
Abstract
Extreme climate events and nitrogen (N) deposition are increasingly affecting the structure and function of terrestrial ecosystems. However, the response of plant biomass to variations to these global change drivers is still unclear in semi-arid regions, especially in degraded sandy grasslands. In this study, a manipulative field experiment run over two years (from 2017 to 2018) was conducted to examine the effect of rainfall alteration and nitrogen addition on biomass allocation of annuals and perennial plants in Horqin sandy grassland, Northern China. Our experiment simulated extreme rainfall and extreme drought (a 60% reduction or increment in the growing season rainfall with respect to a control background) and N addition (20 g/m2) during the growing seasons. We found that the sufficient rainfall during late July and August compensates for biomass losses caused by insufficient water in May and June. When rainfall distribution is relatively uniform during the growing season, extreme rainfall increased aboveground biomass (AGB) and belowground biomass (BGB) of annuals, while extreme drought reduced AGB and BGB of perennials. Rainfall alteration had no significant impacts on the root-shoot ratio (R/S) of sandy grassland plants, while N addition reduced R/S of grassland species when there was sufficient rainfall in the early growing season. The biomass of annuals was more sensitive to rainfall alteration and nitrogen addition than the biomass of perennials. Our findings emphasize the importance of monthly rainfall distribution patterns during the growing season, which not only directly affect the growth and development of grassland plants, but also affect the nitrogen availability of grassland plants.
Similar content being viewed by others

Introduction
Climate change and excessive human activity changed the rainfall patterns and increased the emissions of bioactive nitrogen (N) into the atmosphere, which have produced profound impacts on the global rainfall and N cycle1,2. Rainfall and N are key environmental factor determining ecosystem structure and function, especially in water and N limited grasslands3. Therefore, changes in rainfall and N may have a strong impact on terrestrial ecosystem and may feed back into climate change.
Plant biomass allocation refers to the distribution of limited resources by plants in order to maximize the benefits of growth, maintenance and reproduction in response to environmental clues having profound implications for plant growth and development4. Increasing rainfall can generally promote the accumulation of aboveground biomass while decreasing rainfall can promote the growth of root5. However, the accumulation of aboveground and belowground biomass is not synchronous6. Plants also adapt to drought by increasing the root-shoot ratio (R/S)7. However, some studies have also found a negative correlation between rainfall and biomass, as rainfall increases soil erosion and decreases soil organic matter content, thus reducing grassland productivity8. In summary, the effect of rainfall variation on biomass allocation varies with rainfall gradients, elevation gradient and species composition9.
Changes in global water circulation are forecast to enhance both inter- and intra-annual variability of rainfall10,11. Chinese scientists forecast that rainfall patterns will be more complicated and multifrequency in the next 30 years in north of China12, and this will produce more frequent rainfall and drought events in the future13. Studies of plant biomass have focused mainly on the effect of total annual rainfall14, but recent research showed that rainfall at a particular time of the year (seasonal rainfall) can explain biomass changes better than total rainfall16,17 due to the different water requirements of terrestrial plants at different growth stages15,18. Studies have showed that advanced rainfall during the growing period can lead to an improvement in the utilization of soil water, which is beneficial to root growth19. By contrast, delayed rainfall often increases the availability of soil water at the later stage of growth, thus delaying the plant senescence20. However, it is less known how excessive rainfall and extreme drought affect biomass allocation among years with different rainfall distribution during the growing season.
Nitrogen inputs to the earth’s ecosystems are increasing, and this will have profound impacts on the function of terrestrial ecosystems21. For instance, during the period 1990–2003, atmospheric N deposition increased dramatically from 8.7 kg N ha −1 to 13.8 kg N ha −1 22. Many studies showed that increased amounts of available N decreased root biomass and R/S23. The effect of N on grassland species is determined by rainfall conditions, and increasing soil N availability increased water absorption capacity of plants23,24,25. However, little is known with respect to how rainfall changes, N deposition and their interaction in semiarid grasslands will affect the allocation of aboveground and belowground biomass distribution of annuals and perennials.
In semi-arid regions, the vegetation biomass is most sensitive to water availability because rainfall is concentrated in the plant’s growing period26. Therefore, rainfall events with long duration and copious amounts during the growing season may play a primary role in regulating the growth of vegetation13,27. Semiarid ecosystems such as the Horqin sandy grasslands in Inner Mongolia may be particularly susceptible to N deposition and climate change as the region is largely constrained by both resources28. Annuals and perennials are the main herbaceous plants in the Horqin sandy grassland. Annuals have a fast-growing strategy (e.g., shallower/fine roots), which allows plants to rapidly absorb water and nutrients under favorable conditions to complete their life cycle29. Compared to annuals, perennials have a stronger ability of survival and competition in nutrient-poor and water-deficient environments because of their deeper intricate root system30. So annuals are more likely to survive in harsh environment than perennials, and exploring the biomass allocation strategies of annuals and perennials can be a powerful predictor for environmental change. In the context of global climate change, conducting research through coupled rainfall-nitrogen experiments is of great significance for a deeper understanding of adaptation mechanisms of degraded grassland species to climate change drivers. In this study, we investigated aboveground biomass (AGB), belowground biomass (BGB), coarse roots biomass (CR) and fine roots biomass (FR) of annuals and perennials in a grassland community under different rainfall manipulation and nitrogen treatments. We addressed the following questions: (1) how do AGB, BGB, CR and FR of annuals and perennials change with rainfall alterations and nitrogen addition? (2) How do the allocation of AGB and BGB between annuals and perennials differ in their response to changes in rainfall and N?
Materials and Methods
Study area
This study was carried out in Horqin sandy grassland near the Naiman Desertification Research Station, Chinese Academy of Science. The climate is typical semiarid continental seasonal monsoon. The average annual rainfall is 360 mm, and nearly 75% is concentrated in the growing season (May to August)31. The soil is aeolian sandy soil according to the Chinese soil taxonomy classification system (http://www.resdc.cn).
Experimental design and measuring
The experiment was conducted between 2017 and 2018 on sandy grassland which was relatively homogeneous and not so severely degraded. Based on the long-term observation data of total annual rainfall, extreme rainfall and extreme drought events during the growing season in this region32, we set up the extreme rainfall and extreme drought treatments as follows: a 60% increment (with respect to background rainfall) of rainfall during the growing season from May 1 to August 31 (extreme rainfall), and a 60% reduction of rainfall during the growing season from May 1 to August 31 (extreme drought).
In order to simulate the effect of N deposition, we adopted a rather high N addition level (20 g nitrogen / m22) in our experiment. There are three reasons we used this level of N addition: First, the sandy grassland is relatively barren33, thus in order to achieve a better experimental effect, the amount of added N needs to be high. Second, the Horqin sandy land is located in the ecologically fragile zone of semi-arid farming-pastoral interlacing area in Northern China. Large-scale human activities (farmland fertilization and animal husbandry) in this area have intensified the N input, resulting in excessive N load in the area34. Besides, by referring to N deposition levels in some countries around the world (e.g., USA and Europe)35, a 20 g N / m22 rate is deemed representative of the highest deposition levels in larger areas of China36. Thus in this experiment, 10 g N / m2 were added sequentially both in May and July in 2017 and 2018. Six treatments (with 6 replicates each) were randomly arranged for the interaction of rainfall change and N addition. The treatment included control (CK), + 60%, −60%, +N, + 60% × +N, −60% × +N (Fig. 1d). The rainfall alteration device (rainfall shelters) was firstly described by Yahdjian and Sala28, and has been widely used in research of climate change, owing to its low cost and minimal influence on the microclimate37,38. Each rainfall shelter was made of clear polycarbonate plastic strips that allow 90% sunshine penetration to ensure no great alterations on plant photosynthesis (Fig. 1c).

Location of the study area (a), aerial photograph of the field site (b), rainfall shelters installation (c) and experiment design (d). An artificial simulation of rainfall device in the field experiments to achieve the goal of increasing or decreasing rainfall: (c) ±60%: increasing or decreasing rainfall by 60% during the growing season (May to August).
In mid-August of 2017 and 2018, we selected a representative land with evenly distributed vegetation; containing as many species as possible. For each plot one soil cube of 30 × 30 × 30 cm was excavated with whole species in each of the plots. The samples were gently shaken to get rid of the soil particles attached to roots, and then brought to the laboratory for washing off the remained soil particles. The AGB and BGB of each of plant species were measured. Moreover, BGB was separated into CR (diameter greater than 2 mm) and FR (diameter less than 2 mm) by the vernier caliper39. The separated biomass was oven-dried at 85 °C for 48 h to obtain dry weight.
Data analysis
The slope of the rainfall-biomass relationship reflects the sensitivity of biomass to rainfall variability. A sensitivity of 1 indicates that a relative change in response to parameters in the same direction28. Sensitivity was calculated by the relative change in the rainfall manipulation or N addition plots in comparison to the control plots as:
Where \(biomas{s}_{control}\) and \(biomas{s}_{\overline{ck}}\) represents the biomass of plants under either a control plot or a treatment plot respectively. The higher the value of sensitivity the higher the sensitivity of biomass to changes in variation in rainfall.
Three-way ANOVA was used to test the effects of rainfall pattern, nitrogen addition impact on the AGB, BGB, CR and FR in the two different years. Results were considered significantly different at a P < 0.05 level. Data analysis and plotting were carried out by SPSS 21.0 and SigmaPlot 12.5, respectively.
Results
Rainfall manipulation and nitrogen addition in different years had significant effects on AGB, BGB, CR and FR of annuals and perennials in sandy grassland, but the interactive effects of rainfall and nitrogen had no significant effect on biomass (Table 1).
Rainfall change
Overall, the total rainfall during the growing season did not differed much between 2017 and 2018, but the monthly distribution of rainfall was quite different between the two years. In this way the rainfall amount in the early period of the growing season (May to June) in 2017 was 41.8 mm; only half of that in the early time of the growing season in 2018 (100.8 mm). The rainfall in the late period of the growing season (July to August) in 2017 was 232.6 mm, while in the late period of the growing season in 2018 was just 136.6 mm (Table 2).
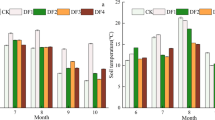
At the grassland community level, extreme drought (-60%) significantly reduced AGB by 44% in 2017, while extreme rainfall (+60%) significantly enhanced it by 115% in 2018 (Fig. 2a). On the other hand, extreme rainfall and extreme drought had no significant effects on the BGB and CR in 2017 and 2018 (Fig. 2b,c). Extreme rainfall and extreme drought had no significant effects on FR in 2017, while extreme drought significantly reduced FR in 2018 by 62% (Fig. 2d). Extreme rainfall and extreme drought had no significant effects on AGB, BGB, CR and FR in 2017 for neither annuals nor perennials, (Fig. 2e–l). On the other hand, extreme rainfall significantly enhanced AGB, BGB and FR of annuals by 579%, 202% and 800% respectively in 2018 (Fig. 2e,f,h), and extreme drought significantly reduced the AGB and BGB and FR of perennials by 53%, 63% and 63% respectively in 2018 (Fig. 2i,j,l). Extreme rainfall and extreme drought had no significant effects on CR of annuals and perennials in both of 2017 and 2018 (Fig. 2g,k).

Influence of rainfall alteration on the biomass of grassland plants. Different values represent mean ± SE. Different letters indicate significant differences between rainfall treatments in the same year (P < 0.05). Significant differences between 2017 and 2018 are indicated by asterisks, *P < 0.05, **P < 0.01.
Nitrogen addition
For the whole grassland community, N addition had no significant effects on AGB, BGB, CR and FR in 2017 (Fig. 3a–d), but it decreased BGB by 32% in 2018 (Fig. 3b). In 2017, N addition significantly decreased AGB and CR of annuals by 50% and 85% respectively (Fig. 3e,j), while it enhanced AGB by 87% of perennials (Fig. 3i). In 2018, N addition had no significant effect on AGB, BGB, CR and FR of neither annuals nor perennials (Fig. 3e–l).

The influence of nitrogen addition on biomass of grassland plants. Different letters indicate significant differences between nitrogen treatments in the same year (P < 0.05). Significant differences between 2017 and 2018 are indicated by asterisks, *P < 0.05, **P < 0.01.
R/S
The R/S of community, annuals and perennials had different responses to rainfall alteration and N addition (Fig. 4). Rainfall change had no significant effects on the R/S of the community, annuals or perennials (Fig. 4a–c). Under extreme drought, R/S of the plant community, annuals and perennials varied significantly between the two years. Besides, the R/S of annuals was significantly different in two years with different rainfall patterns. In this way, N addition significantly decreased the R/S by 56% of perennials in 2017(Fig. 4f), and it significantly decreased the R/S of community, annuals and perennials by 64%, 50% and 51% respectively in 2018(Fig. 4d–f).

The effect of rainfall change and nitrogen addition on the root-shoot ratio (R/S). Different letters indicate significant differences between nitrogen treatments in the same year (P < 0.05). Significant differences between 2017 and 2018 are indicated by asterisks, *P < 0.05, **P < 0.01.
Sensitivity
Sensitivities of AGB, BGB and FR in response to rainfall alteration varied in the annuals and perennials (Fig. 5,6). Specifically, AGB, BGB and FR of the annuals were more sensitive than those of perennials to rainfall changes (Fig. 5a,b,d). Similarly, AGB, BGB and FR of the annuals were more sensitive than those of perennials to N addition (Fig. 6a,b,d). However, CR of the annuals and perennials was not sensitive to rainfall change and N addition (Fig. 5c,6c). In summary, the biomass of annuals was more sensitive than the biomass of perennials to rainfall change and N addition.

Sensitivities of AGB, BGB, CR and FR of grassland plants to rainfall change. Different letters represent the difference in nitrogen sensitivity between annuals and perennials (P < 0.05).

Sensitivities of AGB, BGB, CR and FR of grassland plants to nitrogen addition. Different letters represent the difference in nitrogen sensitivity between annuals and perennials (P < 0.05).
Discussion
Rainfall
In the present study, extreme drought decreased AGB of the whole grassland community in 2017, while extreme rainfall enhanced AGB of grassland community in 2018 (Fig. 2a–d). These results demonstrate a clear pattern that increased rainfall significantly increases the biomass of grassland species, while decreased rainfall significantly decreases the biomass of grassland species. The result is consistent with long-term observations of terrestrial ecosystems around the world40. In addition, our results show that although total rainfall during the growing season of 2017 and 2018 was almost equal, the effects of extreme rainfall and extreme drought on AGB, BGB, CR and FR were greatly different between the two years (Table 1; Fig. 2). This result further confirms previous research which has showed that the seasonal distribution and intensity of rainfall, rather than total rainfall determine grassland productivity16,18.
Plants in arid environments show a delayed phenology to reduce water loss41. This response mechanism occurs under drought conditions, indicating that plants have begun to enter into dormancy, which continues into the next rainfall season, thus forming a strategy to improve survival. In this way, longer intervals between rainfall events shorten the growing season fort plants42. In addition, the recovery of plants after the dormant period is closely related to the interval and intensity of rainfall after drought43. Our results further support this view: rainfall change had impacts on the biomass of both annuals and perennials in 2018, but not in 2017 (Fig. 2e–f). This differential response of biomass was mainly caused by rainfall differences in the early time of growing season (May to June) between the two years. Thus, in 2017, prolonged drought in May and June delayed the germination of grass seeds. Until July, extreme drought did not affect the normal growth and development of grassland plants, because sufficient rainfall in July and August exceeded the rain threshold needed for plant growth, so the 60% decreased rainfall treatment could also meet the requirements for normal growth of sandy grassland plants14,18. These results showed that in late July and August, rainfall compensates for biomass losses caused by insufficient water in early stages of the growing season. Thus, a period of sufficient rainfall during the growing season may play a more important role in promoting biomass accumulation. These findings are inconsistent with studies in other places where rainfall in later periods in the growing season has been associated with a higher risk of microclimate moisture, bacterial invasion, and a higher risk of soil compaction44. The main reason for discrepancies in our results and those of previous studies is the difference of environmental factors (temperature, moisture, elevation, etc.) in different regions. Sandy soil has a large number of large particles, higher soil saturated hydraulic conductivity and evaporation, so the soil water content decreases rapidly after rainfall events due to its low water-holding capacity45. Therefore, rainfall later in the growing season will not harm the ecosystem structure of sandy grassland.
The rainfall was distributed more evenly during the growing season in 2018. Therefore, extreme rainfall or extreme drought would be expected to have positive or negative impacts on vegetation biomass accordingly. Extreme rainfall significantly increased the AGB and BGB of the annuals, and extreme drought significantly reduced the AGB and BGB of perennials (Fig. 2e–l). This was mainly due to the fact that the shallower roots of annuals can use water quickly and complete their life cycle rapidly under favorable water conditions45. Therefore, increasing rainfall by 60% throughout the growing season can effectively increase the AGB and BGB of annuals. Although perennials have a well developed and deep root system, because of the coarse texture of the soil and its lower water-holding capacity, extreme drought dramatically reduced the effective moisture of sandy soil45. This, the roots of perennials could not obtain enough water from deep soil layers, and the AGB and BGB of perennials were synchronously decreased14.
Nitrogen
As a limiting nutrient in semi-arid regions, N has a great influence on plant growth46. Our results showed that in 2017, N addition decreased the AGB of annuals, while increased the AGB of perennials (Fig. 3e, i). This is mainly because the water deficit in the early growing season in 2017 made N the main limiting element for plant growth. While added N eliminated nutrient limitation and turned plants’ competition for nutrients into competition for other resource such as light or water32,47. Taller perennials had a competitive advantage over light resource. In addition, the developed root system of perennials also provided them with a competitive advantage for water resources. As perennial individuals grew larger, they would devote more biomass to photosynthetic organs (e.g. stem and leaf) to enhance productivity48. On the contrary, annuals lose their competitiveness for light and water resources and their AGB decreased accordingly49. This is consistent with previous studies where a reduction in biomass of some plant species was compensated by an increase in biomass of other plant species in the plant community29,30.
R/S
Isometric allocation hypothesis demonstrated that AGB scales one-to-one with respect to BGB among different kinds of plants and this relationship is insensitive to changes in environmental conditions50,51. Other researchers have proposed optimal partitioning theory, which suggests that plants preferentially allocate biomass to the organ that is more efficient in obtaining resources4,50. In this study, rainfall changes had no significant effects on the R/S of community, annuals and perennials in 2017 and 2018 (Fig. 4a–c). These results are inconsistent with previous research showing that plants often allocated more biomass resources to aboveground reproductive organs to further improve reproduction and photosynthetic capacity when rainfall is abundant51, or plants often allocate more biomass to belowground in response to extreme drought52. One possible explanation may be that the allocation of plant biomass depends largely on the size of the plant itself, rather than the external environment53. Another explanation is that R/S of sandy soil species may take longer time to respond to rainfall change. As previous studies have shown rainfall changes do affect the allocation of plant biomass, but these responses become apparent only 10 years after the experimental manipulation54. Although rainfall change had no significant effect on R/S, R/S did vary significantly between the two years, especially in the extreme drought condition. This suggests that under extreme drought conditions, AGB and BGB decreased synchronously. These conclusions verify the role of compensatory interactions among sandy grassland plants, and suggest that competition of plants in the sandy grassland community would result in a trade-off between annuals and perennials55.
In addition, we found that N addition had no effect on R/S of the whole grassland community in 2017, while it decreased R/S of the whole grassland community in 2018 (Fig. 4d–f). The main reason for this result is the difference in rainfall distribution. In 2018, N stimulated by abundant water caused soil acidification56, which causes the roots of plants to be exposed to a high concentration and toxicity of protons (H+), aluminum (Al3+) as well as alteration of resource stoichiometry57. These effects directly lead to a decrease in the respiration rate of soil microorganisms, partially offsetting root respiration and thus reducing BGB56. This is consistent with the results of previous studies, where the responses of different microbial to N addition are likely due to different soil water content58. These results were partly consistent with other grassland manipulative experiments, which suggests that in Horqin sandy grasslands, the coupling effects of rainfall and N early in the growing season may have a negative effect on the growth of plant roots. This further explains why nitrogen-water coupling has no significant effect on the growth of sandy grassland community, annuals and perennials (Table 1), indicating that decoupling of root morphology and their water uptake with increasing soil N availability.
Sensitivity
Our study showed that the R/S of annuals was significantly different in two years with different rainfall patterns (Fig. 4b), and AGB, BGB and FR of annuals were more sensitive to rainfall change and nitrogen addition than that of perennials (Fig. 5–6), which was consistent with the findings that annuals can adjust nutrient allocation much more faster than the perennials to complete their life cycle across favorable water and fertilizer conditions47,51,53. Similarly, we also have found that fine root was more sensitive to rainfall change and nitrogen addition than coarse roots. This can be explained by that coarse roots have little effect on absorption function59, while fine root is closely related to soil water and nutrients and is the main organ of water and nutrient absorption60. From the above, we are convinced that the sensitivity of annuals is mainly caused by their large number of fine roots.
In two-year study, we found that extreme rainfall and extreme drought have produced multiple effects on sandy grassland plants. It will be necessary to repeat our study in a year with more uniform rainfall and with a longer duration to see whether and how the results change. This study will help us to predict the impacts of climate change and make feasible decision for sustainable restoration and management of the degraded sandy grassland.
Conclusions
This study explored the biomass allocation of sandy grassland in semiarid regions and its relationships with environmental factors. We found that the sufficient rainfall in late growth season would promote or compensates for biomass losses caused by insufficient water in early growth season. In the years with more uniform rainfall during the growing season, extreme rainfall increased the biomass of annuals, and extreme drought decreased the biomass of perennials. R/S of sandy grassland plants was not affected by rainfall change, but coupling effects of rainfall and nitrogen in early growing season can reduce R/S of grassland plants. Along with total rainfall during the growing season is increasingly used to explain ecosystem processes, we argue that the distribution pattern of rainfall will have more profound impacts on the distribution of plant biomass in semi-arid grassland. The result of this research is of great importance for deserts plants adapt to global changes.
Change history
08 September 2020
An amendment to this paper has been published and can be accessed via a link at the top of the paper.
References
Basto, S. et al. Severe effects of long‐term drought on calcareous grassland seed banks. Npj Climate and At-mospheric. Science 1, 1 (2018).
Stevens, C. J. Nitrogen in the environment. Science 363, 578–580 (2019).
Epstein, H. E., Burke, I. C. & Lauenroth, W. K. Regional patterns of decomposition and primary production rates in the U.S. great plains. Ecology 83, 320–327 (2002).
Poorter, H., Reich, P. B., Oleksyn, J., Poot, P. & Mommer, L. Biomass allocation to leaves, stems and roots: meta-analyses of interspecific variation and environmental control. New Phytol-ogist 193, 30–50 (2012).
Fay, P. A., Carlisle, J. D., Knapp, A. K., Blair, J. M. & Collins, S. L. Productivity responses to altered rainfall patterns in a C4-dominated grassland. Oecologia 137, 245–251 (2003).
Wilcox, K. R. et al. Con-trasting above- and be-lowground sensitivity of three Great Plains grasslands to altered rainfall regimes. Global Change Biology 21, 335–344 (2014).
Huang, X., Liu, Y. & Li, J. X. The response of mulberry trees after seedling hardening to summer drought in the hydro-fluctuation belt of Three Gorges Reservoir Areas. Environmental Science and Pollution Re-search 20, 7103–7111 (2013).
Gao, Y. Z., Giese, M., Brueck, H., Yang, H. J. & Li, Z. J. The relation of biomass production with leaf traits varied under different land-use and precipitation condi- tions in an Inner Mongo-lia steppe. Ecological Research. 28, 1029–1043 (2013).
Qin, X. J., Sun, J. & Wang, X. D. Plant coverage is more sensitive than species diversity in indicating the dy-namics of the above-ground biomass along a precipitation gradient on the Tibetan Plateau. Ecological Indicators 84, 507–514 (2018).
IPCC Climate change 2014: Impacts, adaptation, and vulnerability. Part B: Re-gional aspects. In V. R. Barros, C. B. Field, D. J. Dokken, M. D. Mastrandrea, K. J. Mach, T. E. Bilir, L. L. White (Eds.), Contribution of working group II to the fifth assessment report of the intergovernmental panel on climate change (pp. 64–65). Cambridge, UK:Cambridge University Press (2014).
Sala, O. E., Gherardi, L. A. & Peters, D. P. C. Enhanced precipitation variability effects on water losses and ecosystem functioning: Differential response of arid and mesic regions. Climatic Change 131, 213–227 (2015).
Liu, Y. X. et al. Rional temperature and precipitation in the past 50 years and the next 30 years over China. Quaternary International 212, 57–63 (2010).
Easterling, D. R. et al. Climate extremes: ob-servations, modeling, and impacts. Science 289, 2068–2074 (2000).
Ma, W. H., Yang, Y. H., He, J. S., Hui, Z. & Fang, J. Y. Above- and belowground biomass in relation to envi-ronmental factors in temperate grasslands, Inner Mongolia. Science in China 51, 263–270 (2008).
Pierre, K. J. L. & Smith, M. D. Explaining temporal variation in above-ground productivity in a mesic grassland: the role of climate and flowering. Journal of Ecology 99, 1250–1262 (2011).
Muldavin, E. H., Moore, D. I., Collins, S. L., Wetherill, K. R. & Lightfoot, D. C. Above-ground net primary production dynamics in a northern Chihuahuan Desert ecosystem. Oecologia 155, 123–132 (2008).
Xia, Y., Moore, D. I., Collins, S. L. & Muldavin, E. H. Aboveground production and species richness of annu-als in Chihuahuan Desert grassland and shrubland plant communities. Journal of Arid Environments 74, 378–385 (2010).
Robinson, T. M. P. & Colby, S. E. Seasonal, not annual precipitation drives community productivity across ecosystems. Oikos 122, 727–738 (2013).
Wan, C., Yilmaz, I. & Sosebee, R. E. Seasonal soil-water availability influences snakeweed root dynamics. Journal of Arid Environments 51, 255–264 (2002).
Liu, Q. et al. Temperature, precipitation, and insolation effects on autumn vegetation phenology in temperate China. Global Change Biology 22, 644–655 (2016).
Tilman, D. et al. Forecasting agriculturally driven global environmental change. Sci-ence 292, 281–284 (2001).
Lü, C. & Tian, H. Spatial and temporal patterns of nitrogen deposition in China: Synthesis of observational data. Journal of Geophysical Research, 112(D22S05) (2007).
Shipley, B. & Meziane, D. The balanced-growth hypothesis and the allometry of leaf and root biomass allo-cation. Funct Ecol 16, 326–331 (2002).
Li, C. B. et al. Precipita-tion and nitrogen addition enhance biomass allocation to aboveground in an alpine steppe. Ecol-ogy and Evolution 9, 12193–12201 (2019).
Huxman, T. E. et al. Precipitation pulses and carbon fluxes in semiarid and arid ecosystems. Oecologia 141, 254–268 (2004).
Swemmer, A. M., Knapp, A. K. & Snyman, H. A. Intra-seasonal Precipitation Patterns and above-ground Productivity in Three Perennial Grasslands. Journal of Ecology 95, 780–788 (2007).
Meehl, G. A., Arblaster, J. M. & Tebaldi, C. Understanding Future Patterns of Increased Precipitation Intensity in Climate Model Simulations. Geophysical Research Letters 32, 109–127 (2005).
Hsu, J. S., Powell, J. & Adler, P. B. Sensitivity of mean annual primary production to precipitation. Global Change Biology 18, 2246–2255 (2012).
Venable, D. L. Bet hedging in a guild of desert annuals. Ecology 88, 1086–1090 (2007).
Kiær, L. P. & Weiner, J. Root and shoot competition: a meta-analysis. Journal of Ecology 101, 1298–1312 (2013).
Zuo, X. A. et al. cale de-pendent effects of environmental factors on vegetation pattern and composition in Horqin Sandy Land, Northern China. Geoderma 173, 1–9 (2012).
Huang, J. H. et al. Case study 3: Xilingol grassland, Inner Mongolia. Rangeland Degradation and Recovery in Chinas Pastoral Lands 8, 273–279 (2009).
Zuo, X. A. et al. Changes in carbon and nitrogen storage along a restoration gradient in a semiarid sandy grass-land. Acta Oecologica 69, 1–8 (2015).
Li, L. J. et al. Effects of nitrogen addition on grassland spe-cies diversity and productivity in Keerqin Ssndy Land. Chinese. Journal of Ap-plied Ecology 20, 1838–1844 (2009).
Clark, C. M. & Tilman, D. Loss of plant species after chronic low-level nitrogen deposition to prairie grass-lands. Nature 451, 712–715 (2008).
Liu, X. J. et al. Enhanced nitrogen deposition over China. Nature 494, 459–462 (2013).
Yahdjian, L. & Sala, O. E. A rainout shelter design for intercepting different amounts of rainfall. Oecologia 133, 95–101 (2002).
Wilcox, K. R., Fischer, J. C., Muscha, J. M., Petersen, M. K. & Knapp, A. K. Contrasting above- and be-lowground sensitivity of three Great Plains grasslands to altered rainfall regimes. Global Chang. Biology 21, 335–344 (2015).
Cornelissen, J. H. C., Lavorel, S. & Garnier, E. A handbook of protocols for standardised and easy measure-ment of plant functional traits worldwide. Australian Journal of Botany 51(4), 335 (2003).
Xiong, P., Shu, J., He, Z., Zhao, J. & Xu, B. Small rainfall pulses affected leaf photosynthesis rather than biomass production of dominant species in semiarid grassland community on Loess Plateau of China. Functional Plant Biology 44, 1229–1242 (2017).
Victoria, F. B., Jairo, A. P., Chen, Y. L. & Kadambot, H. M. Early season drought largely re-duces grain yield in wheat cultivars with smaller root systems. Plants 8, 305 (2019).
Liu, Y. B. et al. Anatomical, morphological and metabolic acclimation in the resurrection plant Reaumuria soongorica during dehydration and rehydration. Journal of Arid Environments 70, 183–194 (2007).
Zhang, Z., Shan, L. & Li, Y. Prolonged dry periods between rainfall events shorten the growth period of the resurrection plant Reaumuria soongorica. Ecology and Evolution 8, 920–927 (2017).
Ylhäisi, J. S. et al. Growing season precipitation in Finland under recent and projected climate. Natural Hazards and Earth System Sciences 10, 1563–1574 (2010).
Luo, Y. Q., Zhao, X. Y., Zuo, X. A., Li, Y. L. & Wang, T. Plant responses to warming and increased pre-cipitation in three categories of dune stabilization in northeastern China. Ecologi-cal. Research 4, 1–12 (2017).
Magill, A. H. et al. Long-Term Nitrogen Additions and Nitrogen Saturation in Two Temperate Forests. Ecosystems 3, 238–253 (2000).
Mcconnaughay, K. D. M. & Coleman, J. S. Biomass allocation in plants: ontogeny or optimality? A test alone three reource grasients. Ecology 80, 2581–2593 (1999).
Lü, X. T., Dijkstra, F. A., Kong, D. L., Wang, Z. W. & Han, X. G. Plant nitrogen uptake drives responses of productivity to nitrogen and water addition in a grassland. Scientific Reports 4, 4817 (2014).
Wu, G., Chen, M., Zhou, X. & Wang, Y. Response of morphological plasticity of three herbaceous seedlings to light and nutrition in the Qing Hai Tibetan Plateau. Asian Journal of Plant Sciences 5, 635–642 (2006).
Poorter, H. & Nagel, O. The role of biomass allocation in the growth response of plants to different levels of light, CO2, nutrients and water: a quantitative review. Australian Journal of Plant Physiology 27, 595–607 (2000).
Xu, B. C., Xu, W. Z., Huang, J., Shan, L. & Li, F. M. Biomass allocation, relative compet-itive ability and water use efficiency of two dominant species in semiarid Loess Plateau under water stress. Plant Sci-ence 181, 644–651 (2011).
Meier, I. C. & Leuschner, C. Belowground drought response of european beech: fine root biomass and carbon partitioning in. Global Change Biology 14, 2081-2095 (2010).
Huang, Y. X. et al. Allometric effects of agriophyllum squarrosum in response to soil nutrients, water, and popula-tion density in the Horqin Sandy Land of China. Journal of Plant Biology 52, 210–219 (2009).
Jones, S. K., Collins, S. L., Blair, J. M., Smith, M. D. & Knapp, A. K. Altered rainfall pat-terns increase forb abundance and richness in native tallgrass prairie. Scientific Reports 6, 20120 (2016).
Loreau, M. Biodiversity and ecosystem functioning: recent theoretical advances. Oikos 91, 3–17 (2000).
Zong, N. et al. Responses of eco-system respiration to nitrogen enrichment and clipping mediated by soil acidification in an alpine meadow. Pedobiologia (2016).
Chen, D. M., Lan, Z. C., Bai, X., Grace, J. B. & Bai, Y. F. Evidence that acidification-induced de-clines in plant diversity and productivity are mediated by changes in below-ground 25 communities and soil properties in a semi-arid steppe. Journal of Ecology. 101, 1322–1334 (2013).
Bi, J., Zhang, N. L., Liang, Y., Yang, H. J. & Ma, K. P. Interactive effects of water and nitrogen addition on soil microbial communities in a semiarid steppe. J. Plant Ecol. 5, 320–329 (2012).
Bodner, G., Leitner, D. & Kaul, H. P. Coarse and fine root plants affect pore size distribu-tions differently. Plant and Soil 380, 133–151 (2014).
Mccarthy, M. C. & Enquist, B. J. Consistency between an allometric approach and optimal partitioning theory in global patterns of plant biomass allocation. Functional Ecology 21, 713–720 (2007).
Acknowledgements
This study was supported by National Key Research and Development Program of China (2016YFC0503706) and National Natural Science Foundation of China (41622103 and 41571106). We would like to thank all the members of Naiman Desertification Research Station, Chinese Academy of Sciences, for their assistance with fieldwork.
Author information
Authors and Affiliations
Contributions
J.Z. organized and wrote the paper. X.Z., X.Z. analyzed the results. X.Z., X.Z. and J.M helped analyze the results. E.M. revised the language. All authors reviewed the manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Zhang, J., Zuo, X., Zhao, X. et al. Effects of rainfall manipulation and nitrogen addition on plant biomass allocation in a semiarid sandy grassland. Sci Rep 10, 9026 (2020). https://doi.org/10.1038/s41598-020-65922-0
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-020-65922-0
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
