
Abstract
We here report a new elateroid, Anoeuma lawrencei Li, Kundrata and Cai gen. et sp. nov., from mid-Cretaceous Burmese amber. Though superficially similar to some soft-bodied archostematans, Anoeuma could be firmly placed in the polyphagan superfamily Elateroidea based on the hind wing venation. Detailed morphological comparisons between extant elateroids and the Cretaceous fossils suggest that the unique character combination does not fit with confidence into any existing soft-bodied elateroid group, although some characters indicate possible relationships between Anoeuma and Omalisinae. Our discovery of this new lineage further demonstrates the past diversity and morphological disparity of soft-bodied elateroids.
Similar content being viewed by others

Introduction
The beetle superfamily Elateroidea is one of the major and oldest polyphagan groups1,2,3. It contains a broad spectrum of forms; from the lineages with a hard body, clicking mechanism, and five abdominal ventrites of which at least some are connate (e.g., the click-beetles) through variously intermediate groups (e.g., Brachypsectridae and Jurasaidae) to the lineages with an extremely soft body with some morphological reductions, and seven to eight free abdominal ventrites connected with extensive membranes1,4,5. Historically, soft-bodied elateroids were thought to form a monophyletic group called cantharoids6. Recent molecular studies, however, definitely rejected the monophyly of Cantharoidea1,2,3.
Although the soft-bodiedness is known in several unrelated beetle groups, it is by far most widespread and extensively studied in the superfamily Elateroidea. Soft-bodiedness is hypothesised to be connected with the neoteny, which is probably resulted from the termination or modification of complete metamorphosis7. The underlying molecular and developmental mechanisms have not been yet explored for elateroids, but studies in other insects suggested that the changes in a few components in hormone signaling pathway could substantially alter the developmental process and lead to neoteny8,9,10. Although there might be some deep homology in soft-bodied elateroids, different lineages still exhibit diverse morphology and may modify the developmental scheme in various ways11. Thus elateroids show a high morphological diversity and developmental plasticity.
Due to the multiple parallelly-evolved morphological traits as a result of incomplete sclerotisation and neoteny in soft-bodied elateroids1,4,12, it is almost impossible to properly assess the interrelationships among elateroid families by morphological phylogenetic analyses11,13. Even in light of more informative molecular data, the relationships among currently recognised elateroid groups remain unstable1,2,3,5,12,14,15,16.
Recent years have witnessed the discoveries of several elateroid families of both extant5,15,17,18 and extinct forms11,19, suggesting a large portion of unknown diversity in Elateroidea. Here, we report a new soft-bodied elateroid from the mid-Cretaceous Burmese amber, which exhibits a unique character combination within the superfamily. We discuss morphology of the newly discovered fossil and compare it to other soft-bodied elateroids.
Systematic palaeontology
-
Order Coleoptera Linnaeus, 1758.
-
Suborder Polyphaga Emery, 1886.
-
Superfamily Elateroidea Leach, 1815.
-
Family incertae sedis.
-
Genus Anoeuma Li, Kundrata & Cai gen. nov.
-
urn:lsid:zoobank.org:act:5F05C3D4-2CFA-4DF5-8B0C-CCF4E35BF708.
Type species
Anoeuma lawrencei sp. nov., here designated.
Etymology
The generic name is an anagram of Euanoma, a genus in Elateridae: Omalisinae, in reference of the morphological similarity shared by the new genus and the Omalisinae. Gender: feminine.
Diagnosis
Adult male
Head distinctly hypognathous. Antennae subfiliform, with 11 antennomeres; antennomeres 2 and 3 short. Mandibles unidentate, slender, sickle-shaped. Maxillary palps 4-segmented, with apical palpomere distinctly elongate. Labial palps 3-segmented. Tentorial pits absent. Prosternum in front of coxae subtriangular, longer than diameter of procoxal cavity. Protrochantins large, triangular and with slender process attached to procoxa. Mesocoxae narrowly separated. Elytra short, not completely covering abdomen. Hind wings fully developed, with radial cell closed. Tibial spurs distinct, double. Tarsi simpe, tarsomeres 2–4 relatively short and stout. Abdomen with eight free ventrites.
Composition and distribution
Monotypic, with Anoeuma lawrencei Li, Kundrata & Cai sp. nov. from the Burmese amber (northern Myanmar).
-
Anoeuma lawrencei Li, Kundrata & Cai sp. nov.
-
urn:lsid:zoobank.org:act:9674500D-16E6-4009-A499-3CE0ECBCDB48.
-
(Figs. 1, 2, 3, 4, 5; Supplementary Figs. S1, S2).
Figure 1 
General habitus of Anoeuma lawrencei gen. et sp. nov., holotype, NIGP175109, under incident light. (A) Dorsal view. (B) Ventral view. Scale bars: 500 μm.
Figure 2 
General habitus of Anoeuma lawrencei gen. et sp. nov., paratypes, under incident light. (A) NIGP175110, dorsal view. (B) NIGP175110, ventral view. (C) NIGP175111, dorsal view. Scale bars: 1 mm.
Figure 3 
Details of Anoeuma lawrencei gen. et sp. nov., holotype, NIGP175109, under widefield fluorescence. (A) Head and prothorax, dorsal view. (B) Elytra, dorsal view. (C) Hind wings and abdomen, dorsal view. (D) Head and prothorax, ventral view. (E) Meso- and metathorax, ventral view, with arrowheads showing the double tibial spurs. (F) Abdomen, ventral view. Abbreviations: el, elytron; msv, mesoventrite; mtv, metaventrite; pn, pronotum; ps, prosternum; v2, ventrite 2. Scale bars: 300 μm.
Figure 4 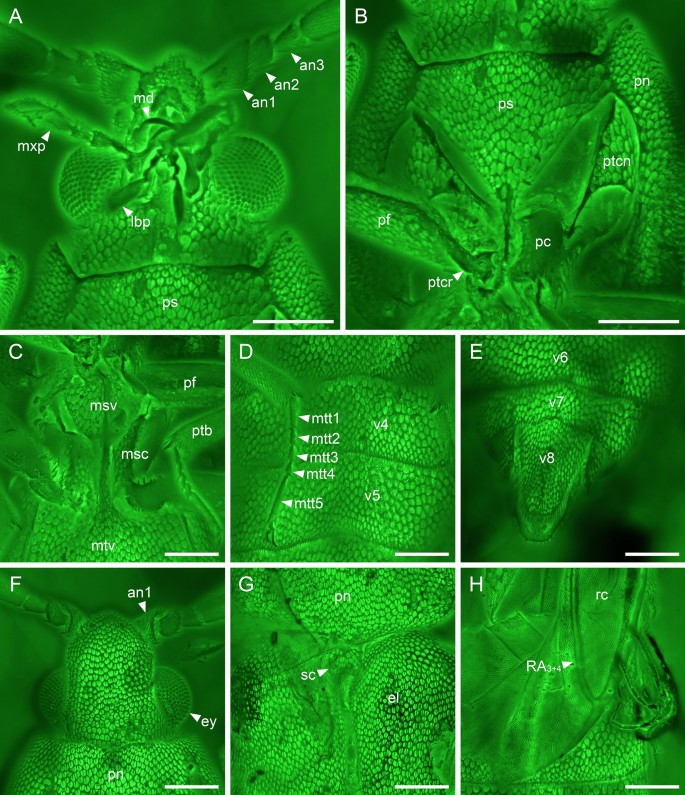
Details of Anoeuma lawrencei gen. et sp. nov., paratype, NIGP175110, under confocal microscopy. (A) Head, ventral view. (B) Prothorax, ventral view. (C) Mesothorax, ventral view. (D) Middle portion of abdomen, ventral view. (E) Abdominal apex, ventral view. (F) Head, dorsal view. (F) Elytral base, dorsal view. (F) Part of hind wing, dorsal view. Abbreviations: an1–3, antennomeres 1–3; el, elytron; ey, compound eye; lbp, labial palp; md, mandible; msc, mesocoxa; mtt1–5, metatarsomeres 1–5; mtv, metaventrite; mxp, maxillary palp; pc, procoxa; pf, profemur; pn, pronotum; ps, prosternum; ptb, protibia; ptcn, protrochantin; ptcr, protrochanter; rc, radial cell; sc, scutellum; v4–8, ventrites 4–8. Scale bars: 200 μm.
Figure 5 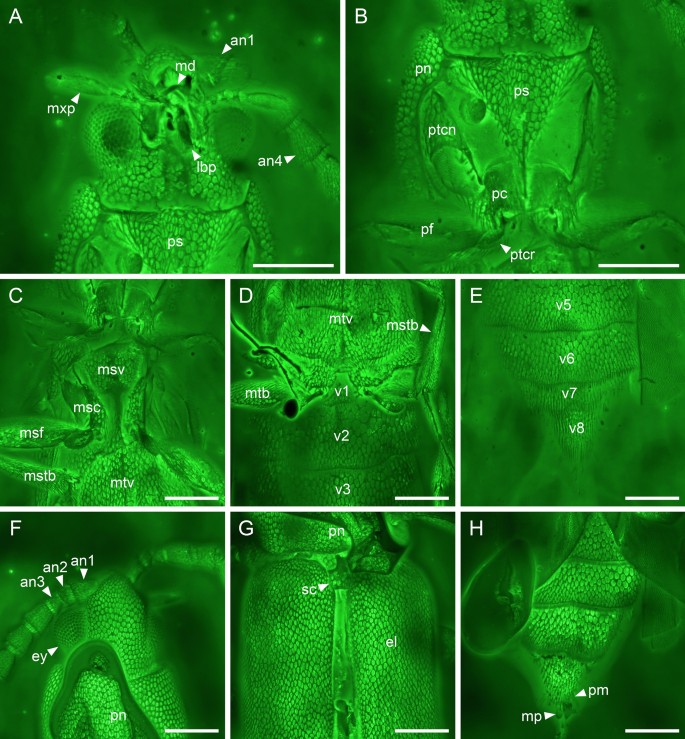
Details of Anoeuma lawrencei gen. et sp. nov., holotype, NIGP175109, under confocal microscopy. (A) Head, ventral view. (B) Prothorax, ventral view. (C) Mesothorax, ventral view. (D) Abdominal base, ventral view. (E) Abdominal apex, ventral view. (F) Head, dorsal view. (F) Elytral base, dorsal view. (F) Abdominal apex, dorsal view. Abbreviations: an1–4, antennomeres 1–4; el, elytron; ey, compound eye; lbp, labial palp; md, mandible; mp, median piece; msc, mesocoxa; msf, mesofemur; mstb, mesotibia; mtb, metatibia; mtv, metaventrite; mxp, maxillary palp; pc, procoxa; pf, profemur; pm, paramere; pn, pronotum; ps, prosternum; ptcn, protrochantin; ptcr, protrochanter; rc, radial cell; sc, scutellum; v1–8, ventrites 1–8. Scale bars: 200 μm.





Etymology
The specific name is a patronym in honor of Dr. John F. Lawrence, an internationally recognised coleopterist.
Type materials
Holotype, NIGP175109, male (NIGP). Five paratypes, males, NIGP175110 (NIGP), NIGP175111 (NIGP), NM-T3471 (NMPC, ex coll. PCRK), BUR003 (PCRK), and BUR004 (PCRK).
Locality and horizon
Amber mine located near Noije Bum Village, Tanai Township, Myitkyina District, Kachin State, Myanmar; unnamed horizon, mid-Cretaceous, Upper Albian to Lower Cenomanian20,21.
Diagnosis
As for the genus (vide supra).
Description
Adult male
Body weakly sclerotised, elongate, about 4.6–4.9 times as long as wide at humeri, densely punctate and setose.
Head (Figs. 4A,F,5A,F) distinctly hypognathous, almost fully exposed, widest basally, 1.1–1.2 times as long as wide (not including eyes), including eyes almost as wide as anterior margin of pronotum; portion in front of eyes distinctly narrowing anteriad; dorsal surface flat, without protuberance or depression. Compound eyes moderately large and protruding, finely facetted, without interfacetal setae, well separated both dorsally and ventrally. Antennal insertions located fronto-laterally, separated by approximately the maximum width of antennomere 1. Subantennal grooves absent. Antennae with 11 antennomeres, reaching basal third of elytra when directed posteriorly, finely setose; antennomere 1 moderately wide, broadest apically; antennomeres 2 and 3 short and ring-like, subequal in length; antennomere 3 slightly wider than antennomere 2; antennomeres 4–10 moderately elongate, broadest apically; antennomere 11 moderately elongate, fusiform. Ratio of antennomere lengths: ~ 1.8:0.9:1.0:2.2:2.1:2.2:2.2:2.2:2.3:2.2:2.9. Labrum plate-like, widely rounded apically. Mandibles narrow and slender, sickle-shaped, gradually curved mesally with sharply acute apices. Maxillary palps 4-segmented; apical palpomere elongate, about 3.3 times as long as penultimate one. Labial palps 3-segmented; apical palpomere relatively elongate, more than three times as long as penultimate one. Posterior tentorial pits absent.
Pronotum (Fig. 3A) subquadrate, 1.1 times as wide as long; pronotal disc with surface flat, without protuberance or depressions; anterior margin widely concave; anterior angles slightly produced forward; posterior angles rounded, not produced; lateral protonal carinae probably present. Scutellar shield small, transverse, anterior and posterior edges slightly emarginate. Prosternum (Figs. 4B,5B) in front of coxae subtriangular, longer than diameter of procoxal cavity; prosternal process developed, extremely slender, reaching middle of procoxae. Pronotosternal sutures short, slightly curved. Protrochantins (Figs. 4B,5B) large, triangular and with slender process attached to procoxa. Procoxae suboval, slightly emarginate at contact point with protrochantin, contiguous, not strongly projecting. Mesoventrite (Figs. 4C,5C) well-developed, sclerotised; anterior edge not emarginate. Mesocoxae elongate, narrowly separated. Suture between mesoventral process and anterior process of metaventrite invisible. Metaventrite (Fig. 3E) large, with almost complete discrimen; metakatepisternal suture absent. Metacoxae (Fig. 5D) transverse, narrowly separated.
Elytra (Fig. 3B) relatively short, covering only about half of abdomen, together 1.7 times as long as wide, 2.1–2.3 times as long as pronotum; surface irregularly punctate, without carinae; elytral apex rounded. Hind wings (Figs. 2C,3C,4H) fully developed; radial cell closed, not bordered by cross-veins; RP branches not present; RP fused with MP1+2 distally.
Legs slender. Trochanters obliquely articulated to femoral bases. Femurs broadest medially. Tibiae each with two distinct spurs. Tarsal formula 5–5-5; all tarsomeres simple, tarsomeres 2–4 relatively short and stout; tarsomere 5 elongate, distinctly longest (Fig. 4D). Claws simple, slightly curved.
Abdomen with eight free ventrites (Figs. 3F,4D,E,5D,E); penultimate ventrite distinctly emarginate medially. Aedeagus trilobate, not fully exposed; median lobe narrow subapically, then gradually widened and rounded apically; parameres with tips simple, narrowly rounded, with long setae (Figs. 1,5H).
Measurements
NIGP175109 (holotype): BL (body length) 3.19 mm, BW (body width) 0.69 mm, HL (head length) 0.41 mm, HW (head width) 0.35 mm, PL 0.50 mm (pronotal length), PW 0.53 mm (pronotal width), EL 1.16 mm (elytral length). NIGP175110 (paratype): BL 4.52 mm, BW 0.92 mm, HL 0.52 mm, HW 0.48 mm, PL 0.75 mm, PW 0.84 mm, EL 1.59 mm. NIGP175111 (paratype): BL 2.77 mm. NM-T3471 (paratype): BL 3.9 mm. BUR003 (paratype): BL 2.8 mm. BUR004 (paratype): BL 4.4 mm.
Discussion
Subordinal and superfamilial placement of Anoeuma gen. nov.
The here presented fossil shares a rather typical habitus of soft-bodied elateroids. However, there are some characters which can be confusing if one tries to classify this lineage within the Coleoptera. The most intriguing is the ventral prothoracic structure which might seem to be unusual for Polyphaga, including Elateroidea. In Polyphaga, the propleuron is internallised as a cryptopleuron, while in the other three beetle suborders the propleuron remains external22. In Anoeuma gen. nov., the sclerite between prosternum and hypomeron (Figs. 4B,5B) could be potentially interpreted as a propleuron. If we follow this scenario, the only reasonable placement for the fossil would be in Archostemata, probably close to an aberrant soft-bodied Micromalthus LeConte, 1878. The new fossil shares a series of characters with Micromalthidae, including e.g., the absence of dorsal head protuberances (Figs. 4F,5F), the shortened elytra (Fig. 3B), and the higher number of abdominal ventrites (Fig. 3F)23. However, many other characters do not support a position of Anoeuma gen. nov. near Micromalthidae at all. For example, Micromalthidae possess mandibles with three vertically arranged teeth, which is an apomorphy of the Micromalthidae + Ommatidae clade24,25, while Anoeuma gen. nov. has simple mandibles (Figs. 4A,5A). When we look at the hind wing, the folding pattern present in the fossil may look untypical for the polyphagan beetles; however, its hind wing venation looks elateroid-like. The radial cell of Archostemata is bordered by the radial bar, R3+4, and two cross-veins, while in Elateroidea (and Polyphaga in general, and also Anoeuma gen. nov.) the radial cell is bordered by the radial bar and R3+4 only, with no cross-veins (Figs. 2C,3C)26,27. Further, in Archostemata, the RP branches are relatively well developed, and RP and MP1+2 are connected by two cross-veins, while in Elateroidea (and also in Anoeuma gen. nov.), the RP branches are usually completely lost, and RP fuses with MP1+2 distally (Figs. 2C,3C). Therefore, we believe the newly discovered fossil should be classified in Elateroidea rather than in Archostemata. The sclerite between prosternum and hypomeron could be interpreted as a well-developed protrochantin (Figs. 4B,5B). Indeed, a similarly shaped protrochantin (i.e., triangular with a slender process attached to procoxa) can be found in some other soft-bodied elateroids (e.g., 15).
Position of Anoeuma gen. nov. within Elateroidea
The soft-bodiedness and neoteny originated multiple times within the Elateroidea1,5,7. The classification of soft-bodied elateroids based solely on morphology is precluded by the fact that many related groups are differently affected by neoteny, thus being not similar, but on the other hand, some lineages look superficially very similar although they are only distantly related1. The morphology-based phylogenetic analyses failed to provide us with the natural classification of the Elateroidea13 or to determine the phylogenetic position of a recently described fossil softbodied lineage11. Thus, since we cannot reliably use the morphological characters to test the position of Anoeuma gen. nov. within the Elateroidea, and the DNA, as an another potential source of data, is not available for the Mesozoic fossil taxa28,29, we have to rely on the comparison of the newly discovered fossil with other soft-bodied elateroids.
There are several groups within the soft-bodied elateroids which can be easily ruled out as potential relatives of Anoeuma gen. nov. due to their prognathous mouthparts (strongly hypognathous in Anoeuma gen. nov.) in combination with various other characters, especially the highly reduced and strongly transverse prosternum (subtriangular and rather long in Anoeuma gen. nov.). Such groups include the recently described Jurasaidae5, Omethidae (including Telegeusinae)30,31,32, and Phengodidae (including Penicillophorini)33,34,35. Rhagophthalmidae and the recently described fossil Cretophengodidae clearly differ from Anoeuma gen. nov. in having antennae with 12 antennomeres11,36 (11 antennomeres in Anoeuma gen. nov.). Drilini (Elateridae: Agrypninae) have only antennomere 2 short, and have bidentate mandibles and setae on outer basal portions of pretarsal claws4 (antennomeres 2 and 3 subequal in length, mandibles unidendate, and pretarsal claws without setae in Anoeuma gen. nov.). Cantharidae have usually prognathous head, contiguous mesocoxae, tarsomere 4 expanded and ventrally bilobed37 (head hypognathous, mesocoxae separated, and tarsomere 4 simple in Anoeuma gen. nov.). Lampyridae have head at least partly covered by pronotum, eyes ususally occupying most of the head, prosternum in front of coxae distinctly transverse, elytra usually not reduced, hind wings absent when elytra reduced, and tibial spurs usually absent or indistinct38,39 (head not concealed by pronotum, eyes smaller, prosternum distinctly longer, elytra reduced but with fully-developed hind wings, and tibial spurs present and distinct in Anoeuma gen. nov.). Lycidae have mostly prognathous mouthparts but the neotenic groups have hypognathous head with reduced mouthparts similar to Anoeuma gen. nov. However, lycids differ from Anoeuma gen. nov. in much more reduced, distinctly transverse prosternum, relatively widely separated mesocoxae, and hind wings dramatically reduced when elytra reduced40,41.
The here described fossil is most similar in habitus to Penicillophorini (Phengodidae), Telegeusinae (Omethidae), Iberobaeniidae, and Omalisinae (Elateridae). Although the Penicillophorini additionally share with Anoeuma gen. nov. the antennae without rami, tiny antennomeres 2 and 3 (most genera), and the reduced elytra with hind wings well-developed42,43,44,45, and Telegeusinae share with with Anoeuma gen. nov. more or less elongate apical maxillary palpomere, short tarsomeres 1–4, and reduced elytra with hind wings well-developed31,32,46, both groups differ in the above mentioned characters typical for Phengodidae and Omethidae, respectively. Recently discovered monogeneric Iberobaeniidae share with Anoeuma gen. nov. the shape of head, hypognathous mouthparts, and short tarsomeres 1–4. However, Iberobaeniidae can be distingushed by 2-segmented labial palps, pronotum without lateral carinae, not apparently shortened elytra, and much more reduced hind wing venation, with e.g., radial cell vestigial)15,47. Representatives of the soft-bodied click-beetle subfamily Omalisinae share with Anoeuma gen. nov. the general appearance, the slightly serrate antennae, tiny and ring-like antennomeres 2 and 3, and at least some have also a relatively long prosternum (although differently shaped) and a more developed mesoventrite (e.g., Cimbrion Kazantsev48). Anoeuma gen. nov. is superficially most similar to Paradrilus Kiesenwetter in the prolonged frontal part of the cranium with (fronto)laterally inserted antennae, hypognathous mouthparts, and the pronotum without well-defined posterior angles (other omalisine genera have better developed posterior angles)49. Nevertheless, the fossil genus differs from Omalisinae in having the apparently narrower frontoclypeal part and even more hypognathous mouthparts, subtriangular prosternum with extremely slender prosternal process (prosternum transverse and without prosternal process or clearly subrectangular in Omalisinae), reduced elytra (all known Omalisinae have elytra long, covering whole or most of abdomen), tarsomeres 2–4 short and distinctly shorter than last tarsomere (Omalisinae usually have elongate tasomeres, with only tarsomere 4 relatively shorter, and last tarsomere never distinctly longer), and the elongate terminal maxillary palpomere, which is much longer than preceding ones (terminal palpomere usually only slightly longer than preceding one and never so long in Omalisinae). Therefore, although Anoeuma gen. nov. is in some characters more similar to Omalisinae than to other elateroid groups, and some of the characters which distinguish them are highly plastic in some soft-bodied elateroid lineages (e.g., 50), we prefer to place the newly discovered fossil in Elateroidea incertae sedis. Discoveries of further taxa related to Anoeuma gen. nov. in future would probably help us to better understand the systematic placement of this taxon, which we hypothesise may either represent a separate evolutionary lineage on a family rank or is related to the soft-bodied omalisine click-beetles.
Conclusions
Anoeuma gen. nov. exhibits a mixture of characters known in several neotenic elateroid lineages, but on the other hand, it differs from them in other taxonomically important characters. Additionally, the morphology of ventral prothorax and the dense body punctation are somewhat unique among soft-bodied Elateroidea. In the current state of knowledge, we are unable to place Anoeuma gen. nov. into any existing family in full confidence. However, since we refrain here from erection of a new suprageneric rank for Anoeuma gen. nov., we tentatively classify it as Elateroidea incertae sedis. Neotenics are rarely found not only in the fossil record but also recently in the field, mostly due to their cryptic lives. The flight-capable males usually fly only reluctantly and spend most of the day hidden in the soil or other substrates, and larviform females do not move much at all5,47,51. Therefore, every report of a new neotenic lineage is extremely important for understanding the past diversity and morphological variability of soft-bodied elateroids. Since the males of Anoeuma gen. nov. exhibit a number of characters which are typical for the lineages where the females are strongly affected by the paedomorphic development5,41, we expect that the females, when/if discovered, will be flightless and larviform.
Materials and methods
Materials
The Burmese amber specimens studied here were derived from amber mines near Noije Bum (26°20' N, 96°36' E), Hukawng Valley, Kachin State, northern Myanmar. The specimens are deposited in the Nanjing Institute of Geology and Palaeontology, Chinese Academy of Sciences, Nanjing, China (NIGP), the Department of Palaeontology of the National Museum, Prague, Czech Republic (NMPC), and the collection of R. Kundrata, Olomouc, Czech Republic (PCRK). The amber pieces were trimmed with a small table saw, ground with emery papers of different grit sizes, and finally polished with polishing powder.
Fossil imaging
Photographs under incident light were mainly taken with a Zeiss Discovery V20 stereo microscope. Widefield fluorescence images were captured with a Zeiss Axio Imager 2 light microscope combined with a fluorescence imaging system. Confocal images were obtained with a Zeiss LSM710 confocal laser scanning microscope, using the 488 nm Argon laser excitation line52. Images under incident light and widefield fluorescence were stacked in Helicon Focus 7.0.2 or Zerene Stacker 1.04. Confocal images were stacked in Helicon Focus 7.0.2. Images were further processed in Adobe Photoshop CC to enhance contrast.
Nomenclatural acts
This published work and the nomenclatural acts it contains have been registered in ZooBank. The LSID for this publication is urn:lsid:zoobank.org:pub:B36118CC-136B-4FCD-B320-B4EB04D0EFB9.
Data availability
The original confocal data are available in Zenodo repository (https://doi.org/10.5281/zenodo.5553492).
References
Kundrata, R., Bocakova, M. & Bocak, L. The comprehensive phylogeny of the superfamily Elateroidea (Coleoptera: Elateriformia). Mol. Phylogenet. Evol. 76, 162–171 (2014).
Zhang, S.-Q. et al. Evolutionary history of Coleoptera revealed by extensive sampling of genes and species. Nat. Commun. 9, 205 (2018).
McKenna, D. D. et al. The evolution and genomic basis of beetle diversity. Proc. Natl. Acad. Sci. 116, 24729–24737 (2019).
Kundrata, R. & Bocak, L. Molecular phylogeny reveals the gradual evolutionary transition to soft-bodiedness in click-beetles and identifies Sub-Saharan Africa as a cradle of diversity for Drilini (Coleoptera: Elateridae). Zool. J. Linn. Soc. 187, 413–452 (2019).
Rosa, S. P., Costa, C., Kramp, K. & Kundrata, R. Hidden diversity in the Brazilian Atlantic rainforest: The discovery of Jurasaidae, a new beetle family (Coleoptera, Elateroidea) with neotenic females. Sci. Rep. 10, 1544 (2020).
Lawrence, J. F. Rhinorhipidae, a new beetle family from Australia, with comments on the phylogeny of the Elateriformia. Invertebr. Taxon. 2, 1–53 (1987).
Kundrata, R. & Bocak, L. The phylogeny and limits of Elateridae (Insecta, Coleoptera): Is there a common tendency of click beetles to soft-bodiedness and neoteny?. Zool. Scr. 40, 364–378 (2011).
Chafino, S. et al. Differential expression of the adult specifier E93 in the strepsipteran Xenos vesparum Rossi suggests a role in female neoteny. Sci. Rep. 8, 14176 (2018).
Vea, I. M. & Minakuchi, C. Atypical insects: Molecular mechanisms of unusual life history strategies. Curr. Opin. Insect Sci. 43, 46–53 (2021).
Vea, I. M. et al. E93 expression and links to the juvenile hormone in hemipteran mealybugs with insights on female neoteny. Insect Biochem. Mol. Biol. 104, 65–72 (2019).
Li, Y.-D. et al. Cretophengodidae, a new Cretaceous beetle family, sheds light on the evolution of bioluminescence. Proc. R. Soc. B: Biol. Sci. 288, 20202730 (2021).
Bocak, L., Motyka, M., Bocek, M. & Bocakova, M. Incomplete sclerotization and phylogeny: The phylogenetic classification of Plastocerus (Coleoptera: Elateroidea). PLoS ONE 13, e0194026 (2018).
Lawrence, J. F. et al. Phylogeny of the Coleoptera based on morphological characters of adults and larvae. Ann. Zool. 61, 1–217 (2011).
Mckenna, D. D. et al. The beetle tree of life reveals that Coleoptera survived end-Permian mass extinction to diversify during the Cretaceous terrestrial revolution. Syst. Entomol. 40, 835–880 (2015).
Bocak, L., Kundrata, R., Andújar Fernández, C. & Vogler, A. The discovery of Iberobaeniidae (Coleoptera: Elateroidea): A new family of beetles from Spain, with immatures detected by environmental DNA sequencing. Proc. R. Soc. B: Biol. Sci. 283, 20152350 (2016).
Douglas, H. B. et al. Anchored phylogenomics, evolution and systematics of Elateridae: Are all bioluminescent Elateroidea derived click beetles?. Biology 10, 451 (2021).
Bi, W.-X., He, J.-W., Chen, C.-C., Kundrata, R. & Li, X.-Y. Sinopyrophorinae, a new subfamily of Elateridae (Coleoptera, Elateroidea) with the first record of a luminous click beetle in Asia and evidence for multiple origins of bioluminescence in Elateridae. ZooKeys 864, 79–97 (2019).
Kusy, D. et al. Phylogenomic relationships of bioluminescent elateroids define the ‘lampyroid’clade with clicking Sinopyrophoridae as its earliest member. Syst. Entomol. 46, 111–123 (2021).
Peris, D. et al. Unlocking the mystery of the mid-Cretaceous Mysteriomorphidae (Coleoptera: Elateroidea) and modalities in transiting from gymnosperms to angiosperms. Sci. Rep. 10, 16854 (2020).
Shi, G. et al. Age constraint on Burmese amber based on U-Pb dating of zircons. Cretac. Res. 37, 155–163 (2012).
Mao, Y. et al. Various amberground marine animals on Burmese amber with discussions on its age. Palaeoentomology 1, 91–103 (2018).
Qvarnström, M. et al. Exceptionally preserved beetles in a Triassic coprolite of putative dinosauriform origin. Curr. Biol. 31(15), 3374–3381 (2021).
Hörnschemeyer, T. In Handbook of Zoology, Arthropoda: Insecta, Coleoptera, beetles, Vol. 1: Morphology and Systematics (Archostemata, Adephaga, Myxophaga, Polyphaga partim) 2nd edn (eds Beutel, R. G. & Leschen, R. A. B.) 48–52 (Walter de Gruyter, 2016).
Hörnschemeyer, T. The species-level phylogeny of archostematan beetles—Where do Micromalthus debilis and Crowsoniella relicta belong?. Syst. Entomol. 34, 533–558 (2009).
Li, Y.-D., Yamamoto, S., Huang, D.-Y. & Cai, C.-Y. A miniaturized ommatid beetle in mid-Cretaceous Burmese amber (Coleoptera: Archostemata: Ommatidae). Papéis Avulsos de Zoologia 60, e20206063 (2020).
Kukalová-Peck, J. & Lawrence, J. F. Evolution of the hind wing in Coleoptera. Can. Entomol. 125, 181–258 (1993).
Lawrence, J. F., Zhou, Y.-L., Lemann, C., Sinclair, B. & Ślipiński, A. The Hind Wing of Coleoptera (Insecta): Morphology nomenclature and phylogenetic significance. Part 1. General discussion and Archostemata–Elateroidea. Ann. Zool. 71, 421–606 (2021).
Allentoft, M. E. et al. The half-life of DNA in bone: Measuring decay kinetics in 158 dated fossils. Proc. R. Soc. B: Biol. Sci. 279, 4724–4733 (2012).
Penney, D. et al. Absence of ancient DNA in sub-fossil insect inclusions preserved in ‘Anthropocene’ Colombian copal. PLoS ONE 8, e73150 (2013).
Ramsdale, A. S. In Handbook of Zoology, Arthropoda: Insecta, Coleoptera, Beetles, Vol. 2: Morphology and Systematics (Elateroidea, Bostrichiformia, Cucujiformia partim) (eds Leschen, R. A. B. et al.) 149–153 (Walter de Gruyter, 2010).
Lawrence, J. F. In Handbook of Zoology, Arthropoda: Insecta, Coleoptera, Beetles, Vol. 2: Morphology and Systematics (Elateroidea, Bostrichiformia, Cucujiformia partim) (eds Leschen, R. A. B. et al.) 123–126 (Walter de Gruyter, 2010).
Roza, A. S., Constantin, R. & Mermudes, J. R. M. Pseudotelegeusis meloi sp. nov., the first Telegeusinae from Peru (Coleoptera: Omethidae, Telegeusinae). Eur. J. Taxon. 580, 1–13 (2019).
Costa, C. & Zaragoza-Caballero, S. In Handbook of Zoology, Arthropoda: Insecta, Coleoptera, Beetles, Vol. 2: Morphology and Systematics (Elateroidea, Bostrichiformia, Cucujiformia partim) (eds Leschen, R. A. B. et al.) 126–135 (Walter de Gruyter, 2010).
Zaragoza-Caballero, S. & Zurita-García, M. L. A preliminary study on the phylogeny of the family Phengodidae (Insecta: Coleoptera). Zootaxa 3947, 527–542 (2015).
Kundrata, R. et al. One less mystery in Coleoptera systematics: The position of Cydistinae (Elateriformia incertae sedis) resolved by multigene phylogenetic analysis. Zool. J. Linn. Soc. 187, 1259–1277 (2019).
Kawashima, I., Lawrence, J. F. & Branham, M. A. In Handbook of Zoology, Arthropoda: Insecta, Coleoptera, Beetles, Vol. 2: Morphology and Systematics (Elateroidea, Bostrichiformia, Cucujiformia partim) (eds Leschen, R. A. B. et al.) 135–140 (Walter de Gruyter, 2010).
Ramsdale, A. S. In Handbook of Zoology, Arthropoda: Insecta, Coleoptera, Beetles, Vol. 2: Morphology and Systematics (Elateroidea, Bostrichiformia, Cucujiformia partim) (eds Leschen, R. A. B. et al.) 153–162 (Walter de Gruyter, 2010).
Branham, M. A. In Handbook of Zoology, Arthropoda: Insecta, Coleoptera, Beetles, Vol. 2: Morphology and Systematics (Elateroidea, Bostrichiformia, Cucujiformia partim) (eds Leschen, R. A. B. et al.) 141–149 (Walter de Gruyter, 2010).
Martin, G. J., Branham, M. A., Silveira, L. F. & Bybee, S. M. Lampyrid-ID: Key to the world genera of fireflies (Coleoptera: Lampyridae). Version: May 2020, https://keys.lucidcentral.org/keys/v3/lampyridae/ (2020).
Bocak, L. & Bocakova, M. In Handbook of Zoology, Arthropoda: Insecta, Coleoptera, Beetles, Vol. 2: Morphology and Systematics (Elateroidea, Bostrichiformia, Cucujiformia partim) (eds Leschen, R. A. B. et al.) 114–123 (Walter de Gruyter, 2010).
Bocak, L., Grebennikov, V. V. & Masek, M. A new species of Dexoris (Coleoptera: Lycidae) and parallel evolution of brachyptery in the soft-bodied elateroid beetles. Zootaxa 3721, 495–500 (2013).
von Paulus, F. Penicillophorus ctenotarsus n. gen. et n. sp. aus Kolumbien, mit einer Beschreibung einer neuen Tribus Penicillophorini der Phengodidae (Col., Polyphaga, Cantharoidea). Zeitschrift der Arbeitsgemeinschaft Österreichischer Entomologen 25, 69–80 (1974).
Wittmer, W. Arbeiten zu einer Revision der Familie Phengodidae (Coloptera). Entomologische Arbeiten Museum G. Frey 27, 415–524 (1976).
Wittmer, W. Zur Kenntnis der Familie Phengodidae (Coleoptera). Mitteilungen Entomologischen der Entomologischen Gesellschaft Basel NF 31, 105–107 (1981).
Zaragoza-Caballero, S. D. géneros y tres especies nuevos de Penicillophorinae (Coleoptera: Phengodidae) de México. Revista Mexicana de Biodiversidad 79, 363–368 (2008).
Zaragoza-Caballero, S. & Rodriguez-Velez, B. Five new species of Telegeusis Horn, 1895 (Coleoptera: Telegeusidae) from Mexico, with a key to the species. Zootaxa 2917, 59–68 (2011).
Kundrata, R., Baena, M. & Bocak, L. Iberobaenia andujari sp. nov., the third species of Iberobaeniidae (Coleoptera: Elateroidea) from southern Spain. Ann. Zool. 67, 121–129 (2017).
Kundrata, R., Sormova, E. & Kakiopoulos, G. Revisiting the diversity of Euanoma Reitter, 1889 (Coleoptera: Omalisidae), with descriptions of four new species from Turkey. Ann. Zool. 68, 357–373 (2018).
Kundrata, R., Baena, M. & Bocak, L. Classification of Omalisidae based on molecular data and morphology, with description of Paradrilinae subfam. nov. (Coleoptera: Elateroidea). Zootaxa 3915, 413–422 (2015).
Ferreira, V. S., Keller, O. & Branham, M. A. Multilocus phylogeny support the nonbioluminescent firefly Chespirito as a new subfamily in the Lampyridae (Coleoptera: Elateroidea). Insect Syst. Diversity 4, 2 (2020).
Bocak, L., Bocakova, M., Hunt, T. & Vogler, A. P. Multiple ancient origins of neoteny in Lycidae (Coleoptera): Consequences for ecology and macroevolution. Proc. R. Soc. B: Biol. Sci. 275, 2015–2023 (2008).
Fu, Y.-Z., Li, Y.-D., Su, Y.-T., Cai, C.-Y. & Huang, D.-Y. Application of confocal laser scanning microscopy to the study of amber bioinclusions. Palaeoentomology 4, 266–278 (2021).
Acknowledgements
We are grateful to J.F. Lawrence (Australia), M.A. Ivie (USA), R.G. Beutel (Germany), M. Fikáček (Czech Republic), V.S. Ferreira (Brazil), and S.P. Rosa (Brazil) for their invaluable comments on the morphology of Anoeuma gen. nov. We thank Yan Fang for technical help in confocal imaging. Financial support was provided by the Second Tibetan Plateau Scientific Expedition and Research project (2019QZKK0706), the Strategic Priority Research Program of the Chinese Academy of Sciences (XDB26000000 and XDB18000000), the National Natural Science Foundation of China (41688103), and an internal grant from Palacky University (IGA_PrF_2021_019; RK, GP).
Author information
Authors and Affiliations
Contributions
C.-Y.C., R.K. and Y.-D.L. conceived the study. C.-Y.C. and R.K. processed the fossils. Y.-D.L, R.K. and G.P. acquired and processed the photomicrographs. Y.-D.L., R.K. and C.-Y.C. wrote the paper with contributions from the remaining authors.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Li, YD., Kundrata, R., Packova, G. et al. An unusual elateroid lineage from mid-Cretaceous Burmese amber (Coleoptera: Elateroidea). Sci Rep 11, 21985 (2021). https://doi.org/10.1038/s41598-021-01398-w
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-021-01398-w
This article is cited by
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
