
Abstract
Based on sexual selection theory, the reproductive potential of male primates is expected to be limited by access to fertile females. Alpha males, the highest ranking males in a social group, are predicted to have better access to mates and produce more offspring until they are no longer dominant, which usually corresponds with age. Little is known about male reproductive senescence independent of rank changes in nonhuman primates. Here, we examine variation in the reproductive success of high-ranking male rhesus macaques on Cayo Santiago. We recorded behavioral data for 21 adult males across 9 social groups during the 2013 mating season. Additionally, we used paternity data from the long-term database to determine the number of offspring each subject sired over his lifetime and during the study period. Older high-ranking males in stable groups had fewer offspring than younger high-ranking males in stable groups in 2013. The low reproductive output for the older males was not a result of lower mating effort, and reproductive output in 2013 was not predicted by total prior reproductive success. Our results provide novel evidence of post-copulatory reproductive senescence in high-ranking male nonhuman primates.
Similar content being viewed by others


Introduction
Sexual selection theory predicts that male primates are mainly limited in their reproductive potential by access to fertile females1. Within nonhuman primate groups, alpha males (the highest-ranking male in each group) have better access to mates2 and are therefore expected to produce more offspring until they are no longer dominant (e.g. baboons3; mandrills4), which is usually associated with older age due to the energetic and physical demands of maintaining dominance status5,6. This pattern varies greatly between primate species depending on their social structure and grouping pattern. Rhesus macaques live in large multi-male, multi-female groups where both males and females form strong linear dominance hierarchies in which high-ranking individuals have better access to resources, including mating opportunities.
Relatively little is known about male reproductive senescence, or age-related declines in reproduction, independent of rank changes in primates, although a large body of research literature on this topic exists for human males7,8,9,10,11,12,13. In men, age has been negatively associated with sperm quality and positively associated with a risk of genetic disorders in offspring9,12,14 (but see10). Importantly, studies on men do not show consistent changes in hormone concentrations with age, suggesting that hormone concentrations are not a good predictor of reproductive function and should not be used as a proxy measure in primates15,16.
In nonhuman primates, the focus has primarily been on female reproductive senescence (e.g. rhesus macaques17,18,19), and many studies have simply assumed that alpha males who are seen mating frequently are producing offspring, regardless of their age20. As studies have started incorporating paternity data, it has become clear that mating success does not always equal reproductive success21. A few studies have found evidence of changes in sexual behaviors with age in male nonhuman primates, but there is limited evidence of a decrease in fertility (reviewed in22). The ability to study reproductive senescence in free-ranging primates is limited by the confounding factor that many older males lose their high-ranking status prior to a decrease in their reproductive success5,23,24. For example, male baboons experience a decline in reproductive output as they age, but this decline coincides with a loss in dominance rank5. In these cases, a decrease in reproductive success appears to be associated with reproductive senescence in pre-copulatory mechanisms; however, it does not address potential senescence in post-copulatory mechanisms, such as ejaculate quality. Males that are able to maintain mating opportunities with females throughout their life may still experience senescence in the ability to fertilize an egg or produce a viable offspring25.
The population of rhesus macaques (Macaca mulatta) on Cayo Santiago, Puerto Rico26, provides a unique opportunity to examine the impact of age on male reproductive output for several reasons. First, the long-term database for this population includes the age of each individual and detailed records of how many offspring each male has sired over his lifetime. Second, the influence of socio-demographic variables on reproductive output can be assessed with minimal confounding ecological variation compared to studies of wild populations by comparing groups of different size and composition that live in the same ecological habitat. Finally, there is significant variation in the age of high-ranking males on the island, which may be facilitated by food provisioning that may provide opportunities for older males to maintain high-ranking positions more frequently than they would be able to under more ecologically stressful conditions. This pattern may be further facilitated by the succession rank system in rhesus macaques, in which males typically achieve high dominance status based on their tenure in a group27 (but see28) rather than by challenges to other males29.
Importantly, dominance hierarchies are not always stable and variation in group dynamics could also impact male reproductive success. Periods of social instability in nonhuman primate groups can have behavioral and physiological consequences for males in a dominance hierarchy30,31,32,33,34. These consequences could impact a male’s ability to successfully reproduce. Social instability has been associated with energetic stress in male primates (rhesus macaques33, baboons30,32), which could lower a male’s ability to compete for mates or produce viable offspring. Additionally, males may have to focus more time on aggressive interactions with other males instead of mating effort. For example, we have previously documented variation in this population in the use of behavioral displays—males in stable groups display in mating contexts more often than males in groups with unstable hierarchies and males in groups with unstable hierarchies instead display in aggressive or dominance interactions with other males34.
Here, we examine variation in the reproductive success of high-ranking male rhesus macaques on Cayo Santiago and the potential factors influencing this variation to determine if reproductive senescence is present in this species. We compare the number of offspring sired by high-ranking males in 2013 in relation to their age, rank, and the stability of their social group, as well as their mating effort. We also investigated how their reproductive output for the study year compared to their lifetime reproductive success prior to that year. Although we have data on the total number of surviving offspring males have sired over their lifetime, we do not have any behavioral data (including mating behaviors and assessments of dominance rank) outside of the study period. Therefore, we present detailed analyses of variation in male reproduction for the 2013 study period and compare this to what is known from the genetic database for the study subjects. Overall, we expected top ranking males in stable social groups (where their dominance rank did not change during the study) to produce more offspring than high-ranking males in groups with unstable hierarchies and that there would be a positive correlation between mating effort (days spent consorting, number of mounts, and number of ejaculations observed) and the number of offspring sired in 2013. We considered deviations from these patterns for older males as evidence of reproductive senescence. Specifically, we predicted that if reproductive senescence impacts male reproductive success in rhesus macaques, then (1) older high-ranking males of stable social groups would have lower reproductive output than younger high-ranking males in stable social groups, (2) older high-ranking males would have fewer offspring in recent years than over their lifetime, and (3) the mating behaviors of older males would not be correlated with the number of offspring produced in that year. Our results provide novel evidence of reproductive senescence in high-ranking male nonhuman primates.
Methods
Site and subjects
The island of Cayo Santiago in Puerto Rico is home to a colony of free-ranging rhesus macaques that was first established in 1938 with approximately 400 wild-caught animals from India35. At the time of this study, the population had grown to over 1,200 individuals, which had naturally divided themselves into 9 social groups. The monkeys are provisioned with water and commercially available monkey chow. Records of all births and deaths have been maintained since 1956 and all individuals on the island currently have been genotyped to determine both maternal and paternal lineage36.
As previously described34, the subjects of this research were 21 adult males that were the highest ranking individuals in the 9 social groups, as determined at the beginning of the study by outcome of fights, access to food, displacement events, fear grins, and avoidance behaviors37. Dominance rank was subsequently reassessed throughout the study through ad libitum observations, and five of the nine groups were considered unstable because of changes in the dominance ranks of the top-ranking males in these groups. Two of these unstable groups were small groups that had formed within the year prior to this study, stayed on the periphery of the larger group from which they had split-off, and were not considered independent from the larger group during parts of the study period. For this reason, only three males were studied from these two groups combined. These males were the highest ranking within their small groups and were therefore classified as top-ranking (because of their high rank within the newly formed groups), but none of the individuals were identified as alpha because they were submissive to the higher-ranking individuals in the larger group to which they were peripheral. The other three groups with unstable hierarchies had major changes in the rank of the highest ranking males throughout the study, resulting in rank changes for the alpha, beta (second highest-ranking males in a group), and gamma (third highest-ranking male in a social group) males in those groups. The other four groups had stable dominance hierarchies throughout the behavioural study period of 2013. The rank of an individual within a stable group or membership in an unstable group is being referred to as “rank stability” within this paper (see Supplementary Table 1 for further information about the study subjects). Due to the comparative nature of this study, including the need to gather detailed behavioral and reproductive data on each subject with subjects ranging across every group on the island, it was not possible to include more subordinate males. Given that high-ranking male primates generally have access to mating partners, prioritizing data collection from these individuals provided the best opportunity to measure variation in reproductive success for males who were actively engaging in mating behaviors. The investigation was approved by the IACUC of the University of Puerto Rico, Medical Sciences Campus (protocol No. A0100108). All methods were performed following the relevant regulations and guidelines.
Behavioural observations
Behavioural data were collected 5 days a week from approximately 0700 to 1430 during the 2013 mating season from mid-February to mid-July. The rhesus macaques on Cayo Santiago have distinct breeding and birth seasons, so behavioral observations were collected during the breeding season and data about the offspring produced during the birth season were gathered from the database. Each male was observed for a 10-min focal follow on a rotating basis at least every other day during the week. These follows were extended to 60 min if the male was engaged in a sexual consortship with a female. Consortships are prolonged associations between the male and female where the pair travels together, has high rates of affiliative behaviors, and mates during one or multiple mounting series. Scans were taken at the start of the focal and every 2 min after until the 10 min were complete or until the 60 min were complete. During consortships, all mating behaviors were recorded, including mounting and ejaculation.
Paternity data
Genetic paternity data were gathered from the long-term database managed by the Caribbean Primate Research Center. Through this database, we were able to determine the number of offspring each focal male sired in 2013, as well as how many offspring he sired prior to 2013. We used both the total counts of offspring up to 2013, as well as the average number of offspring per year during a male’s “adult” life by averaging the total number of offspring over the number of years older than four the male was. This method was to account for different ages, and used five as the earliest possible age of reproduction for males on Cayo Santiago, based on the earliest age of first reproduction for males in this population. Offspring paternity was determined when they were yearlings, meaning that infants who died prior to this sampling period would not have been included in the calculations. In 2013, there were 33 infant deaths and seven stillbirths. Infant mortality was 13% of all infants born that year. We cannot account for who sired these offspring.
Statistical analyses
Generalized linear mixed models fitted with a Poisson error structure were used to examine how male age, group stability, and the interaction of these two factors impacted the number of offspring produced, as well as the interaction between a male’s age and the number of days he spent consorting on the number of offspring he sired that year. We treated group ID as a categorical random effect in the model. To compare the number of offspring a male had sired prior to 2013 and in 2013 to their rank stability, we used Kruskal–Wallis tests. In these analyses, there were four categories of males: (1) alpha males in stable groups, (2) beta males in stable groups, (3) gamma males in stable groups, and (4) high-ranking males in groups with unstable hierarchies. The dataset included three males that were seven or eight years old, lived in groups with unstable hierarchies, and sired no offspring. This age category is the average age for first reproduction in male rhesus macaques, but many males do not produce their first offspring until after this age38. Given this pattern for age at first reproduction and that there were no males from the stable groups that were this young, we removed these three males from additional analyses on reproductive success. A Spearman correlation coefficient was calculated for male age (males > 8 years old) and number of offspring sired in 2013, as well as different indicators of male mating effort (days spent consorting, number of mounts, and number of ejaculations) and the number of offspring spired in 2013. All analyses were conducted in SAS version 9.4 (SAS Institute Inc., Cary, NC, USA). A p value < 0.05 was considered significant.
Results
Age and male reproduction in the study period
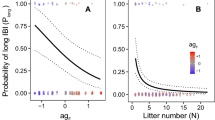
Using a generalized linear mixed model, we found that during the 2013 breeding season, group stability (p = 0.0101) and the interaction of group stability and male age (p = 0.0214) significantly accounted for the number of offspring produced that year (Table 1). In stable groups, older males had fewer offspring, but in unstable groups, the number of offspring did not decrease with age. Importantly, the oldest males in our study were all in the groups with stable dominance hierarchies. In our dataset, there were three males younger than age nine. These three males (ages seven and eight) were part of unstable groups and sired no offspring. Given the young age of these males and that there were no males from the stable groups that were this young, we removed these three males from additional analyses. Using the 18 remaining subjects that were nine years old and older, we found a significant negative correlation between age and the number of offspring sired in 2013 (ρ = -0.58, p = 0.0117). The alpha, beta, and gamma males in stable groups were driving this relationship while males in groups with unstable hierarchies had highly variable reproductive rates (Fig. 1). Importantly, all of the males in the groups with unstable hierarchies were age 15 and younger whereas high-ranking males in stable groups ranged in age from 10 to 21 years. Older males in stable groups had fewer infants, if any, in 2013 than the younger males in those groups (Fig. 1).

The age of high-ranking males living in stable groups (S) is negatively correlated with the number of offspring they sired during the 2013 breeding season, particularly after the age of 15, but this pattern does not apply to males in unstable hierarchies (U) between the ages of 9 and 15.
Older males (males > 15 years old) consorted with females on many of the days they were observed (Fig. 2). Across all males, there was a significant positive relationship between the number of days a male was observed consorting (ρ = 0.49, p = 0.0242; Fig. 3), the number of times they were observed mounting a female (ρ = 0.57, p = 0.0072), and the number of times they were seen ejaculating (ρ = 0.52, p = 0.0159) with the number of offspring sired in 2013, but deviations from this pattern were primarily the males over 15 years old from stable groups. Using a generalized linear mixed model, we found that both the number of days consorting (p = 0.0008) and the interaction between the number of days consorting and the male’s age (p = 0.0387) were significantly associated with variation in the number of offspring sired in 2013 (Table 2). This interaction is such that the amount of time a male spends consorting with females impacts the number of offspring they produce until a certain age when the likelihood of producing an offspring becomes very low regardless of mating effort.

Older males did not spend less time in consortships with females than other males. This graph shows the number of observation days a male was observed consorting with a female for each subject (N = 21). The black dots each represent one male, and the white dots each represent two males (i.e. there were two 11-year-olds that each were observed consorting on 14 days of the study period and two 18-year-olds who were each observed consorting for 15 days each). The oldest males in the study consorted with females on many of the days they were observed, as shown in this figure.

There was a positive association between the number of days males spent in consortships and the number of offspring they sired that year, but outliers included the older males from stable groups (the ages of these males over 15 years old are indicated). The LOESS (locally weighted smoothing) line created from the regression analysis uses a smoothing value of 1 and the 95% confidence limits are shown with the shading.
Two of the oldest males in this study (18 years and 21 years) were both alpha males of stable groups. They had produced the most offspring prior to 2013 out of all the focal males (52 and 47 offspring, respectively). They both were observed in consortships frequently with females on days they were observed (63% and 41% of observation days, respectively, compared to, for example, 60% and 48% for the two males who produced the most offspring in 2013 out of the study subjects). The 18-year-old had the highest number of observed ejaculations in the study. Yet, these males produced the fewest offspring (1 and 0, respectively) out of all the alpha males in the study. The other two alpha males of stable groups were 10 and 11 years old and produced 6 and 8 offspring, respectively, in 2013.
In groups with old alpha males, the ability to sire a high number of offspring in 2013 moved down the dominance hierarchy to other high-ranking males (Supplementary Table 1). The beta male in the group with the 18-year-old alpha male had 7 offspring. The gamma male in that group was 19 years old and did not produce any offspring. The beta male in the group with the 21-year-old alpha was also old (18 years) and also did not produce any offspring. However, the gamma male in that group was 12-years-old and had 8 offspring (tied with one of the alpha males for the most offspring produced in the study).
Male reproductive success prior to the study period
The total number of offspring a male sired leading up to the 2013 breeding season differed significantly based on the males’ rank and membership in a stable or unstable group (χ2 = 13.61; p = 0.0035) with alpha males of stable groups, in particular, having more offspring than males in groups with unstable hierarchies (Z = 2.91; p = 0.0188; Fig. 4A). To account for variation in age (and thus the number of years a male had to reproduce), we also compared the average number of offspring a male produced per year since the age of 5 and found the same pattern (χ2 = 10.03; p = 0.0183) with alpha males of stable groups, in particular, having more offspring than males in groups with unstable hierarchies (Z = 2.79; p = 0.0270). However, there was no difference in the number of offspring males of different rank categories sired during the 2013 breeding season (χ2 = 2.41; p = 0.4922; Fig. 4B). While we cannot account for variation in rank and mating effort in other years, we did also find that these older males with high lifetime reproductive success had declining reproductive output in the years immediately prior to this study (as they aged further away from 15). The 21-year-old alpha male had no offspring in both years prior to the study period—at ages 19 and 20. The 18-year-old alpha male that had one offspring during the study period also had one offspring per year at ages 16 and 17. The 18-year-old beta male had 2 offspring at age 16 and 1 offspring at age 17.

Alpha (1), beta (2), and gamma (3) males of stable groups and males in unstable hierarchies (4) differ in the total number of offspring they sired up to the 2013 mating season (A), but these rank categories did not predict reproductive success in the 2013 mating season (B). These boxplots show the variation in the mean (diamond) and median (line).
Discussion
Through this study of the reproductive output of high-ranking male rhesus macaques across all the groups on Cayo Santiago in 2013, we provide evidence of variation in male reproductive success across males of different rank stabilities and males of different ages. High-ranking male rhesus macaques in stable groups had high lifetime reproductive success prior to 2013. Given that male dominance ranks are not regularly assessed in every group on Cayo Santiago, we cannot comment on the directionality of that relationship or changes in rank status before or after the study period. High-ranking males in stable groups may be able to secure long tenures at top ranked position and therefore sire many offspring over the years or males that sire many offspring may be able to rise to high positions within the dominance hierarchy. Although we cannot assess the directionality of that relationship, it is clear that a male’s rank stability during the 2013 mating season was significantly associated with variation in the number of offspring that male had prior to 2013. For males under 16 years old, this pattern was maintained in 2013, with those high-ranking males in stable groups having high reproductive output; however, older males had significantly lower reproductive output, particularly those males aged 18 to 21 years. Males of this age make up a small percentage of the population on Cayo Santiago, with the life expectancy at birth of males being 14.4 years and the maximum lifespan of males being 29-years-old39. Hoffman et al.17 found a similar result for female rhesus macaques on Cayo Santiago with poor infant survival for older females, especially those who were 18 years and older.
Patterns of male rank and lifetime reproductive success in the high-ranking males on Cayo Santiago are consistent with findings from other species of primates (Papio cynocephalus21, Pan troglodytes verus40, Pan troglodytes schweinfurthii41) and in previous studies of this population42,43. The findings are also consistent with previous work showing a positive association between dominance rank and reproductive interactions with females around ovulatory periods in this population44. The patterns also fit with the predictions of the priority of access model, which posits that the number of offspring sired by a male will be influenced by the male’s rank, his access to females, and the number of competitors present2. The deviation from these patterns for the older high-ranking males in the group are important.
In other species, declines in reproductive output for older males have been associated with a loss of dominance status5,45, loss in attractiveness46, and/or loss in body condition5 leading to a decline in mating activity for older males5,45,46,47. But the older males in this study maintained high rates of mating behaviors while still experiencing the same decline in reproductive output. Additionally, previous studies showed that male body condition remained stable in adult male rhesus macaques (including those over the age of 15) even as reproductive output declined, with males between 17 and 24 years old producing no more than one infant per year38, and older males had high measures of immune function and low oxidative stress48. Furthermore, aging rhesus macaque males do not have erectile issues16.
Previous studies of reproductive success in older male macaques have had variable results. In Japanese macaques (Macaca fuscata), old males that were central in the study group did not sire any offspring49. Yet in Barbary macaques (Macaca sylvanus), there was no decline in reproduction for older males, with males aged 18 and older siring offspring50. In our study, we found that males aged 9 to 14 produced the most infants during the study period. Previous studies of the Cayo Santiago rhesus macaques found similar results with the highest number of offspring being sired by males that were 9 to 11 years old24 or 7 to 12 years old51 and very few offspring being produced by the oldest males in the study (18 and 19 years old51), but these studies did not show that age-related differences existed despite older males continuing to be high-ranking and having high rates of mating behaviors.
In previous studies of changes in rhesus macaque reproductive function over age, older males were found to exhibit fewer sexual behaviors, including mounting and ejaculation22; however, we found that in this study, older high-ranking males maintained a high rate of sexual activity. Older males that produced no surviving offspring or only one surviving offspring had comparable rates of consorting with females, mounting females, and ejaculating during mounts to their younger counterparts. Given the patterns of mating effort by these males and the evidence of previous fertility and reproductive success for these individuals, our results suggest that the low reproductive output of older high-ranking males is a result of post-copulatory reproductive senescence. These results add to a sparse literature on male post-copulatory reproductive senescence in free-ranging animals. Lemaître and Gaillard25 argue that this lack of age-specific data on reproductive success prevents research on the factors impacting reproductive senescence. Reed et al.52 found reduced reproductive output for male seabirds, common guillemots (Uria aalge), in the final year of life. Similar to our findings in this study, Wroblewski et al.41 found that older male (> 19 years old) chimpanzees were less successful at producing offspring than younger males (15 to 19 years old), even as the rank of those older males continued to rise. Our results provide some of the only evidence of lower reproductive output for older males despite maintaining high rates of mating behaviors. Additionally, our results indicate that behavioral observations of mating behaviors are not reliable indicators of male reproductive output and should not be used as such.
The relatively large testes of male rhesus macaques compared to their body size suggest that sperm competition53 is likely an important aspect of reproductive success in this species54,55. Female rhesus macaques consort with multiple males during an estrous cycle56,57. Additionally, sneaky copulations are a successful mating strategy in macaques (rhesus macaques58, Japanese macaques59). Although this mating strategy is not as successful as consorting for siring offspring in male rhesus macaques58, it does introduce additional opportunities for sperm competition. Alpha male rhesus macaques are unable to fully monopolize females because of the high degree of estrous synchrony43,60, and they may benefit from being tolerant of other males in their group mating because it may lead to longer tenure length for those alpha males. For example, dominant male geladas (Theropithecus gelada) that were tolerant of other males in their groups had larger groups of females and longer tenure lengths compared to single-male units; thus, they had reproductive benefits even though they were sharing reproductive opportunities with other males each year61. Given the patterns of multiple matings by females, it is likely that sperm competition plays an important role in paternity in rhesus macaques. Furthermore, these patterns highlight the potential importance of female mate choice in rhesus macaques. Future research should investigate the role of females in biasing paternity against males over 15 years of age.
It is possible that the older males in our study may have had reduced sperm quantity or quality. Sperm depletion may be more noticeable in highly-ranked males who have more mating opportunities, and older males may no longer be physiologically capable of producing as much sperm as necessary for these high rates of mating behaviors41. For example, in a study of domestic fowl (Gallus gallus domesticus), older male ejaculate had fewer sperm than young males and those sperm had lower swimming velocity, which meant that young male ejaculate had a strong fertilizing advantage over older male ejaculate62. Our previous work suggests that high-ranking male rhesus macaques, especially alpha males, experience significant energetic stress during the breeding season, particularly at the beginning of the season when the maximum number of fertile females are available63. The toll of this stress and of investing effort into mate guarding and behavioral displays during mate guarding34 may have reproductive costs for older males. In men, age-related declines in sperm production and quality have been reported8,9,11,12,13, but there are contradictory reports (see10,12) and variation cross-culturally9. Future studies should explore variation in sperm quantity and quality over the lifetime. Given the difficulty of measuring ejaculate quality directly, examining changes in testes size may provide a proxy for understanding reproductive senescence in older male rhesus macaques, especially given that Bercovitch64 found that high-ranking males had the largest testes during the mating season.
In addition to decreased sperm quality or quantity, low reproductive output for older males may be the results of low infant survival (similar to the pattern shown in females17). Given that we could not account for the paternity of the 33 infants that died and the 7 stillbirths from the study year, we cannot determine how many of those infants were fathered by our study subjects. It is possible that older males have low infant survival or low sperm quality or both. Ultimately, these issues still result in reduced reproductive success for older males, indicating post-copulatory reproductive senescence.
Evidence of mechanisms of age-specific rates of reproduction are important for understanding life-history evolution, yet few studies exist on this topic. Most studies of reproductive skew focus on a male’s ability to monopolize females and female mate choice, but do not examine differences in male reproductive physiology over age in relation to skew. Future studies should incorporate measures of reproductive senescence in understanding variation in male reproductive success over their lifetime, including examining long-term cost of early reproductive effort (see65). The timing of reproduction in male rhesus macaques is influenced by variation in the serotonin transporter gene51, and future studies should investigate differences in reproductive senescence in relation to genotype. Importantly, we were not able to compare males of all ranks within this study. Future studies should also compare variation in reproductive success for males in other rank categories.
Our results provide important considerations for understanding male reproductive senescence across primates and potential evolutionary underpinnings of reproductive senescence in humans. Infertility is a worldwide clinical problem for human health and affects 8–12% of couples9. To add to this problem, there has been a significant increase in parental age in recent years12. As with studies of nonhuman animals, most studies of reproductive senescence in humans have focused on women. Studies of age-related changes to men’s sperm have provided contradictory evidence10,12. And cross-cultural variation in these changes make the issue difficult to understand through human studies9. Furthermore, most studies in reproductive senescence use a cross-sectional approach to examine variation in reproductive functioning. Here, we provide evidence of decline in reproductive activity within individuals. Future studies should continue to gather these long-term data on variation in male reproductive health and how social and physiological factors can impact a male’s ability to sire offspring.
References
Kappeler, P. M. & van Schaik, C. P. Sexual selection in primates: review and selective preview. In Sexual Selection in Primates (eds Kappeler, P. M. et al.) 3–23 (Cambridge University Press, Cambridge, 2004). https://doi.org/10.1017/CBO9780511542459.003.
Altmann, S. A. A field study of the sociobiology of rhesus monkeys, Macaca mulatta*. Ann. N. Y. Acad. Sci. 102, 338–435 (1962).
Alberts, S. C., Watts, H. E. & Altmann, J. Queuing and queue-jumping: long-term patterns of reproductive skew in male savannah baboons, Papio cynocephalus. Anim. Behav. 65, 821–840 (2003).
Setchell, J. M., Charpentier, M. & Wickings, E. J. Mate guarding and paternity in mandrills: factors influencing alpha male monopoly. Anim. Behav. 70, 1105–1120 (2005).
Altmann, J., Gesquiere, L., Galbany, J., Onyango, P. O. & Alberts, S. C. Life history context of reproductive aging in a wild primate model. Ann. N. Y. Acad. Sci. 1204, 127–138 (2010).
Setchell, J. M., Jean Wickings, E. & Knapp, L. A. Signal content of red facial coloration in female mandrills (Mandrillus sphinx). Proc. R. Soc. B Biol. Sci. 273, 2395–2400 (2006).
Bribiescas, R. G. On the evolution, life history, and proximate mechanisms of human male reproductive senescence. Evolut. Anthropol. Issues News Rev. 15, 132–141 (2006).
Eskenazi, B. et al. The association of age and semen quality in healthy men. Hum. Reprod. 18, 447–454 (2003).
Kumar, N. & Singh, A. K. Trends of male factor infertility, an important cause of infertility: a review of literature. J. Hum. Reprod. Sci. 8, 191–196 (2015).
Plas, E., Berger, P., Hermann, M. & Pflüger, H. Effects of aging on male fertility?. Exp. Gerontol. 35, 543–551 (2000).
Jung, A., Schuppe, H.-C. & Schill, W.-B. Comparison of semen quality in older and younger men attending an andrology clinic. Andrologia 34, 116–122 (2002).
Sharma, R. et al. Effects of increased paternal age on sperm quality, reproductive outcome and associated epigenetic risks to offspring. Reprod. Biol. Endocrinol. 13, 35 (2015).
Ng, K. K. Sperm output of older men. Hum. Reprod. 19, 1811–1815 (2004).
Kuhnert, B. Reproductive functions of the ageing male. Hum. Reprod. Update 10, 327–339 (2004).
Bribiescas, R. G. An evolutionary and life history perspective on human male reproductive senescence. Ann. N. Y. Acad. Sci. 1204, 54–64 (2010).
Dixson, A. F. Primate Sexuality: Comparative Studies of the Prosimians, Monkeys, Apes, and Humans (Oxford University Press, Oxford, 2012).
Hoffman, C. L., Higham, J. P., Mas-Rivera, A., Ayala, J. E. & Maestripieri, D. Terminal investment and senescence in rhesus macaques (Macaca mulatta) on Cayo Santiago. Behav. Ecol. 21, 972–978 (2010).
Johnson, R. L. & Kapsalis, E. Ageing, infecundity and reproductive senescence in free-ranging female rhesus monkeys. Reproduction 105, 271–278 (1995).
Johnson, R. L. & Kapsalis, E. Menopause in free-ranging rhesus macaques: estimated incidence, relation to body condition, and adaptive significance. Int. J. Primatol. 19, 751–765 (1998).
Cowlishaw, G. & Dunbar, R. I. M. Dominance rank and mating success in male primates. Anim. Behav. 41, 1045–1056 (1991).
Alberts, S. C., Buchan, J. C. & Altmann, J. Sexual selection in wild baboons: from mating opportunities to paternity success. Anim. Behav. 72, 1177–1196 (2006).
Black, A. & Lane, M. A. Nonhuman primate models of skeletal and reproductive aging. Gerontology 48, 72–80 (2002).
Setchell, J. M. Sexual Selection and the differences between the sexes in Mandrills (Mandrillus sphinx). Am. J. Phys. Anthropol. 159, 105–129 (2016).
Widdig, A. et al. A longitudinal analysis of reproductive skew in male rhesus macaques. Proc. R. Soc. Lond. B 271, 819–826 (2004).
Lemaître, J.-F. & Gaillard, J.-M. Reproductive senescence: new perspectives in the wild: Reproductive senescence in the wild. Biol. Rev. 92, 2182–2199 (2017).
Maestripieri, D. Macachiavellian Intelligence: How Rhesus Macaques and Humans Have Conquered the World (University of Chicago Press, Chicago, 2007).
Manson, J. H. Evolved psychology in a novel environment. Hum. Nat. 9, 97–117 (1998).
Georgiev, A. V. et al. Breaking the succession rule: the costs and benefits of an alpha-status take-over by an immigrant rhesus macaque on Cayo Santiago. Behaviour 153, 325–351 (2016).
Noordwijk, M. A. V. & Schaik, C. P. V. Career moves: transfer and rank challenge decisions by male long-tailed macaques. Behaviour 138, 359–395 (2001).
Sapolsky, R. M. Endocrine aspects of social instability in the olive baboon (Papio anubis). Am. J. Primatol. 5, 365–379 (1983).
Sapolsky, R. M. Endocrinology Alfresco: psychoendocrine studies of wild baboons. In Recent Progress in Hormone Research (ed. Bardin, C. W.) 437–468 (Academic Press, New York, 1993). https://doi.org/10.1016/B978-0-12-571148-7.50020-8.
Sapolsky, R. M. Cortisol concentrations and the social significance of rank instability among wild baboons. Psychoneuroendocrinology 17, 701–709 (1992).
Higham, J. P., Heistermann, M. & Maestripieri, D. The endocrinology of male rhesus macaque social and reproductive status: a test of the challenge and social stress hypotheses. Behav. Ecol. Sociobiol. 67, 19–30 (2013).
Milich, K. M. & Maestripieri, D. Sex or power? The function of male displays in rhesus macaques. Behaviour 153, 245–261 (2016).
Rawlins, R. G. & Kessler, M. J. Climate and seasonal reproduction in the Cayo Santiago macaques. Am. J. Primatol. 9, 87–99 (1985).
Rawlins, R. G. & Kessler, M. J. The Cayo Santiago Macaques: History, Behavior, and Biology (SUNY Press, New York, 1986).
Maestripieri, D. & Wallen, K. Affiliative and submissive communication in rhesus macaques. Primates 38, 127–138 (1997).
Bercovitch, F. B. et al. A longitudinal study of age-specific reproductive output and body condition among male rhesus macaques, Macaca mulatta. Naturwissenschaften 90, 309–312 (2003).
Kessler, M. J. et al. Long-term effects of tetanus toxoid inoculation on the demography and life expectancy of the Cayo Santiago rhesus macaques. Am. J. Primatol. 77, 211–221 (2015).
Boesch, C., Kohou, G., Néné, H. & Vigilant, L. Male competition and paternity in wild chimpanzees of the Taï forest. Am. J. Phys. Anthropol. 130, 103–115 (2006).
Wroblewski, E. E. et al. Male dominance rank and reproductive success in chimpanzees, Pan troglodytes schweinfurthii. Anim. Behav. 77, 873–885 (2009).
Bercovitch, F. B. & Nürnberg, P. Socioendocrine and morphological correlates of paternity in rhesus macaques (Macaca mulatta). Reproduction 107, 59–68 (1996).
Dubuc, C., Muniz, L., Heistermann, M., Engelhardt, A. & Widdig, A. Testing the priority-of-access model in a seasonally breeding primate species. Behav. Ecol. Sociobiol. 65, 1615–1627 (2011).
Chapais, B. Reproductive activity in relation to male dominance and the likelihood of ovulation in rhesus monkeys. Behav. Ecol. Sociobiol. 12, 215–228 (1983).
Charlesworth, B., Harvey, P. H., Packer, C., Collins, D. A. & Eberly, L. E. Problems with primate sex ratios. Phil. Trans. R. Soc. Lond. Ser. B Biol. Sci. 355, 1627–1635 (2000).
Torres, R. & Velando, A. Male reproductive senescence: the price of immune-induced oxidative damage on sexual attractiveness in the blue-footed booby. J. Anim. Ecol. 76, 1161–1168 (2007).
Richard, M., Lecomte, J., Fraipont, M. D. & Clobert, J. Age-specific mating strategies and reproductive senescence. Mol. Ecol. 14, 3147–3155 (2005).
Georgiev, A. V., Thompson, M. E., Mandalaywala, T. M. & Maestripieri, D. Oxidative stress as an indicator of the costs of reproduction among free-ranging rhesus macaques. J. Exp. Biol. 218, 1981–1985 (2015).
Inoue, E. & Takenaka, O. The effect of male tenure and female mate choice on paternity in free-ranging Japanese macaques. Am. J. Primatol. 70, 62–68 (2008).
Kuester, J., Paul, A. & Arnemann, J. Age-related and individual differences of reproductive success in male and female barbary macaques (Macaca sylvanus). Primates 36, 461–476 (1995).
Krawczak, M. et al. Male reproductive timing in rhesus macaques is influenced by the 5HTTLPR Promoter polymorphism of the serotonin transporter gene. Biol. Reprod. 72, 1109–1113 (2005).
Reed, T. E. et al. Reproductive senescence in a long-lived seabird: rates of decline in late-life performance are associated with varying costs of early reproduction. Am. Nat. 171, E89–E101 (2008).
Parker, G. A. Sperm competition and its evolutionary consequences in the insects. Biol. Rev. 45, 525–567 (1970).
Harcourt, A. H., Harvey, P. H., Larson, S. G. & Short, R. V. Testis weight, body weight and breeding system in primates. Nature 293, 55–57 (1981).
Bercovitch, F. B. & Rodriguez, J. F. Testis size, epididymis weight, and sperm competition in rhesus macaques. Am. J. Primatol. 30, 163–168 (1993).
Carpenter, C. R. Sexual behavior of free ranging rhesus monkeys (Macaca mulatta). I. Specimens, procedures and behavioral characteristics of estrus. J. Compar. Psychol. 33, 113–142 (1942).
Loy, J. Estrous behavior of free-ranging rhesus monkeys (Macaca mulatta). Primates 12, 1–31 (1971).
Berard, J. D., Nurnberg, P., Epplen, J. T. & Schmidtke, J. Alternative reproductive tactics and reproductive success in male rhesus macaques. Behaviour 129, 177–201 (1994).
Soltis, J., Thomsen, R. & Takenaka, O. The interaction of male and female reproductive strategies and paternity in wild Japanese macaques, Macaca fuscata. Anim. Behav. 62, 485–494 (2001).
Vandenbergh, J. G. & Vessey, S. Seasonal breeding of free-ranging rhesus monkeys and related ecological factors. Reproduction 15, 71–79 (1968).
Snyder-Mackler, N., Alberts, S. C. & Bergman, T. J. Concessions of an alpha male? Cooperative defence and shared reproduction in multi-male primate groups. Proc. R. Soc. B 279, 3788–3795 (2012).
Dean, R. et al. Male reproductive senescence causes potential for sexual conflict over mating. Curr. Biol. 20, 1192–1196 (2010).
Milich, K. M., Georgiev, A. V., Petersen, R. M., Emery Thompson, M. & Maestripieri, D. Alpha male status and availability of conceptive females are associated with high glucocorticoid concentrations in high-ranking male rhesus macaques (Macaca mulatta) during the mating season. Horm. Behav. 97, 5–13 (2018).
Bercovitch, F. B. Dominance rank and reproductive maturation in male rhesus macaques (Macaca mulatta). Reproduction 99, 113–120 (1993).
Lemaître, J.-F., Gaillard, J.-M., Pemberton, J. M., Clutton-Brock, T. H. & Nussey, D. H. Early life expenditure in sexual competition is associated with increased reproductive senescence in male red deer. Proc. R. Soc. B Biol. Sci. 281, 20140792 (2014).
Acknowledgements
We thank the staff of the Caribbean Primate Research Center (CPRC) for support, especially Nahiri Rivera, Edgar Davila, Giselle Caraballo, and Julio Resto. Field assistance was provided by Diana Christie, Clement Ludcher, Jesus Madrid, and Amaury Michel. Thank you to Anja Widdig for contributing to the CPRC genetic database. Thanks, also, to Hongjie Gu and the Washinton University in St. Louis Division of Biostatistics. Thank you to Tommaso Pizzari, Josh Firth, and one anonymous reviewer for helpful comments. Genetic and demographics data of the CPRC database are funded by Grant Number 8 P40 OD012217 from the National Center for Research Resources (NCRR) and the Office of Research Infrastructure Programs (ORIP) of the National Institutes of Health. The content of this publication is solely the responsibility of the authors and does not necessarily represent the official views of NCRR, ORIP, or the NIH. The investigation was approved by the IACUC of the University of Puerto Rico, Medical Sciences Campus (Protocol No. A0100108).
Author information
Authors and Affiliations
Contributions
K.M.M. designed the study, collected the behavioral observations, participated in data analysis, and drafted the manuscript. A.R.L. conducted molecular lab work, oversaw the maintenance of the long-term database and extraction of data from it, and reviewed the manuscript draft. E.M. maintained the long-term database and extracted relevant paternity data from it for analysis. D.M. conceived of and coordinated the study and critically revised the manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Milich, K.M., Ruiz-Lambides, A., Maldonado, E. et al. Age negatively impacts reproduction in high-ranking male rhesus macaques on Cayo Santiago, Puerto Rico. Sci Rep 10, 13044 (2020). https://doi.org/10.1038/s41598-020-69922-y
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-020-69922-y
This article is cited by
-
Facial and genital color ornamentation, testosterone, and reproductive output in high-ranking male rhesus macaques
Scientific Reports (2024)
-
Should I stay or should I go now: dispersal decisions and reproductive success in male white-faced capuchins (Cebus imitator)
Behavioral Ecology and Sociobiology (2022)
-
Twenty-three-year demographic history of the Affenberg Japanese macaques (Macaca fuscata), a translocated semi-free-ranging group in southern Austria
Primates (2021)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
