
Abstract
In colonially breeding marine predators, individual movements and colonial segregation are influenced by seascape characteristics. Tidewater glacier fronts are important features of the Arctic seascape and are often described as foraging hotspots. Albeit their documented importance for wildlife, little is known about their structuring effect on Arctic predator movements and space use. In this study, we tested the hypothesis that tidewater glacier fronts can influence marine bird foraging patterns and drive spatial segregation among adjacent colonies. We analysed movements of black-legged kittiwakes (Rissa tridactyla) in a glacial fjord by tracking breeding individuals from five colonies. Although breeding kittiwakes were observed to travel up to ca. 280 km from the colony, individuals were more likely to use glacier fronts located closer to their colony and rarely used glacier fronts located farther away than 18 km. Such variation in the use of glacier fronts created fine-scale spatial segregation among the four closest (ca. 7 km distance on average) kittiwake colonies. Overall, our results support the hypothesis that spatially predictable foraging patches like glacier fronts can have strong structuring effects on predator movements and can modulate the magnitude of intercolonial spatial segregation in central-place foragers.
Similar content being viewed by others


Introduction
Identification of key factors driving animal distribution patterns and movements is a fundamental goal in ecology1,2. In various species, individuals must return to a central base between foraging bouts, greatly shaping their spatial ecology3,4,5. Optimal foraging models assume that central‐place foragers, such as colonial-nesting species, are adapted to maximize their rate of net energy gain per unit of time and that the costs of foraging increase with increasing distance from the colony6,7. If foragers do not exhibit territorial defence and if prey are abundant and uniformly and/or unpredictably distributed, then the theory predicts that the density of foragers should decrease with increasing distance from the colony. Under such conditions, foraging areas of neighbouring colonies could overlap when separated by less than the distance covered by their respective foraging ranges. However, segregated foraging grounds of neighbouring colonies appear widespread among colonial central‐place foragers like eusocial insects8, bats9, pinnipeds10, and seabirds11.
Several factors could promote spatial segregation between neighbouring colonies in seabirds, including intraspecific competition12,13, distribution of predators14, individual specialization in space use11, and associated cultural effects5,13,15. Prey distribution around colonies is another potential driver of their spatial segregation16,17,18. Since optimal foraging models predict that individuals should mainly exploit patches closer to their colony to maximize their net energy gain6,7, prey distribution and physical structures of the seascape that lead to their aggregations could have an important effect on how animals distribute themselves within the shared foraging ranges11,17,18. The patchiness of prey (i.e., patch size and distribution) and its associated predictability (i.e., spatial and temporal) appear as important drivers of the spatial structuring of seabird movements19,20,21. These factors could thus modulate the level of segregation between neighbouring colonies11,16,17.
Tidewater glaciers fronts are known to be important foraging hotspots for seabirds and other marine predators22,23,24. They have recently gained increasing attention due to their potential to alleviate the short-term negative effect of ice-associated habitat loss on Arctic wildlife25,26. Driven by the meltwater glacier discharge at the fronts, zooplankton are entrained and transported by the buoyant subglacial plumes at the ice-sea interface, making prey readily available at the surface for predators22,27,28,29. Although their relative profitability can vary over time30,31,32,33,34, glacier fronts may nonetheless represent spatially predictable foraging habitats for breeding colonial seabirds30,31,33 due to their geographically restricted and relatively fixed location. Despite their documented importance as foraging hotspots, little is known about their structuring effect on predator distribution patterns and movements.
Focusing on Arctic breeding black-legged kittiwakes (Rissa tridactyla), our study aims at testing the hypothesis that tidewater glacier fronts represent high value patches that modulate seabird foraging patterns and generate fine-scale spatial segregation among neighbouring colonies. Kittiwakes are surface-feeders, central place foragers during the breeding season, and are commonly observed at tidewater glacier fronts22,30,31. We predicted that kittiwakes breeding within a glacial fjord (Kongsfjorden, Svalbard) in the Arctic should predominantly use glacier fronts located closer to their colony. Then, while controlling for differences in colony size and distance separating colonies11,13, we further predicted that differential use of glacier fronts promotes a fine-scale spatial segregation (i.e., less overlap than what would be expected from distance alone) among neighbouring colonies.
Methods
Study system
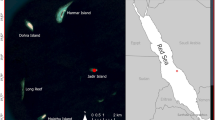
This study was conducted in the Kongsfjorden region, Svalbard, during the chick-rearing period of 2017. The study area was delimited by a radius of 50 km from the centre of Kongsfjorden (Fig. 1). In total, 25 tidewater glacier fronts are present in the area and are used to a varying extent by kittiwakes breeding in the five studied colonies, i.e., Blomstrand (12.11° E, 78.99° N), Krykkjefjellet (12.18° E, 78.89° N), Observasjonsholmen (12.28° E, 78.93° N), Ossian Sarsjellet (12.44° E, 78.92° N), and one colony outside the fjord; Fuglehuken (10.47° E, 78.89° N) (Fig. 1). Following standardized procedures35, breeding pairs at each colony were counted once between 2011 and 2017 to assess colony size (Table 1). The kittiwake population of Svalbard was relatively stable from 2009 to 201936, and hence we assumed that colony sizes assessed a few years prior to 2017 were representative of the colony sizes during our study.

Svalbard archipelago (top; land in black), (a) the 50 km radius area corresponding to the regional scale, and (b) the Kongsfjorden area corresponding to the fjord scale. Kittiwake breeding colonies (circles: BLM (purple) = Blomstrand; FGL (dark blue) = Fuglehuken; KRY (cyan) = Krykkjefjellet; OBS (green) = Observasjonsholmen; OSS (yellow) = Ossian Sarsfjellet) and glacier fronts (blue areas) are also shown. Maps were generated using R (version 4.0.242; URL: https://www.R-project.org/).
GPS tracking
In total, 48 birds were caught at the nest using a noose pole and were fitted with a GPS device (minimum and maximum dates of deployment: 7–28 July). Each individual was sexed, either by a molecular approach using DNA from blood or feather samples (69% of the individuals; see details in Harris et al.37) or by morphometry, using a cut-off value (\(x\)) of 90.5 mm on the birds’ head-bill length38 (31% of individuals: i.e., female ≤ \(x\) > male). All birds had at least one chick hatched at the time of logger deployment. Three different types of GPS were deployed in random order (i.e., i-gotU GT-120, Mobile Action; CatLog Gen1, and CatLog Gen2). All GPSs were sealed in waterproof heat-shrink tubes, and their total mass ranged from 7.3–18.1 g (\(\tilde{x } \pm SD\); 13.9 ± 2.9), representing 2–5% of the birds’ mass. Loggers were fitted to the birds’ back feathers with TESA tape. We found no indication that the mass of the GPS modified bird foraging behaviour (see details in Supplementary Information S1).
On average, the GPS devices deployed on birds were retrieved after 71.2 h (range = 23.5–160.5 h). Sampling intervals ranged from 2 to 10 min, and all tracks were subsampled to obtain a standard interval of 10 min between consecutive locations. Speed was filtered for a maximum of 80 km h−1 to remove aberrant locations39,40. We defined a trip as (1) all consecutive GPS positions located outside the colony area, which was defined as an area of 200-m radius from its centroid, and (2) being outside the colony’s area for a minimum of 50 min (thresholds based on sensitivity analyses, see Bertrand et al.34). To limit the number of non-foraging trips in the analyses, we excluded from subsequent analyses all tracks that had a proportion of locations over land equal to or greater than 50%, as kittiwakes can spend time bathing in freshwater41 and fetching nest materials on land. Moreover, only complete tracks were considered in the analyses (i.e. trips for which locations were registered until the bird returned to the colony). This choice did not bias samples towards shorter trips (i.e., we found no difference between the “complete” and “non-complete” groups in their maximum distances travelled per trip; Mann–Whitney U-test; U = 22 645, p = 0.94). In total, 205 foraging trips were used in this study (Table 1, see details in Supplementary Information S2), ranging from 1–10 per individual (\(\overline{x } \pm SD\); 4.3 ± 2.7).
All bird captures and handling were performed following relevant guidelines and regulations regarding animal welfare in Norway (Forsøksdyrforvatningen tilsyns- og søknadssystem, FOTS). All field protocols were formerly approved by the ethics committees of the Governor of Svalbard (program for research 361) and the Norwegian Food Safety Authority (Mattilsynet; permits #6439, #8602, and #15503).
Data analysis
All analyses were performed with R (version 4.0.242).
Estimation of individual-level foraging ranges
We used an autocorrelated kernel density approach (AKDE43,44) to estimate each individual’s probability distribution (hereafter utilization distribution; UD) using the ctmm45 and ctmmweb46 packages. Contrary to conventional range estimates (e.g., Kernel Density Estimation (KDE), Minimum Convex Polygon), AKDE does not assume independency and identical distribution (IID) among GPS locations, and explicitly accounts for the temporal autocorrelation inherent to every trip while calculating an individual’s space use47,48. To reduce the weight of the locations at the colonies in individual UD calculations, we considered all consecutive trips from the same individual as continuous while removing all points located at the colony. All individual UDs were computed over a grid size of 500 m. In total, three different models were tested for each individual: (1) the IID, as assumed by conventional KDE, (2) the Ornstein–Uhlenbeck (OU: featuring positional autocorrelation timescale); and (3) the OU-Foraging processes (OUF: featuring both velocity and positional autocorrelation timescale)49. Model selection for each individual’s UD estimation was performed using the Akaike Information Criterion with correction for small sample sizes (AICc), and the model with the lowest AICc was selected50,51. We thereafter used the effective sample size (\({\widehat{N}}_{area}\)) as a proxy of an individual’s UD representativeness, which theoretically corresponds to the number of statistically independent positions represented by the tracks, corresponding roughly to the number of range crossings undergone by the animal during the tracking period43,48. A minimum \({\widehat{N}}_{area} \ge 4.5\) for each individual’s UD was considered as a minimal threshold47,48 (see details in Supplementary Information S3). Based on this threshold, 10 of the 13 individuals (i.e. ~ 77%) from Fuglehuken were filtered out. Finally, we assessed the representativeness of colony-level grouping as a function of sample sizes using the bootstrap approach described in Lascelles et al.52 and implemented via the track2kba package53 (see details in Supplementary Information S3).
Quantifying segregation level among colonies
Spatial segregation among colonies was evaluated using the calculated individual UDs and by extracting the Bhattacharyya coefficient (BA) calculated upon pairs of individual UDs54,55,56,57. This index relies on the probability density functions of the two UDs and does not require any subjective choice of specific quantile contours54. We used the inverse coefficient (i.e., \(1-BA\); hereafter called the segregation index, SI), which ranged from 0 (complete overlap) to 1 (full segregation) and was computed using the ctmm package45 along the approximate debiasing correction for small sample size54. We used an analysis of similarities (ANOSIM) for testing the null hypothesis that the dissimilarity between colonies is equal to or lower than the dissimilarity among individuals within colonies58, or in other words, that segregation in space use between members of different colonies is equal to or weaker than that of members belonging to the same colony. For that purpose, we used the segregation matrix (i.e., pairwise segregation index (SI) among all individuals) as a response variable and the colony affiliation as a grouping factor. We considered that the hypothesis of segregation among colonies was supported if the dissimilarity among colonies was greater than the dissimilarity within colonies, using a nominal threshold of 0.05 along 999 permutations59,60. If the test was significant at the population-level (i.e., involving all colonies), we then tested segregation on a pairwise basis, this time applying ANOSIM analyses to each colony dyad. Since multiple testing was conducted, we controlled the false discovery rate using the Benjamini–Hochberg procedure61. Since the ANOSIM approach is sensitive to intragroup dispersion58, we also tested for the null hypothesis of homogeneity in group multivariate dispersion62. ANOSIM and the test for multivariate homogeneity were performed using the vegan package63.
Effect of distance on the probability to use glacier fronts
We used a generalized linear mixed model with a binomial distribution to test the prediction that kittiwakes were more likely to use glacier fronts located closer to their colony rather than those located farther away. We tested the prediction at two spatial scales (i.e., regional scale and fjord scale). First, to include all glaciers that birds could potentially use, we considered a radius of 50 km from the Kongsfjorden centre (corresponding to the distance of the most distant glacier front used by kittiwakes in our dataset). A total of 25 glacier fronts were located within that radius (i.e., regional scale; Fig. 1a). Second, we considered only the four colonies and 6 glacier fronts located within Kongsfjorden to confirm that the relationship observed at the regional scale was similar at a finer scale (i.e., fjord scale; Fig. 1b). We defined our response variable as binary: a value of 1 was assigned to a specific front if the birds used it during a given foraging trip, and the value 0 was assigned if they did not. We defined the front areas by creating a spatially-explicit buffer around the front line (radius = 400 m). Based on a previous study34, a glacier front was considered to be used when two consecutive locations (10-min interval) were within the buffer zone.
Distances between a colony and glacier’s front line centroid were calculated using the Great Circle distance via the sp package64,65 and were subsequently used as predictors of glacier front use. We also included individual sex as a predictor to control for potential variation between males and females37,39,66. The interaction between distance and sex was investigated to ensure that the space use patterns of colonies was not driven by a particular sex. Moreover, the colony was included as a fixed effect to account for the average difference in glacier front use among colonies. Significance of each predictor was evaluated by likelihood ratio tests for nested model, comparing the fits with (H1) and without (H0) the addition of the corresponding variable (nominal threshold of 0.05). For all models, we fitted the individual ID as random factors. We found no sign of overdispersion in the full models using the nonparametric dispersion test over quantile residuals (p = 0.87) available via the dharma package67.
Drivers of intercolony spatial segregation
Drivers of spatial segregation among colonies were investigated by means of a multiple regression on distance matrices (MRM68) using the ecodist package69. Colony size (Table 1), the distance separating colonies, and the dissimilarity in glacier front use were used as predictors and the segregation matrix as the response variable. We calculated the dissimilarity in glacier front use among individuals by using individual-level counts of use of each glacier (i.e., from the binary variable detailed in the previous section) and the pairwise individual similarity index from Bolnick et al.70:
where pik and pjk represent the proportion of use of glacier k for the two selected birds (i, j). Since birds performed several trips during their tracking period, we first summed the counts by glacier during the trips of each individual before calculating the proportions. One individual (out of 13 birds) from Observasjonsholmen and all birds from Fuglehuken did not use any fronts during their tracking period and were thus removed from the analysis. For ease of interpretation, we subsequently used the inverse index, i.e., \(DH=1-H\), hence generating a dissimilarity index. The dissimilarity index (DH) in glacier front use thus ranged from 0 (complete overlap in glacier front use) to 1 (complete segregation in glacier front use) and was calculated via the package rinsp71.
Due to its potential importance in modulating intraspecific competition for prey acquisition11,13,72, we controlled for colony size while investigating drivers of intercolonial segregation. We used the Manhattan distance, also known as the \({L}^{1}\) norm, to calculate the difference among the colony sizes. The Euclidean distance was chosen for calculating the distance separating colony centroid of each individual. Predictor significance was evaluated via permutations along 999 iterations and using the nominal threshold of 0.05.
Results
Space use varied substantially among the different kittiwake colonies. Birds breeding in the Fuglehuken colony showed no overlap with any glacier front, although the closest front was only 23 km away from the colony. Although all colonies have access to the pelagic zone, only birds from Fuglehuken relied exclusively on the pelagic environment to forage (Fig. 2a, see Supplementary Information S2 for raw tracks). Within Kongsfjorden, the core-use area derived for each colony (i.e., 50% isopleth) showed highly restricted and delimited space use, which always overlapped at least one glacier front (Fig. 2b). Representativeness analyses suggested that most of the intracolonial variation in space use was captured by colony UD estimates (range: 84–95%, see Supplementary Information S3 for details).

(a) Study area and the mean utilization distribution (UD) estimated for each kittiwake breeding colony (95% and 50% isopleths based on the individuals’ UD weighted by the number of trips sampled per individual; BLM (purple) = Blomstrand, FGL (dark blue) = Fuglehuken, KRY (cyan) = Krykkjefjellet, OBS (green) = Observasjonsholmen, OSS (yellow) = Ossian Sarsfjellet) and (b) core-ranges (50% isopleths) for the four colonies (circles) located within Kongsfjorden. Glacier fronts (crosshatched) are also shown. All birds were tracked during the chick-rearing period in 2017. Maps were generated using R (version 4.0.242; URL: https://www.R-project.org/).
The global analysis of similarities (ANOSIM) indicated that the dissimilarity in space use was greater among colonies (average: 0.57; range: 0.39–0.98) than within colonies (average: 0.27; range: 0.17–0.50). This indicates that the overlap between foraging ranges was higher among birds from the same colony than among birds from different colonies (ANOSIM-R = 0.59, p = 0.001). The same pattern was observed when considering only the four colonies located within Kongsfjorden (ANOSIM-R = 0.50, p = 0.001). While showing no sign of heterogeneous dispersion among groups (F4,33 = 0.93, p = 0.457), the ANOSIM analysis conducted on each pair of colonies suggested that all colonies showed significant spatial segregation, except the Blomstrand (BLM)—Observasjonsholmen (OBS) dyad (Table 2). Although the core-use area of these two colonies showed little overlap, it suggests that some birds breeding in these colonies shared similar foraging space (see raw tracks distribution in Supplementary Information S2).
Glacier front use varied substantially for birds breeding in various neighbouring colonies located within Kongsfjorden (see details in Supplementary Information S4). The use of glacier fronts decreased significantly with the distance separating them to a given colony at the two different spatial scales used for the analysis (Table 3, Fig. 3). A threshold around 18 km was also observed at the regional scale, which corresponded to the maximum distance separating a colony and a front located within Kongsfjorden (Fig. 3a). This indicates that kittiwakes breeding in Kongsfjorden essentially used glacier fronts located inside the fjord. A similar decaying relationship was observed at the fjord scale when considering only colonies and glacier fronts occurring within Kongsfjorden (Fig. 3b). All models showed no effect of individual sex (Table 3).

Relationship between black-legged kittiwake use of glacier fronts as a function of the distance separating their colonies to a given front at two different scales; (a) regional scale; involving 5 colonies and 25 glacier fronts (effect of distance; estimate = − 0.22, 95% CI = − 0.26: − 0.18) and (b) fjord scale; involving 4 colonies and 6 glacier fronts (effect of distance; estimate = − 0.16, 95% CI = − 0.22: − 0.09). For the ease of representation, points represent individual males and females average use (circles: yellow = female; green = male) of glacier fronts. Solid curves and shaded areas are the regression line and associated 95% confidence interval (back-transformed) estimated from generalized linear mixed models, using the glacier front use as response variable, the distance, sex and colony as additive fixed predictors and the bird ID as random factor.
Spatial segregation in foraging ranges of breeding kittiwakes was positively correlated to the distance separating their colonies (estimate = 2.17 × 10−5, p = 0.001) and the level of dissimilarity in glacier front use (DH; estimate = 0.41, p = 0.001; Fig. 4). Surprisingly, however, segregation was not related to the relative size of the colonies (estimate = 8.31 × 10–6, p = 0.44; Fig. 4). Overall, the distance separating colonies and the dissimilarity in glacier front use explained about 41% of the spatial segregation in foraging ranges (F = 191.10, p = 0.001).

Spatial segregation level (i.e., 1 − Bhattacharyya coefficient) of breeding kittiwake foraging ranges as a function of (a) the difference in colony size (i.e., Manhattan distance), (b) the distance separating colonies, and (c) the dissimilarity in glacier front use between each individual dyad. Solid and dashed lines depict, respectively, the significant and non-significant partial slopes of the multiple regression on distance matrices. Intercepts of each slope have been adjusted for illustration using as reference the mean of the remaining covariates. Symbols represent the median and associated 25th and 75th percentiles in panels (a) and (b).
Discussion
The investigation of marine predator movements is central to understanding how individuals respond to seascape dynamics and structures11,73,74. In this study, we tested the hypothesis that spatially-predictable seascape elements like tidewater glacier fronts can influence kittiwake foraging patterns and drive spatial segregation among adjacent colonies. Although chick-rearing kittiwakes can feed up to ca. 280 km away from their colony (see also Christensen-Dalsgaard et al.40), individuals in Kongsfjorden were more likely to use glacier fronts located closer to their colony and rarely used glacier fronts located > 18 km away. As predicted, the dissimilarity in the use of glacier fronts partly explained the spatial segregation observed between adjacent kittiwake colonies. Overall, our results illustrate how spatially-predictable foraging patches like glacier fronts can modulate the magnitude of intercolonial segregation in central-place foragers.
Optimal foraging theory predicts that central-place foragers should spend more time in highly profitable foraging patches located close to their breeding colony75,76. When two neighbouring colonies are separated by less than the distance covered by their members’ foraging range, patch predictability and distribution between the colonies could strongly affect the behaviour of foraging individuals and determine their level of spatial segregation11,17. As predicted, the spatial distribution of tidewater glacier fronts, which can offer relatively predictable foraging patches to various marine predators22,30,33, seems to partly drive consumer movements and promotes small-scale intercolonial segregation in our study system. However, glacier fronts might not offer equal foraging opportunity to breeding birds33, and the relatively low explanatory power of the distance between colony and glacier fronts on the probability of front use highlights the needs for further research on the relative profitability of these foraging patches.
Tidewater glacier fronts are often defined as foraging hotspots for various Arctic predators22,26,31. Subglacial plumes constitute major components of the mechanism modulating glacier front profitability for marine predators28,30. Originating from the subglacial meltwater discharge, the buoyant meltwater plume entrains water masses from intermediate depths and transports prey to the surface while making them accessible to predators22,28,30. A plume’s entrainment capacity relies on both the discharge velocity and the depth at which the subglacial plume originates29,77, which are both known to vary among glacier fronts in Kongsfjorden33,78,79. Glacier fronts in Kongsfjorden thus likely offer variable entrainment capacity and level of predictability. Moreover, the relative profitability of a given glacier front can vary depending on the environmental conditions30,31,32,33,34. Integrating such spatiotemporal variation in the profitability of glacier fronts should strongly help understanding their impacts on the spatial distribution of Arctic marine predators78,80. Along deglaciation, however, retreat of glacier termini above the sea-level in Kongsfjorden could ultimately reduce foraging opportunities in the fjord and modify the spatial distribution of seabirds by increasing their use of more distant, pelagic feeding areas as seen in kittiwakes breeding outside the fjord (see Fig. 2).
The spatial distribution of marine predators is known to be dependent on the distribution and patchiness of prey, which is strongly affected by physical forcings that promote their aggregations73,81. Eddies66,82,83 and fronts84,85,86,87 are oceanographic features that are associated with important foraging habitats for seabirds, by either enhancing primary production through the transport of nutrients or by the advection of biomass88. Ultimately, the spatiotemporal variability of these physical processes can modulate the level of predictability of prey aggregations in the seascape, which in turn affects the profitability of these patches to marine predators19,20,74. In parallel, foraging patches like glacier fronts that are predictable in space, and to some extent in time during a given summer89, are likely to promote foraging site fidelity20,90 and aggregations in seabirds91,92. If knowledge of locations of prey and/or foraging grounds is transferable to conspecifics at the colony level through the use of public information (e.g., bearings of departing/returning foraging birds93,94), then a cultural foraging pattern could arise and enhance intercolonial difference in space use5,11,15.
Our results add to a growing body of research indicating that segregation level in marine bird colonies is affected by seascape structures11,17,18,95. In a study conducted on Cory’s shearwater (Calonectris borealis), Ramos et al.17 showed that birds from different colonies tended to segregate in most of their foraging ranges, but overlapped largely in areas characterized by one predictable but distant foraging patch (i.e., a large upwelling area17). Similarly, Dean et al.18 showed that colonies of Manx shearwater (Puffinus puffinus) were more likely to segregate around their colony while overlapping at a distant, productive tidal front system. Taken altogether, these results indicate that the distribution of high-profitability foraging hotspots can have important effects on the spatial segregation of colonial breeding predators11,17. The varying level of segregation over such foraging patches (from overlap to segregation) likely reflects the distribution of individuals maximizing their net energy gain among all patches available in the seascape75,96. Further research is warranted to understand the complex relationship between central-place forager behaviors, space use and seascape heterogeneity20,74,88.
Data availability
The dataset that supports current findings is available from the dryad digital repository: https://doi.org/10.5061/dryad.bnzs7h4c1. R codes can be accessed from the github repository: https://github.com/PhilBertrand/GF_segregation.
References
Nathan, R. et al. A movement ecology paradigm for unifying organismal movement research. Proc. Natl. Acad. Sci. 105, 19052–19059 (2008).
Sutherland, W. J. et al. Identification of 100 fundamental ecological questions. J. Ecol. 101, 58–67 (2013).
Bernstein, R. A. & Gobbel, M. Partitioning of space in communities of ants. J. Anim. Ecol. 48, 931–942 (1979).
Jenkins, S. H. A size-distance relation in food selection by beavers. Ecology 61, 740–746 (2018).
Grémillet, D. et al. Offshore diplomacy, or how seabirds mitigate intra-specific competition: A case study based on GPS tracking of Cape gannets from neighbouring colonies. Mar. Ecol. Prog. Ser. 268, 265–279 (2004).
Orians, G. H. & Pearson, N. E. On the theory of central place foraging. In Analysis of Ecological Systems (eds Horn, D. J. et al.) 154–177 (The Ohio State University Press, 1979).
Schoener, T. W. Generality of the size-distance relation in models of optimal feeding. Am. Nat. 114, 902–914 (1979).
Brown, M. J. F. & Gordon, D. M. How resources and encounters affect the distribution of foraging activity in a seed-harvesting ant. Behav. Ecol. Sociobiol. 47, 195–203 (2000).
Dawo, B., Kalko, E. K. V. & Dietz, M. Spatial organization reflects the social organization in Bechstein’s bats. Ann. Zool. Fennici 50, 356–370 (2013).
Nordstrom, C. A., Battaile, B. C., Cotté, C. & Trites, A. W. Foraging habitats of lactating northern fur seals are structured by thermocline depths and submesoscale fronts in the eastern Bering Sea. Deep Res. Part II Top. Stud. Oceanogr. 88–89, 78–96 (2013).
Bolton, M., Conolly, G., Carroll, M., Wakefield, E. D. & Caldow, R. A review of the occurrence of inter-colony segregation of seabird foraging areas and the implications for marine environmental impact assessment. Ibis (London 1859) 161, 241–259 (2019).
Cairns, D. K. The regulation of seabird colony size: A hinterland model. Am. Nat. 134, 141–146 (1989).
Wakefield, E. D. et al. Space partitioning without territoriality in gannets. Science (80-.) 341, 68–70 (2013).
Masello, J. F. et al. Diving seabirds share foraging space and time within and among species. Ecosphere 1, art19 (2010).
Cecere, J. G. et al. Spatial segregation of home ranges between neighbouring colonies in a diurnal raptor. Sci. Rep. 8, 11762 (2018).
Ainley, D. G., Ford, R. G., Brown, E. D., Suryan, R. M. & Irons, D. B. Prey resources, competition, and geographic structure of Kittiwake colonies in Prince William Sound. Ecology 84, 709–723 (2003).
Ramos, R. et al. Meta-population feeding grounds of cory’s shearwater in the subtropical Atlantic ocean: Implications for the definition of marine protected areas based on tracking studies. Divers. Distrib. 19, 1284–1298 (2013).
Dean, B. et al. Simultaneous multi-colony tracking of a pelagic seabird reveals cross-colony utilization of a shared foraging area. Mar. Ecol. Prog. Ser. 538, 239–248 (2015).
Hunt Jr., G. L. et al. Physical processes, prey abundance, and the foraging ecology of seabirds. In Proceedings of the 22nd International Ornithological Congress, Durban (eds Adams, N. J. & Slotow, R. H.) 2040–2056 (BirdLife South Africa, 1999).
Weimerskirch, H. Are seabirds foraging for unpredictable resources? Deep Res. Part II Top. Stud. Oceanogr. 54, 211–223 (2007).
Benoit-Bird, K. J. et al. Prey patch patterns predict habitat use by top marine predators with diverse foraging strategies. PLoS ONE 8, e53348 (2013).
Lydersen, C. et al. The importance of tidewater glaciers for marine mammals and seabirds in Svalbard, Norway. J. Mar. Syst. 129, 452–471 (2014).
Hamilton, C. D., Lydersen, C., Ims, R. A. & Kovacs, K. M. Coastal habitat use by ringed seals Pusa hispida following a regional sea-ice collapse: Importance of glacial refugia in a changing Arctic. Mar. Ecol. Prog. Ser. 545, 261–277 (2016).
Nishizawa, B. et al. Contrasting assemblages of seabirds in the subglacial meltwater plume and oceanic water of Bowdoin Fjord, northwestern Greenland. ICES J. Mar. Sci. 77, 711–720 (2020).
Grémillet, D. et al. Arctic warming: Nonlinear impacts of sea-ice and glacier melt on seabird foraging. Glob. Change Biol. 21, 1116–1123 (2015).
Hamilton, C. D. et al. Contrasting changes in space use induced by climate change in two Arctic marine mammal species. Biol. Lett. 15, 20180834 (2019).
Hartley, C. H. & Fisher, J. The marine foods of birds in an inland fjord region in West Spitsbergen: Part 2. Birds. J. Anim. Ecol. 5, 370–389 (1936).
Everett, A. et al. Subglacial discharge plume behaviour revealed by CTD-instrumented ringed seals. Sci. Rep. 8, 13467 (2018).
Carroll, D. et al. Modeling turbulent subglacial meltwater plumes: Implications for fjord-scale buoyancy-driven circulation. J. Phys. Oceanogr. 45, 2169–2185 (2015).
Urbański, J. A. et al. Subglacial discharges create fluctuating foraging hotspots for seabirds in tidewater glacier bays. Sci. Rep. 7, 43999 (2017).
Stempniewicz, L. et al. Marine birds and mammals foraging in the rapidly deglaciating Arctic fjord—Numbers, distribution and habitat preferences. Clim. Change 140, 533–548 (2017).
Stempniewicz, L. et al. Advection of Atlantic water masses influences seabird community foraging in a high-Arctic fjord. Prog. Oceanogr. 193, 102549 (2021).
Dragańska-Deja, K., Błaszczyk, M., Deja, K., Wesławski, J. M. & Rodak, J. Tidewater glaciers as feeding spots for the Black-legged kittiwake (Rissa tridactyla): A citizen science approach. Polish Polar Res. 41, 69–93 (2020).
Bertrand, P. et al. Feeding at the front line: interannual variation in the use of glacier fronts by foraging black-legged kittiwakes. Mar. Ecol. Prog. Ser. 677, 197–208 (2021).
Walsh, P. M. et al. Seabird Monitoring Handbook for Britain and Ireland. A Compilation of Methods for Survey and Monitoring of Breeding Seabirds (JNCC/RSPB/ITE/Seabird Group, 1995).
Anker-Nilssen, T. et al. Key-Site Monitoring in Norway 2018, Including Svalbard and Jan Mayen (SEAPOP, 2020).
Harris, S. M. et al. Personality predicts foraging site fidelity and trip repeatability in a marine predator. J. Anim. Ecol. 89, 68–79 (2020).
Coulson, J. C. Sexing black-legged kittiwakes by measurement. Ringing Migr. 24, 233–239 (2009).
Paredes, R. et al. Proximity to multiple foraging habitats enhances seabirds’ resilience to local food shortages. Mar. Ecol. Prog. Ser. 471, 253–269 (2012).
Christensen-Dalsgaard, S., May, R. & Lorentsen, S. H. Taking a trip to the shelf: Behavioral decisions are mediated by the proximity to foraging habitats in the black-legged kittiwake. Ecol. Evol. 8, 866–878 (2018).
Coulson, J. C. & Macdonald, A. Recent changes in the habits of the Kittiwake. Br. Birds 55, 171–177 (1962).
R Core Team. R: A Language and Environment for Statistical Computing (R Foundation for Statistical Computing, 2020). https://www.R-project.org/.
Fleming, C. H. & Calabrese, J. M. A new kernel density estimator for accurate home-range and species-range area estimation. Methods Ecol. Evol. 8, 571–579 (2017).
Fleming, C. H. et al. Rigorous home range estimation with movement data: A new autocorrelated kernel density estimator. Ecology 96, 1182–1188 (2015).
Fleming, C. H. & Calabrese, J. M. ctmm: Continuous-time movement modeling. R package version 0.5.10. https://cran.r-project.org/package=ctmm (2020).
Dong, X., Fleming, C. H., Noonan, M. J. & Calabrese, J. M. ctmmweb: A Shiny web app for the ctmm movement analysis package. https://github.com/ctmm-initiative/ctmmweb. (2018).
Fleming, C. H., Noonan, M. J., Medici, E. P. & Calabrese, J. M. Overcoming the challenge of small effective sample sizes in home-range estimation. Methods Ecol. Evol. 10, 1679–1689 (2019).
Noonan, M. J. et al. A comprehensive analysis of autocorrelation and bias in home range estimation. Ecol. Monogr. 89, 1–21 (2019).
Fleming, C. H. et al. From fine-scale foraging to home ranges: A semivariance approach to identifying movement modes across spatiotemporal scales. Am. Nat. 183, E154–E167 (2014).
Calabrese, J. M., Fleming, C. H. & Gurarie, E. Ctmm: An R package for analyzing animal relocation data as a continuous-time stochastic process. Methods Ecol. Evol. 7, 1124–1132 (2016).
Burnham, K. P. & Anderson, D. R. Model selection and multimodel inference: A practical information-theoretic approach (Springer, 2002).
Lascelles, B. G. et al. Applying global criteria to tracking data to define important areas for marine conservation. Divers. Distrib. 22, 422–431 (2016).
Beal, M. et al. BirdLifeInternational/track2kba: First Release (Version 0.5.0). Zenodo. https://doi.org/10.5281/zenodo.3823902 (2020).
Winner, K. et al. Statistical inference for home range overlap. Methods Ecol. Evol. 9, 1679–1691 (2018).
Fieberg, J. & Kochanny, C. O. Quantifying home-range overlap: The importance of the utilization distribution. J. Wildl. Manag. 69, 1346–1359 (2005).
Bhattacharyya, A. On a measure of divergence between two multinomial populations. Indian J. Stat. 7, 401–406 (1946).
Bhattacharyya, A. On a measure of divergence between two statistical populations defined by their probability distribution. Bull. Calcutta Math. Soc. 35, 99–109 (1943).
Anderson, M. J. & Walsh, D. C. I. PERMANOVA, ANOSIM, and the Mantel test in the face of heterogeneous dispersions: What null hypothesis are you testing? Ecol. Monogr. 83, 557–574 (2013).
Carneiro, A. P. B. et al. Consistency in migration strategies and habitat preferences of brown skuas over two winters, a decade apart. Mar. Ecol. Prog. Ser. 553, 267–281 (2016).
Dehnhard, N. et al. High inter-and intraspecific niche overlap among three sympatrically breeding, closely related seabird species: Generalist foraging as an adaptation to a highly variable environment? J. Anim. Ecol. 89, 104–119 (2020).
Benjamini, Y. & Hochberg, Y. Controlling the false discovery rate: A practical and powerful approach to multiple testing. J. R. Stat. Soc. Ser. B 57, 289–300 (1995).
Anderson, M. J. Distance-based tests for homogeneity of multivariate dispersions. Biometrics 62, 245–253 (2006).
Oksanen, J. et al. vegan: Community Ecology Package (2019). R package version 2.5-7. https://CRAN.R-project.org/package=vegan
Bivand, R. S., Pebesma, E. & Gomez-Rubio, V. Applied Spatial Data Analysis with R 2nd edn. (Springer, 2013).
Pebesma, E. & Bivand, R. S. Classes and methods for spatial data in R. R News 5 (2). https://cran.r-project.org/doc/Rnews/ (2005).
Paredes, R. et al. Foraging responses of black-legged kittiwakes to prolonged food-shortages around colonies on the Bering Sea shelf. PLoS ONE 9, e92520 (2014).
Hartig, F. Residual diagnostics for hierarchical (multi-level/mixed) regression models. R package version 0.3.3.0. https://CRAN.R-project.org/package=DHARMa (2020).
Legendre, P., Lapointe, F.-J. & Casgrain, P. Modeling brain evolution from behavior: A permutational regression approach. Evolution 48, 1487–1499 (1994).
Goslee, S. C. & Urban, D. L. The ecodist package for dissimilarity-based analysis of ecological data. J. Stat. Softw. 22, 1–19 (2007).
Bolnick, D. I., Yang, L. H., Fordyce, J. A., Davis, J. M. & Svanbäck, R. Measuring individual-level resource specialization. Ecology 83, 2936–2941 (2002).
Zaccarelli, N., Mancinelli, G. & Bolnick, D. I. RInSp: An R package for the analysis of individual specialisation in resource use. Methods Ecol. Evol. 4, 1018–1023 (2013).
Lewis, S., Sherratt, T. N., Hamer, K. C. & Wanless, S. Evidence of intra-specific competition for food in a pelagic seabird. Nature 412, 816–819 (2001).
Bertrand, A. et al. Broad impacts of fine-scale dynamics on seascape structure from zooplankton to seabirds. Nat. Commun. 5, 5239 (2014).
Fauchald, P. Spatial interaction between seabirds and prey: Review and synthesis. Mar. Ecol. Prog. Ser. 391, 139–151 (2009).
MacArthur, R. H. & Pianka, E. R. On optimal use of a patchy environment. Am. Nat. 100, 603–609 (1966).
Schoener, T. W. Theory of feeding strategies. Annu. Rev. Ecol. Syst. 2, 369–404 (1971).
Carroll, D. et al. The impact of glacier geometry on meltwater plume structure and submarine melt in Greenland fjords. Geophys. Res. Lett. 43, 9739–9748 (2016).
Halbach, L. et al. Tidewater glaciers and bedrock characteristics control the phytoplankton growth environment in a fjord in the Arctic. Front. Mar. Sci. 6, 254 (2019).
Pramanik, A. et al. Hydrology and runoff routing of glacierized drainage basins in the Kongsfjord area, northwest Svalbard. Cryosph. Discuss. [preprint]. https://doi.org/10.5194/tc-2020-197.
Hopwood, M. J. et al. Non-linear response of summertime marine productivity to increased meltwater discharge around Greenland. Nat. Commun. 9, 3256 (2018).
Arimitsu, M. L., Piatt, J. F. & Mueter, F. Influence of glacier runoff on ecosystem structure in Gulf of Alaska fjords. Mar. Ecol. Prog. Ser. 560, 19–40 (2016).
Haney, J. Seabird segregation at Gulf Stream frontal eddies. Mar. Ecol. Prog. Ser. 28, 279–285 (1986).
Hyrenbach, K. D., Veit, R. R., Weimerskirch, H. & Hunt, G. L. Jr. Seabird associations with mesoscale eddies: The subtropical Indian Ocean. Mar. Ecol. Prog. Ser. 324, 271–279 (2006).
Cox, S. L. et al. Seabird diving behaviour reveals the functional significance of shelf-sea fronts as foraging hotspots. R. Soc. Open Sci. 3, 160317 (2016).
Schneider, D. C. Seabirds and fronts: A brief overview. Polar Res. 8, 17–21 (1990).
Bost, C. A. et al. The importance of oceanographic fronts to marine birds and mammals of the southern oceans. J. Mar. Syst. 78, 363–376 (2009).
Durazo, R., Harrison, N. M. & Hill, A. E. Seabird observations at a tidal mixing front in the Irish Sea. Estuar. Coast. Shelf Sci. 47, 153–164 (1998).
Wakefield, E. D., Phillips, R. A. & Matthiopoulos, J. Quantifying habitat use and preferences of pelagic seabirds using individual movement data: A review. Mar. Ecol. Prog. Ser. 391, 165–182 (2009).
How, P. et al. Rapidly changing subglacial hydrological pathways at a tidewater glacier revealed through simultaneous observations of water pressure, supraglacial lakes, meltwater plumes and surface velocities. Cryosphere 11, 2691–2710 (2017).
Navarro, J. & González-Solís, J. Environmental determinants of foraging strategies in Cory’s shearwaters Calonectris diomedea. Mar. Ecol. Prog. Ser. 378, 259–267 (2009).
Irons, D. B. Foraging area fidelity of individual seabirds in relation to tidal cycles and flock feeding. Ecology 79, 647–655 (1998).
Piatt, J. F. et al. Predictable hotspots and foraging habitat of the endangered short-tailed albatross (Phoebastria albatrus) in the North Pacific: Implications for conservation. Deep Res. Part II Top. Stud. Oceanogr. 53, 387–398 (2006).
Ward, P. & Zahavi, A. The importance of certain assemblages of birds as “information-centres” for food-finding. Ibis (London 1859) 115, 517–534 (1973).
Weimerskirch, H., Bertrand, S., Silva, J., Marques, J. C. & Goya, E. Use of social information in seabirds: Compass rafts indicate the heading of food patches. PLoS ONE 5, e9928 (2010).
Ainley, D. G. et al. Geographic structure of Adélie penguin populations: Overlap in colony-specific foraging areas. Ecol. Monogr. 74, 159–178 (2004).
Fretwell, S. D. & Lucas, H. L. J. On territorial behavior and other factors influencing habitat distribution in birds. Acta Biotheor. 19, 16–36 (1970).
Acknowledgements
This study was funded by the ICE Centre of the Norwegian Polar Institute, the Fonds de recherche du Québec—Nature et technologies (FRQNT), Natural Sciences and Engineering Research Council of Canada (NSERC), MITACS (Globalink), the Norwegian Polar Institute MOSJ program, (www.mosj.no), the SEAPOP program (www.seapop.no). Olivier Chastel and Pierre Blévin were funded by the French Polar Institute (IPEV, Prog 330). We are thankful to Maite Cerezo Araujo, Delphin Ruche and Saga Svavarsdóttir, and the Norwegian Polar Institute's logistic department for their support in the field. We also want to thank Christen H. Fleming for his assistance with the kernel analyses, and Kyle Elliott and Kimberly Malcolm for their insights on previous versions of this manuscript.
Author information
Authors and Affiliations
Contributions
P.Be., S.D., J.B., H.Stø. and H.Ste. conceived the study; P.Be. wrote the manuscript with contributions from S.D. and J.B. P.Be. analyzed data with help from N.G.Y., S.H. and M.J.F.; P.Be., S.D., P.B.L., O.C. collected the tracking data and G.M., J.K. and J.M. geoprocessed glacier front locations; all co-authors commented on the first drafts of the study.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Bertrand, P., Bêty, J., Yoccoz, N.G. et al. Fine-scale spatial segregation in a pelagic seabird driven by differential use of tidewater glacier fronts. Sci Rep 11, 22109 (2021). https://doi.org/10.1038/s41598-021-01404-1
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-021-01404-1
This article is cited by
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
