized by higher precipitation at the later part, around 11 to 14 days. The highest precipitation of almost 50 mm was observed on the first day of the third week, but no precipitation was observed afterwards until the last few days of the fourth week. This was followed by steady precipitation until the last sampling (Fig. 1).

Average daily rainfall and temperature in the field during the experiment period.
Fungal population dynamics in decomposing straw under laboratory and field conditions
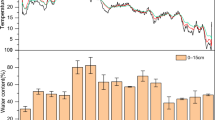
Under both laboratory and field conditions, Trichoderma was only observed in inoculated rice straw (Fig. 2A and B). This indicated that the inoculation of Trichoderma was successful and that the Trichoderma did not contaminate the uninoculated samples. Under laboratory conditions, no fungal population was detected in the unsterile soil sample (USO treatment). This indicates that the fungal population observed mainly comes from the rice straw and Trichoderma that was added to the rice straw-soil mixture. From a population of a 4 × 104 CFU/g sample after the first week of incubation, the Trichoderma population in the USRST treatment was doubled to 8 × 104 after 14 days. On the other hand, the Trichoderma population in SSRST treatment was reduced to 3.4 × 104. The Trichoderma population declined at days 28 and 42, with the USRST consistently having the highest Trichoderma population. Populations of other fungi were lower in the Trichoderma-treated samples than the uninoculated counterparts during the first 4 weeks (7, 14, and 28 days) of incubation. A significant effect of the inoculation was found at day 14, in which the other fungal populations in the USRS were 8.12 × 104 while USRST had 3.52 × 104. The obtained values indicated a 56.7% reduction of fungal populations brought about by Trichoderma inoculation. The same was observed in sterile samples in which SSRS had 9.3 × 104 while USRST had a population of 4.32 × 104 (53.55%). Further reduction in population was observed at day 28, but the reduction of the populations of other fungi was reduced to 6.75 and 46.98% in USRST and SSRST, respectively. Trichoderma was found to dominate the inoculated samples during the first 4 weeks of incubation. The fungal populations of both the USRST and SSRST treatments had around 40% Trichoderma after 1 week of incubation. At day 14, a spike in the Trichoderma population was observed in USRST, wherein 70% of the total fungal population consisted of Trichoderma. SSRST, on the other hand, maintained the 40% Trichoderma population until day 28. At day 28, the Trichoderma population in USRST was reduced to around 40% of the total fungal population. The Trichoderma population in both treatments was further reduced at day 42. The reduction in the Trichoderma population was simultaneous with the increase in the population of other types of fungi. Although inoculated treatments had lower populations of other fungi at the initial stages of the experiment, Trichoderma-treated samples had a higher total and other fungal populations compared to the uninoculated counterpart at 42 days. Populations of other fungi in USRST at day 42 were 6.13 × 104, 185% higher than USRS, which had other fungal populations of only 2.15 × 104.

Fungal population dynamics in the (A) laboratory and (B) field incubation experiments. USO = unsterile soil only; USRS = unsterile soil plus rice straw; USRST = unsterile soil plus rice straw plus Trichoderma; SSRS = sterile soil plus rice straw; SSRST = sterile soil plus rice straw plus Trichoderma; NTSS = no Trichoderma on soil surface; NTBG = no Trichoderma below ground; WTSS = with Trichoderma on soil surface; WTBG= with Trichoderma below ground.
The increase in total fungal population reached up to 300%, with USRS having 2.15 × 104 and USRST having 8.75 × 104. Similar results were obtained in sterile soil samples, with 87 and 115% increases in other fungi and total fungal populations, respectively.
The trends observed in the laboratory experiment were also observed in the field. The Trichoderma population increased at days 14, 21, and 28, and eventually declined at day 42. A significant effect of placement was observed wherein buried litterbags had significantly higher Trichoderma and total fungal population compared to the litterbags placed on the surface (Table 1; Fig. 2B). The Trichoderma population in WTBG increased from 4.03 × 104 at day 7 to 9.78 × 104 at day 14 (a 142% increase) and further increased to 1.80 × 105 at day 21, which was the highest observed throughout the incubation period. The Trichoderma population slightly declined on day 28, with a population of 1.5 × 105 and further after 42 days, with a population of only 3.03 × 104. The same trend was observed with the indigenous fungal populations, in which inoculated samples had lower indigenous fungal populations compared with the uninoculated counterpart. The effect, however, was not as drastic as the effect observed in the laboratory. A reduction of 29.30% (3.49 × 105 in NTBG, 2.46 × 105 in WTBG) and 26.69% (2.45 × 105 in NTBG, 1.89 × 105 in WTBG) was observed at days 14 and 21, respectively. The effect of placement on the populations of other fungi was also significant at day 21, with the litterbags placed below ground having higher other fungal populations (Table 1). At day 42, the other fungal populations in WTBG (9.98 × 104) were slightly higher than that of NTBG (8.35 × 104) but was not significantly different. The total fungal population (Trichoderma + other fungi) was significantly affected by the placement method at days 14, 21, and 28 (Table 1). Higher total fungal population was consistently observed in the WTBG treatments where the litterbags were inoculated with Trichoderma and placed belowground (Fig. 2B).
Observation of decomposition through CO2 evolved from the decomposing rice straw in the laboratory experiment
Because the liberation of CO2 occurs during decomposition, CO2 production is a widely accepted quantitative expression of the decay process43. In the laboratory experiment, it was observed that treatments inoculated with Trichoderma, USRST and SSRST, consistently released higher amounts of CO2 compared to their uninoculated counterparts, USRS and SSRS, respectively (Table 2). The highest amount of CO2 evolved was observed in the SSRST treatment with sterile soil and Trichoderma inoculant at days 1, 3, and 7. The SSRST treatment consistently had significantly higher amounts of CO2 evolved compared with SSRS during the first 4 weeks of incubation (p < 0.05). At day 1, SSRST released 201.23 mg CO2. This is 35% compared to SSRS, which released only 149 mg CO2. In the succeeding sampling periods, SSRST consistently released higher amounts of CO2, around 16% higher than SSRS on the average. The effect of Trichoderma inoculation was more pronounced in sterile soil samples than unsterile soil samples for the first 4 weeks of incubation. A significant effect on soil condition and inoculation was observed at day 21, in which USRST (unsterile soil + rice straw + Trichoderma) released the highest amount of CO2. At day 42, simultaneous to the decline in fungal population, the effect of inoculation on CO2 production was no longer significant. Having the highest total fungal population at the end of the 42-day incubation, the USRST treatment (unsterile soil + rice straw + Trichoderma inoculant) also had the highest amount of CO2 evolved at 2943.16 mg CO2, followed by USRS (unsterile soil without Trichoderma) with a value of 2763.64 mg CO2 and SSRST (sterile soil + rice straw + Trichoderma inoculant), which released a total of 2,405.04 mg CO2.
Field experiment
Effect of Trichoderma inoculation and placement method on the percent weight loss and decomposition rate (k) of rice straw
While Trichoderma inoculation resulted in higher decomposition rates in the laboratory experiment, the field experiment seems to show otherwise. Lower percent mass loss and decomposition rates were observed in treatments inoculated with Trichoderma (WTSS and WTBG) compared to those without Trichoderma (NTSS and NTBG). At day 21, when the highest Trichoderma population was observed in the WTBG treatment, the mass loss in NTSS and NTBG was significantly higher at 32.20 and 37.67%, respectively, while inoculated treatments WTSS and WTBG only resulted in 20.06 and 28.29% mass losses, respectively. The highest percent mass loss value of 38.99% was observed in the NTBG treatment at day 28, while inoculated samples WTSS and WTBG were significantly lower at only 23.69 and 24.01%, respectively. At day 42, a decrease in mass loss was observed in NTBG (37.83%) while the mass loss in WTBG increased to 31.23%. At the end of the 6-week incubation, the percent mass loss in WTBG was comparable with NTSS and NTBG.
Effect of Trichoderma inoculation and straw placement on C and N contents and C:N ratio
No significant difference was observed in the C content of litterbags during the first 3 weeks of incubation. Significant effects of placement and Trichoderma application were observed at days 28 and 42, in which the WTBG treatment had the lowest C content of 29.77% compared to 34.30% in NTSS, 31.10% in NTBG, and 33.83% in WTSS. The N content was not significantly different among the treatments during the first 4 weeks of incubation. At day 42, WTBG had the highest N content of 1.1%, resulting in the lowest C:N ratio compared to the other treatments. The C:N ratio of WTBG was only 26.27—39.51% lower than the C:N ratio of NTBG, which was 43.43.
Discussion
Effect of inoculation on Trichoderma and total fungal population in the field and laboratory
For both the laboratory and field experiments, the Trichoderma population was significantly higher in inoculated samples (p < 0.01), indicating that no contamination occurred during the course of the study. Since the environmental conditions in the laboratory, such as temperature and moisture, were constant, the fungal population dynamics observed is a reflection of the microbial interactions that occurred in the samples. At the start of the laboratory incubation, there was a significant reduction of the indigenous fungal populations in inoculated samples. Trichoderma spp. are active ingredients in many commercial bio-fungicides used to control a range of economically important fungal plant pathogens44. Trichoderma was reported to control soilborne pathogens, which are mostly fungi, such as Fusarium, Rhizoctonia, and Pythium45. The antagonistic activity of the Trichoderma strains is attributable to complex mechanisms, including the activity of cell wall-degrading enzymes, direct competition for nutrients, antibiosis, and the induction of plant defense responses at both the local and systemic levels46. Trichoderma attaches to the host fungi and then coils its hyphae around the host, forms appressoria on the host surface, penetrates the host cell, and collapses the host hyphae47. The inoculant applied may have used the indigenous fungal populations as hosts and this resulted in the increase in the Trichoderma population and simultaneous reduction in the populations of other fungi.
Fungal succession was reported to be related to substrate quality This has been well documented in studies on decomposition48,49,50,51,52. In the laboratory incubation experiment performed in this study, limited amounts of substrates are available because rice straw is highly cellulolytic and ligninolytic. In cases like this, extracellular enzymes, such as cellulases, play a key role in decomposing complex organic polymers into oligomeric and monomeric substances that are small enough to be transported inside the cell43, 53. Because of the known ability of Trichoderma to produce high amounts of cellulolytic and hemicellulolytic enzymes, it can be assumed that inoculated samples have higher amounts of cellulases compared to uninoculated ones. This will enhance the hydrolysis of the cellulosic substrates in the rice straw and eventually lead to the production of higher amounts of monomeric glucose units that can be taken up by the indigenous fungal populations. The increased amount and quality of substrates available in inoculated samples can lead to the promotion of microbial growth. This was reflected during the last part of the experiment wherein the total fungal population was significantly higher in inoculated samples (p < 0.05). Similarly, the continuous decline of the fungal populations in uninoculated samples can be attributed to the depletion of the readily available substrates because the microorganisms present are not capable of producing cellulolytic enzymes that can degrade the complex cellulosic substrates.
Independent and combined effects of Trichoderma inoculation and the method of straw placement on the fungal populations were observed during the last 2 weeks of field incubation. A higher Trichoderma population was observed in inoculated samples that were placed below ground than those placed on the surface (Table 1, p < 0.01). The significantly higher Trichoderma in the WTBG over the WTSS could be explained by the difference in soil moisture and the degree of contact between rice straw and soil as affected by the method of straw placement. Buried litterbags are expected to have higher moisture since the moisture of surface-applied litterbags may dry up easily during warm days54. This is consistent with previous research that reported greater fungal colonization in litterbags with intimate litter-soil contact55. A consistent trend was observed between the laboratory and field experiments, in which the Trichoderma population starts to decline at the fourth week of incubation. It is interesting to note that, while the indigenous fungal populations initially declined in inoculated samples during the first 2 weeks of incubation, inoculated samples were the ones with a higher total fungal population at the end of the incubation period. The total fungal population in the USRST (8.75 × 104) was 300% higher than the USRS (2.15 × 104), while the total fungal population in the WTBG (1.30 × 105) was 55.66% higher than the NTBG (8.35 × 104).
Effect of Trichoderma inoculation on CO2 evolution
The overall process of decomposition generally involves a broad spectrum of complementary microbes that act in concert on a substrate43. These microorganisms produce different kinds of enzymes that act on the organic compounds present. Soil microbes use residue components as substrates for energy and as carbon sources in the synthesis of new cells. Energy is furnished to the microbial cells through the oxidation of the organic compounds and a major product is CO2, which is released back into the atmosphere. Microorganisms are generally short-lived and themselves are decomposed by successive populations that find the dying cells to be more suitable substrates than the initial residues43. Given this principle, we can assume that the C present in the rice straw has three faiths: (1) released as CO2; (2) converted into microbial biomass, whether bacterial or fungal; or (3) remaining as an undecomposed component of the straw.
In the laboratory experiment, the use of sterile or unsterile soil was intentionally done to consider the influence of the soil’s indigenous microbial populations on rice straw decomposition. The SSRST treatment released the highest amount of CO2 during the first week of incubation. The amount released was 35% more compared to its uninoculated counterpart (SSRS) on day 1, suggesting that Trichoderma contributed greatly to the decomposition of the substances present in the soil. The high amounts of CO2 released during day 1 were an indication that most of the C present was being used by the microorganisms (composed mainly of Trichoderma) for respiration to produce energy that will be used for metabolic activities during the period of adjustment to the new environment (the incubation jar). The contribution of Trichoderma to CO2 evolution lowered to around 16% in the succeeding days, but the difference in CO2 evolved between inoculated (SSRST) and uninoculated (SSRS) samples were still significantly different until day 28 (Table 2).
At this point, we can assume that some of the C was now being used as part of the microbial biomass, especially since Trichoderma produces hyphae that branch into thick mycelia. Microbial efficiency is the term used for the percentage of C that is being converted into microbial biomass43. In unsterile soil samples, the amount of CO2 evolved in inoculated samples (USRST) was also higher than the CO2 evolved in uninoculated ones during the first 2 weeks of incubation. The difference, however, was not significant and this was possibly caused by the indigenous microorganisms that are present in the unsterile soil. While some groups of microorganisms are metabolizing and releasing CO2, another group may be using the C to build microbial biomass. Microbial interactions are also most likely to occur, and this complex process may have influenced the release of CO2. A significant effect of inoculation on CO2 evolution was observed on day 21. From day 14 onwards, the USRST treatment (unsterile soil + Trichoderma inoculant) had the highest amount of CO2 evolved among all the samples. This was followed by SSRST (sterile soil with Trichoderma) and USRS (unsterile soil without Trichoderma) treatments. These observations indicate that the Trichoderma inoculant played a big role during the first 3 weeks of incubation.
Effect of Trichoderma inoculation on % mass loss and decomposition rate
Trichoderma has been reported to be capable of degrading straw and other organic materials38, 40, 56,57,58,59,60. The success in using Trichoderma for composting has been well-documented. The inoculation of straw-based compost piles with Trichoderma harzianum was reported to have a composting time of less than half of the conventional methods of composting36, 37. Rice straw treated with Trichoderma harzianum and cow dung slurry was reported to increase the population of beneficial microorganisms and improved crop yield61. The decomposition of empty fruit bunches (EFB) and palm oil mill effluent (POME) was also reported to be reduced from 4 to 6 months to 21–45 days62. In vitro and in vivo conversion of solid organic waste was also reported possible using Trichoderma59. Commercially available Trichoderma-based inoculants are currently available in the market and most of these products are successfully used in composting.
The results of the laboratory experiment in this research supported the previous claims where inoculated samples release higher amounts of CO2. In the field experiment, however, rice straw samples inoculated with Trichoderma were the ones with the low percent mass loss and decomposition rate throughout the incubation period. The highest percent mass loss value of 38.99% was observed in NTBG treatment at day 28, while inoculated samples WTSS and WTBG were significantly lower at only 23.69 and 24.01%, respectively. While most research considers mass loss as an indicator of decomposition, this may not be applicable in this study because of the proliferation of fungi in the inoculated litterbags, which resulted in an increase in biomass. Recall that the WTBG treatment had the highest Trichoderma and total fungal population with sampling at both days 21 and 28. During these sampling days, the percent mass loss values of Trichoderma-treated samples were significantly lower than the uninoculated ones, possibly because of the increased biomass from the high fungal population. At day 42, simultaneous to the decrease in total fungal population in WTBG, the percent mass loss increased to 31.23% and was no longer significantly different with NTBG, which had a mass loss of 37.83%. Since the increase in mass brought about by the growth of Trichoderma and other fungi was not accounted for, the measurement of decomposition rate through mass loss and exponential decay model cannot accurately represented the rate of decomposition that occurred. A previous study reported increased mass loss in buried rice straw with the use of Trichoderma and frequent watering58. However, upon checking, that research only used 5 g of rice straw, which is 20 times smaller than the initial amount of straw used in this experiment. The previous research also mentioned that watering daily resulted in increased mass loss, but further details were provided in the methodology. Fungal biomass varies widely within and across biomes in relation to litter composition, root density, and nutrient availability. Fungi may comprise of up to 20% of the mass of decomposing plant litter and the fungal mycelia grow radially as fractal networks in soil, wood, and litter63, 64 Since determining the fungal biomass was not done in this study, we cannot be sure of the actual proportion of biomass from the fungi and from the rice straw.
Effect of Trichoderma inoculation on C and N content and C:N ratio of decomposing rice straw
As mentioned above, C present in the rice straw has three faiths: (1) released as CO2; (2) converted into microbial biomass, whether bacteria or fungi; or (3) remaining as an undecomposed component of the straw. Since the CO2 is released back into the atmosphere, the C present in the litterbags represented C from both the microbial biomass and undecomposed rice straw component. At the start of the experiment, the C content in all the treatments was 34 or 35%. During this period, the total fungal population was also similar among the treatments. While the mass gain brought about by the increased Trichoderma and total fungal population cannot be determined, a comparison of the fungal population and C content of the WTBG and NTBG treatments can help explain the relevance of high fungal population on the mass and C content of the samples. After 3 weeks of incubation, the C content of uninoculated samples, NTBG and NTSS, was lower than the initial value by about 2%, while the C content of WTBG was still close to the initial value (Table 3). During the same period, the WTBG treatment had the highest Trichoderma population among the treatments, (p < 0.01). Since the other treatments did not have a significantly high Trichoderma and total fungal population, we can assume that the C in the NTBG, NTSS, and WTSS was mainly in the form of undecomposed rice straw. On the other hand, we can also assume that a significant amount of C measured in WTBG treatment came from both the undecomposed straw and the fungal biomass present in the litterbags. At day 28, despite having the highest total fungal population, the C content in the WTBG treatment decreased to 29% from an initial value of 34.5%. This was significantly lower compared with NTBG (31.1% C), WTSS (33.83% C), and NTSS (34.30% C) (p < 0.05). Since the total fungal population in the WTBG remained the same from days 21 to 28, the reduction in C content can be attributed solely to rice straw decomposition. At day 42, the fungal population between NTBG and WTBG were not significantly different, while the C content in the WTBG was down to 28.63%, which is the lowest C content among the other samples (Table 3). This indicates that the difference in C content between the treatments was mainly due to the degradation of the rice straw. Differences in N content were also observed among the samples. A significant effect was observed on day 42, with the WTBG treatment having the highest N content at 1.1%.
Aside from determining weight loss, other researchers have used the C:N ratio as an indicator of decomposition rate65, 66. The C:N ratio of the plant litter is an indicator of the speed of decomposition and mineralization in soils. Crop residues with a narrow C:N ratio decompose faster than crop residues with a wide C:N ratio67. Crop residues with a high C:N ratio are also considered to be of low quality, whereas a low C:N ratio results in high-quality materials with a faster decomposition68. This is because residues with a low C:N ratio exhibit net N mineralization, while residues with a high C:N ratio exhibit immobilization69,70,71 In farming systems where the straw remains on the field after harvest, rapid decomposition is important to minimize negative effects caused by N immobilization on the following crops72, 73. The availability of other nutrients is also affected by the low quality of the rice straw, with a high C:N ratio, resulting in slow decomposition and mineralization of nutrients, particularly short-term availability of N and, to some extent, P71. The C:N ratio can be a reliable basis for measuring the decomposition of rice straw instead of only considering the results based on mass loss.
A computed C:N ratio of the plant litter is a better indicator of the decomposition rate since the values are expressed on a dry weight basis and the fluctuation in the values can be related to the Trichoderma population. In addition, it can indicate whether N will be mineralized or immobilized for the use by the succeeding crop. Rice straw is mostly made up of cellulose, resulting in its high C:N ratio and slow decomposition rate. A previous report mentioned that yield depression following straw incorporation can be mitigated by adding inorganic N74. In this study, the WTBG treatment had the lowest C/N Ratio at day 42, which was 26.27, compared with the NTBG, which had a high C/N ratio of 43.43. The C/N ratio of the WTBG was 39.53% lower than its uninoculated counterpart, even though no inorganic N had been added to the incorporated straw. If the fallow period of a farm is only 8 weeks, it is likely that the crops planted in areas with buried straw inoculated with Trichoderma will exhibit mineralization, while the uninoculated straws will show signs of immobilization.
Conclusions
This study assessed the potential of a Trichoderma-based compost activator for the enhanced decomposition of incorporated rice straw by conducting experiments under both laboratory and field conditions. Results of the laboratory experiment indicated that the inoculant could increase decomposition rates under laboratory conditions, with an average of 16% higher amounts of CO2 released compared to uninoculated straw in sterile soil samples. In the field experiment, improved decomposition was observed in samples inoculated with Trichoderma and placed below ground (WTBG). From the initial value of around 35%, the C content in the WTBG was down to 28.63% after 42 days of incubation and was the lowest among treatments. The WTBG treatment also had the highest N content of 1.1%. The C:N ratio of the WTBG was only 26.27. This was 39.51% lower than the C:N ratio of the NTBG (no Trichoderma placed below ground), which was 43.43. At the end of the 6-week incubation period, the straw that was buried and inoculated with Trichoderma had the highest total fungal population, highest N content, lowest C content, and lowest C:N ratio, which are the conditions of an effective decomposition. These results confirmed that the Trichoderma-based compost activator can hasten the decomposition of incorporated rice straw. The increased fungal population in inoculated samples is an indication of enhanced biodiversity and the low C:N ratio promotes N mineralization.
While the improved chemical and biological properties in the incorporated straw indicated better decomposition, the changes in the physical conditions of the soil were not assessed in this study. Additional experiments may be needed to determine such properties since this information would be helpful in the field operations for the succeeding cropping period, which includes land and seedbed preparation. The use of additional amendments, such as manure and/or organic fertilizers or performing additional field operation activities, may also be tested to determine if the decomposition period can be shortened further.
Change history
25 January 2022
A Correction to this paper has been published: https://doi.org/10.1038/s41598-022-05490-7
References
IRRI. World Rice Statistics, http://ricestat.irri.org:8080/wrsv3/entrypoint.htm (2021).
Gadde, B., Bonnet, S., Menke, C. & Garivait, S. Air pollutant emissions from rice straw open-field burning in India, Thailand, and the Philippines. Environ. Pollut. 157(5), 1554–1558 (2009).
Mendoza, T.C. & Samson, R. Strategies to avoid crop residue burning in the Philippine context. Resource Efficient Agricultural Production (REAP). 1–18; http://reap-canada.com/library/IntDev (1999).
Kutcher, H. R. & Malhi, S. S. Residue burning and tillage effects on diseases and yield of Barley (Hordeum vulgare) and canola (Brassica napus). Soil Till. Res. 109(2), 153–160 (2010).
Singh, B., Shan, Y. H., Johnson-Beebout, S. E., Singh, Y. & Buresh, R. J. Crop residue management for lowland rice-based cropping systems in Asia. Adv. Agron. 98, 177–199 (2008).
Dobermann, A. & Fairhurst, T. H. Rice straw management. Better Crops Int. 16(1), 7–11 (2002).
Raison, R. J. Modification of the soil environment by vegetation fires, with particular reference to nitrogen transformations: A review. Plant Soil 51, 73–108 (1979).
Nigussie, A. & Kissi, E. Impact of biomass burning on selected physicochemical properties of nitisol in Jimma zone, Southwestern Ethiopia. Int. Res. J. Agric. Sci. Soil Sci. 1, 394–401 (2011).
Ponnamperuma, F. Straw as a source of nutrients for wetland rice. Organic Matter Rice. 117–136 (IRRI,1984).
Lefroy, R., Chaitep, W. & Blair, G. Release of sulfur from rice residues under flooded and non-flooded soil conditions. Aust J. Agric. Res. 45, 657–667 (1994).
Romasanta, R. R. et al. How does burning of rice straw affect CH4 and N2O emissions? A comparative experiment of different on-field straw management practices. Agric. Ecosyst. Environ. 239, 143–153 (2017).
Launio, C.C., Asis, C.A., Manalili, R.G., Javier, E.F. Economic Analysis of Rice Straw Management Alternatives and Understanding Farmers’ Choices. Cost-Benefit Studies of Natural Resource Management in Southeast Asia, Springer, 93–111 (2013).
Akagi, S. K. et al. Emission factors for open and domestic biomass burning for use in atmospheric models. Atmos. Chem. Phys. 11(9), 4039–4072 (2011).
Zhang, G. B., Ji, Y., Ma, J., Xu, H. & Cai, Z. C. Case study on effects of water management and rice straw incorporation in rice fields on production oxidation, and emission of methane during fallow and following rice seasons. Soil Res. 48, 238–246 (2011).
Nguyen, V. H. et al. An assessment of irrigated rice production energy efficiency and environmental footprint with in-field and off-field rice straw management practices. Sci. Rep. 9, 16887 (2019).
Gummert, M., Quilty, J., Hung, N.V. & Vial L. Engineering and management of rice harvesting in Advances in Science & Engineering of Rice (ed. Pan, Z & Khir, R). (2018)
Fei, L. How can straw incorporation management impact on soil carbon storage? A meta-analysis. Mitigat. Adapt. Strategies Global Change Springer 20(8), 1545–1568 (2015).
Liu, C., Lu, M., Cui, J., Li, B. & Fang, C. Effects of straw carbon input on carbon dynamics in agricultural soils: A meta-analysis. Glob Chang Bio 20, 1366–1381 (2014).
Allen, J. et al. Rice straw management effects on greenhouse gas emissions and mitigation options in Sustainable Rice Straw Management (ed. Gummert M., Hung N., Chivenge P. & Douthwaite B.) 145–159 (Springer, 2020).
Launio, C. C., Asis, C. A., Manalili, R. G. & Javier, E. F. Cost-effectiveness analysis of farmers’ rice straw management practices considering CH4 and N2O emissions. J. Environ. Manage. 183, 245–252 (2016).
Dobermann, A. & Fairhurst T.H. Rice: Nutrient Disorders and Nutrient Management. (International Rice Research Institute, 2000).
Yadvinder, S. et al. Effects of residue decomposition on productivity and soil fertility in rice–wheat rotation. Soil Sci. Soc. Am. J. 68(3), 854–864 (2004).
Gupta, R., Ladha, J., Singh, J., Singh, G. & Pathak, H. Yield and phosphorus transformations in a rice–wheat system with crop residue and phosphorus management. Soil Sci. Soc. Am. J. 71(5), 1500–1507 (2007).
Chivenge, P. et al. Rice Straw Incorporation Influences Nutrient Cycling and Soil Organic Matter in Sustainable Rice Straw Management (ed. Gummert M., Hung N., Chivenge P. & Douthwaite B.) 131–144 (Springer, 2020).
Schmidt, A. et al. Effects of residue management on decomposition in irrigated rice fields are not related to changes in the decomposer community. PLoS ONE. 10(7), (2015).
Thanh, N.D., Hoa, H.T.T., Hung, H.C., Nhi, P.T.P. & Thuc, D.D. Effect of fertilizer on rice yield improvement in coastal sandy soil of Thua Thien Hue province. Hue Univ. J. Sci. Agric. Rural Dev. 119, (2016).
Pan, M., Gan, X., Mei, C. & Liang, Y. Structural analysis and transformation of biosilica during lignocellulose fractionation of rice straw. J. Mol. Struct. 1127, 575–582 (2017).
Wu, W., Wang, Z., Jin, Y., Matsumoto, Y. & Zhai, H. Effects of LiCl/DMSO dissolution and enzymatic hydrolysis on the chemical composition and lignin structure of rice straw. Biomass Bioenergy. 71, 357–362 (2014).
Eremeeva, T., Bikova, T., Eisimonte, M., Viesturs, U. & Treimanis, A. Fractionation and molecular characteristics of cellulose during enzymatic hydrolysis. Cellulose 8, 69–79 (2001).
Pan, M., Zhou, D., Zhou, X. & Lian, Z. Improvement of straw surface characteristics via thermomechanical and chemical treatments. Biores Technol. 101, 7930–7934 (2010).
Yan, C. et al. Decomposition characteristics of rice straw returned to the soil in northeast China. Nutr. Cycl. Agroecosyst. 114, 211–224 (2019).
Eagle, A. J. et al. Rice yield and nitrogen utilization efficiency under alternative straw management practices. Agron J. 92(6), 1096–1103 (2000).
Thuan, H. N. & Long, D. T. Use rice straw from previous season for the following season on degraded soil, Bac Giang province Vietnam. J. Sci. Dev. Hanoi Agric. Univ. 8, 843–849 (2010).
Hudson, H.J. The development of the saprophytic fungal flora as leaves senesce and fall in Ecology of Leaf Surface Microorganisms (ed. Preece, T.F., Dickinson, C.H.) 431–445 (University of Michigan Academic Press, 1971).
Harper, S.H.T. & Lynch, J.M. Colonization and decomposition of straw by fungi. Trans. Br. Mycol. Soc. 85(4), (1985).
Cuevas, V. C., Samulde, S. N. & Pajaro, P. Trichoderma harzianum Riifai as an activator for rapid composting of agricultural wastes. Philipp. Agric. 71(4), 461–469 (1988).
Cuevas, V. C. Rapid composting technology in the Philippines: Its role in producing good-quality organic fertilizers. Food Fertilizer Technology. Center Extn. Bull. 444, 1–13 (1997).
Ilias, G. N. M., Rahman, M. A. & Molla, A. H. Composting of municipal garbage by using Trichoderma—a new approach in context of Bangladesh. Bangladesh J. Genet. Biotech. 6(1 & 2), 75–78 (2005).
Lee, S. Rice straw-decomposing fungi and their cellulolytic and xylanolytic enzymes. J. Microbiol. Biotechnol. 21(12), 1322–1329 (2011).
Ghorbani, F., Karimi, M., Biria, D., Kariminia, H. R. & Jeihanipour, A. Enhancement of fungal delignification of rice straw by Trichoderma viride sp. to improve its saccharification. Biochem. Eng. J. 101, 77–84 (2015).
Shah, S., Nasreen, S. & Sheikh, P. A. Cultural and morphological characterization of Trichoderma spp. associated with green mold disease of Pleurotus spp. in Kashmir. Res. J. Microbiol. 7, 139–144 (2012).
Graca, M., Barlocher, F. & Gessner, M. Methods to study litter decomposition: A practical guide. Preprint at www.springer.com/gp/book/9781402033483 (2005).
Wagner, G.H. & Wolf, D.C. Carbon transformations and soil organic matter formation in Principles and Applications of Soil Microbiology (ed. Sylvia, D.M., J.J. Fuhrmann, P.G. Hartel, D.A. Zuberer) 218–258 (Prentice Hall, Inc., 1999).
Harman, G. E. Myths and dogmas of biocontrol: changes in perceptions derived from research on Trichoderma harzianum T-22. Plant Dis. 84, 377–393 (2000).
Mohiddin, F. A., Khan, M. R., Khan, S. M. & Bhat, B. H. Why Trichoderma is considered super hero (Super Fungus) against the evil parasites?. Plant Pathol. J. 9, 92–102 (2010).
Harman, G. E., Howell, C. R., Viterbo, A., Chet, I. & Lorito, M. Trichoderma species—opportunistic, avirulent plant symbionts. Nat. Rev. Microbiol. 2, 43–56 (2004).
Steyaert, J. M., Ridgway, H. J., Elad, Y. & Stewart, A. Genetic basis of mycoparasitism: A mechanism of biological control by species of Trichoderma. J. Crop. Horticul. Sci. 31, 281–291 (2003).
Deacon, J. W. Fungal Biology, 4th ed., Preprint at https://www.wiley.com/en-us/Fungal+Biology%2C+4th+Edition-p-9781405130660 (2005). http://dspace.fudutsinma.edu.ng/jspui/bitstream/123456789/1808/1/Fungal Biology.pdf
Finlay, R.D. & Rosling, A. Integrated nutrient cycles in forest ecosystems: The role of ectomycorrhizal fungi in Fungi in Biogeochemical Cycles (ed Gadd, G.M.) Cambridge University Press, 2006).
Leake, J. R. Plants parasitic on fungi: Unearthing the fungi in myco- heterotrophs and debunking the “saprophytic” plant myth. Mycologist 19, 113–122 (2005).
Lindahl, B.D., Finlay, R.D. & Cairney, J.W.G. Enzymatic activities of mycelia in mycorrhizal fungal communities in The Fungal Community: Its Organization and Role in the Ecosystem (ed. Dighton, J., Oudemans, P. & White, J.) 331–348 (New York, 2005).
Lindahl, B. D. & Finlay, R. D. Activities of chitinolytic enzymes during primary and secondary colonisation of wood-by-wood degrading basidiomycetes. New Phytol. 169, 389–397 (2006).
Sinsabaugh, R. L. & Follstad Shah, J. J. Ecoenzymatic stoichiometry of recalcitrant organic matter decomposition: the growth rate hypothesis in reverse. Biogeochemistry 102, 31–43 (2011).
Abera, G., Meskel, E. W. & Bakken, L. R. Unexpected high decomposition of legume residues in dry season soils from tropical coffee plantations and crop lands. Agron. Sustain. Dev. 34, 667–676 (2013).
Henriksen, T. M. & Breland, T. A. Carbon mineralization, fungal and bacterial growth, and enzyme activities as affected by contact between crop residues and soil. Biol. Fertil. Soils 35(1), 41–48 (2002).
Chet, I., Henis. Y. Trichoderma as a biological control agent against soilborne root pathogens in Ecology and Management of Soilborne Pathogens, Proceedings of section 5 of the 4th International Congress of Plant Pathology (ed. Parker, C.A., Rovina, A.D., Moore, K.I., Wong, P.T.W. & Koumorgen, J.F.) 110–112 (1985).
Chet I. Trichoderma—Application, mode of action and potential as a biocontrol agent of soilborne plant pathogens in Innovative Approaches to Plant Disease Control (ed. Chet, I.) 137–160 (John Wiley and Sons, 1987).
Cumagun, C. J. R., Manalo, J. O., Salcedo-Bacalangco, N. A. & Ilag, L. L. Cellulose decomposing ability of Trichoderma in relation to their saprophytic survival. Arch. Phytopathol. Plant Protect. 42(7), 698–704 (2009).
Islam, M. A., Mostafa, M. G. & Rahman, M. R. Conversion of solid organic waste into compost using Trichoderma spp and its application on some selected vegetables. Int. J. Environ. Waste Manage. 14(3), 211–221 (2014).
Karumuri, L. Effect of Trichoderma isolates on decomposition of paddy straw and incidence of Alternaria Blight (Alternaria triticina Prasada and Prabhu) of wheat under a rice–wheat cropping system. PhD Thesis. Department of Plant Pathology, College of Agriculture Faculty Agriculture Indira Gandhi Krishi Vishwavidyalaya Raipur (Chhattisgarh). (2017).
Sannathimmappa, H. G., Gurumurthy, B. R., Jayadeva, H. M. & Shivanna, M. B. Effective recycling of paddy straw through microbial degradation for enhancing grain and straw yield in rice. IOSR J. Agric. Veterinary Sci. 8(1), 70–73 (2015).
Amira, R. D. et al. Bioconversion of empty fruit bunches (EFB) and palm oil mill effluent (POME) into compost using Trichoderma virens. Afr. J. Biotechnol. 10, 18775–18780 (2011).
Bolton, R. G. & Boddy, L. Characterization of the spatial aspects of foraging mycelial cord systems using fractal geometry. Mycol. Res. 97, 762–768 (1993).
Horwath, W. Carbon cycling: the dynamics and formation of organic matter in Soil Microbiology, Ecology, and Biochemistry 4th edition (ed. Paul E.A.) 339–382 (Academic Press, New York, 2015).
Kausar, H., Sariah, M., Saud, H. M., Alam, M. Z. & Ismail, M. R. Development of compatible lignocellulolytic fungal consortium for rapid composting of rice straw. Int. Biodeterior. Biodegradation 64(7), 594–600 (2010).
Mukhlis, H. et al. Potential of lignocellulolytic Trichoderma for bioconversion of oil palm empty fruit bunches. Aust. J. Crop Sci. 7(3), 425–431 (2013).
USDA NRCS. Carbon to nitrogen ratios in cropping systems. Preprint at https://www.nrcs.usda.gov (2011).
Bird, J. A., Horwath, W. R., Eagle, A. J. & Van Kessel, C. Immobilization of fertilizer nitrogen in rice. Soil Sci. Soc. Am. J. 65, 1143–1152 (2001).
Van Kessel, J. S., Reeves, J. B. & Meisinger, J. J. Nitrogen and carbon mineralization of potential manure components. J. Environ. Qual. 29, 1669–1677 (2000).
Qian, P. & Schoenau, J. Availability of nitrogen in solid manure amendments with different C: N ratios. Can. J. Soil Sci. 82, 219–225 (2002).
Thuy, N. H., Shan, Y., Bijay-Singh, W. K., Cai, Z. & Yadvinder-Singh, B. R. J. Nitrogen supply in rice- based cropping systems as affected by crop residue management. Soil Sci. Soc. Am. J. 72, 514–523 (2008).
Chesire, M. V. et al. The immobilization of nitrogen by straw decomposition in soil. Eur. J. Soil Sci. 50, 329–341 (1999).
Henriksen, T. M. & Breland, T. A. Evaluation of criteria for describing crop residues degradability in a model of carbon and nitrogen turnover in soil. Soil Biol. Biochem. 31, 1135–1149 (1999).
Azam, F., Lodhi, A. & Ashraf, M. Availability of soil and fertilizer N to wetland rice following wheat straw amendment. Biol. Fertil. Soils 11, 97–100 (1991).
Acknowledgements
This research was funded by (1) The Philippine Department of Agriculture through the project “Sustainable rice straw management for bioenergy, food, and feed in the Philippines”, Grant Ref 101101-03-042-2018 and (2) the CGIAR Research Program on Rice (RICE). The authors convey their thankful appreciation to staff of the Zeigler Experiment Station at the International Rice Research Institute (IRRI) for providing the facilities, fields, and support services for this research. We also thank the Mechanization and Postharvest Cluster, Sustainable Impact Platform, IRRI, for its support over the duration of the study. The valuable support of research technicians and other students from the University of the Philippines Los Baños is also acknowledged.
Funding
Philippine Department of Agriculture through the project “Sustainable rice straw management for bioenergy, food, and feed in the Philippines", 101101-03-042-2018, 101101-03-042-2018, 101101-03-042-2018, 101101-03-042-2018, 101101-03-042-2018, 101101-03-042-2018, CGIAR Research Program on Rice (RICE).
Author information
Authors and Affiliations
Contributions
N.D.O. led the research. S.M.J.M.G. and H.G.P collected data and performed the soil analysis. J.M.S. developed the research protocol, implemented the experiment and collected agronomic data. N.V.H. and M.G. provided the general supervision and reviewed the manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
The original online version of this Article was revised: The original version of this Article contained an error in Figure 1 where the x-axis was incorrectly numbered and the labels were illegible.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Organo, N.D., Granada, S.M.J.M., Pineda, H.G.S. et al. Assessing the potential of a Trichoderma-based compost activator to hasten the decomposition of incorporated rice straw. Sci Rep 12, 448 (2022). https://doi.org/10.1038/s41598-021-03828-1
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-021-03828-1
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
