
Abstract
Multidisciplinary approaches have demonstrated that the brain is potentially modulated by the long-term acquisition and practice of specific skills. Chess playing can be considered a paradigm for shaping brain function, with complex interactions among brain networks possibly enhancing cognitive processing. Dynamic network analysis based on resting-state magnetic resonance imaging (rs-fMRI) can be useful to explore the effect of chess playing on whole-brain fluidity/dynamism (the chronnectome). Dynamic connectivity parameters of 18 professional chess players and 20 beginner chess players were evaluated applying spatial independent component analysis (sICA), sliding-time window correlation, and meta-state approaches to rs-fMRI data. Four indexes of meta-state dynamic fluidity were studied: i) the number of distinct meta-states a subject pass through, ii) the number of switches from one meta-state to another, iii) the span of the realized meta-states (the largest distance between two meta-states that subjects occupied), and iv) the total distance travelled in the state space. Professional chess players exhibited an increased dynamic fluidity, expressed as a higher number of occupied meta-states (meta-state numbers, 75.8 ± 7.9 vs 68.8 ± 12.0, p = 0.043 FDR-corrected) and changes from one meta-state to another (meta-state changes, 77.1 ± 7.3 vs 71.2 ± 11.0, p = 0.043 FDR-corrected) than beginner chess players. Furthermore, professional chess players exhibited an increased dynamic range, with increased traveling between successive meta-states (meta-state total distance, 131.7 ± 17.8 vs 108.7 ± 19.7, p = 0.0004 FDR-corrected). Chess playing may induce changes in brain activity through the modulation of the chronnectome. Future studies are warranted to evaluate if these potential effects lead to enhanced cognitive processing and if “gaming” might be used as a treatment in clinical practice.
Similar content being viewed by others

Introduction
Neuroplasticity has always been considered as one of the most intriguing characteristics of the human brain1,2,3,4. In the last years, multidisciplinary research efforts have progressively demonstrated that the brain is potentially modulable by the long-term acquisition and practice of specific skills5,6. From this point of view, several studies have clearly reported how chess playing can be considered a paradigm that may induce long term changes in the brain. Indeed, advanced chess players can simulate/imagine the best next moves starting from a reservoir of chess patterns (chunks)7 through the involvement of high-level cognitive functions such as planning future actions, visuo-spatial perception, working memory, problem solving, judgment and decision making, and selection of previously acquired schemes7,8,9,10,11,12.
Continuous practice can boost these specific cognitive processes, with consequent enhancement of neuroplasticity mechanisms13,14. In particular, neuroimaging studies have shed light on how the brain is engaged during chess playing7,15,16 and how the brain undergoes long term reshaping due to practice8,9,10,17. Interestingly, chess players show complex interactions among brain networks with i) greater deactivation of the default mode network (DMN) and enhanced striatal-DMN integration during problem-solving8,9, ii) increased hub functional connectivity between the posterior fusiform gyrus and visuospatial attention and motor networks17,18, and iii) a more efficient whole-brain organization (increased small-world topology)10 as compared to healthy volunteers not engaged in chess playing.
However, functional connectivity approaches usually rely on the conceptual framework that the functional coupling among brain regions is a static feature, with no change over short periods of time19,20,21. In the last few years, this paradigm has been shown to be simplistic, as methodological approaches able to study the human brain as an interacting dynamic system have been developed (dynamic functional network connectivity (dFNC), the chronnectome)22,23,24,25,26,27,28. Cross-network correlations on successive sliding windows from the original scan-length network time-courses have revealed reproducible reccurring patterns of brain functional connectivity (time-varying connectivity)24,29,30. Recently, to incorporate key features of dynamic functional connectivity, the meta-state approach has been proposed31: for each subject, at a given point in time, the weighted probability to be in more than one state (distribution of probability of meta-state)31 can be defined. Briefly, with this more flexible approach multiple states might be represented to varying degrees at the same point in time, exhibiting lesser distortion in the features under investigation since contributions of all overlapping states are considered32, also providing a much more condensed summary measure of dynamic functional connectivity. In this view, objective measures of meta-state dynamic fluidity may be computed, such as the number of meta-states a subject passes through or the overall distance travelled by each subject through the state space. This highly reproducible approach31,33 has been already applied to schizophrenia31 and to neurodegenerative disorders34 and has helped in further understanding the neural basis of brain functioning22,35,36,37,38.
These premises set the stage for the present work, where we used the meta-state dynamic connectivity approach to explore differences between professional and beginner chess players, considering whole-brain fluidity/dynamism (the chronnectome).
Methods
Subjects
Data were derived from a public dataset on chess players39 encompassing professional chess players with a regular training (training time: 4.17 ± 1.72 h/day) and a control group of sex- and age-matched beginner chess players, who knew rules and simple strategies of Chinese chess. Dataset access was made from the “B1000 Functional Connectomes Project” (https://www.nitrc.org/projects/fcon_1000/). Subjects were recruited from the First National Intelligence Games held in Chengdu, China. All participants were right-handed and had no history of psychiatric or neurological disorders. No differences on observation skills or clear-thinking ability were found between these two groups, as already published39. Written informed consent was obtained from each subject and approval was obtained through the local Institutional Review Board of the West China Hospital of Sichuan University. Studies performed on this dataset are in accordance with relevant guidelines and regulations. Detailed information on this dataset has been reported in Li and colleagues’ study39.
Magnetic resonance Imaging (MRI) acquisition and preprocessing
MRI data were acquired on a 3T Siemens Trio system (Erlangen, German) at the MR Research Center of West China Hospital of Sichuan University, Chengdu, China. All MRI scans were performed when subjects were relaxed with their eyes open and fixated on a cross-hair centered on the screen. A T2-weighted gradient- echo echo-planar pulse sequence was used to obtain functional MRI (fMRI) images. A total of 205 whole brain echo-planar pulse sequence volumes were acquired using the following parameters: TR = 2000 ms, TE = 30 ms, flip angle = 90°, axial slice thickness = 5 mm, with no gap, slice number = 30, voxel size = 3.75 × 3.75 × 5 mm3. Functional preprocessing was carried out as previously published (for each subject, the first 5 volumes of the fMRI series were removed to account for magnetization equilibration; the remaining 200 volumes (total acquisition time: 6 minutes and 40 seconds) underwent slice-timing correction and were realigned to the first volume)34, using the toolbox for data preprocessing and analysis for brain imaging (DPABI, http://rfmri.org/dpabi)40 based on the Statistical Parametric Mapping (SPM12) software. Taking into account the significant impact of head motion on resting-state fMRI41,42, we considered absolute (mean translation and mean rotation) and relative (framewise displacement, FD-P)42 motion parameters. We consequently applied four levels of correction based on motion parameters: i) the absolute motion cut-off: any subject who had an absolute maximum displacement in any direction larger than 2.5 mm, or a maximum rotation (x,y,z) larger than 2.5°, was excluded from the study; ii) the relative motion cut-off: the framewise displacement of head position index (FD-P) (calculated as the sum of the absolute values of the 6 translational and rotational realignment parameters)42 with a cut-off of mean FD ≤ 0.2 mm, excluding subjects beyond this limit; iii) 12-motion parameters (6 original motion parameters and the 6 first-order derivatives) were applied on networks time-courses: the single time-courses were detrended (to remove baseline drifts from the scanners and/or physiological pulsations) and orthogonalized with respect to 12-motion parameters; and iv) the FD-P index previously calculated42 for each subject was included as nuisance variable in the final statistical analysis. Data were subsequently spatially normalized to the EPI unified segmentation template in Montreal Neurological Institute coordinates derived from SPM12 software and resampled to 3 × 3 × 3 mm3 cubic voxels. Spatial smoothing with an isotropic Gaussian kernel with full width at half-maximum (FWHM) 10 mm was applied.
Functional network decomposition
As previously reported, the functional imaging data were preprocessed using GIFT (GIFT toolbox, http://trendscenter.org/software/gift)43 and a spatially constrained ICA algorithm44. Spatially constrained ICA was used to compute intrinsic connectivity networks (ICNs) that corresponded to those from a previous analysis on a very large dataset of healthy subjects for test-retest reliability (37 ICNs derived from 7500 healthy subjects as spatial references, see Supplementary Fig. 1 for the visualization of the spatial maps of the ICNs used)34,45,46,47. Subject-specific spatial patterns and time-courses were calculated and then converted to Z-scores. As already described above, the single time-courses were detrended, orthogonalized with respect to 12-motion parameters, despiked (replacement of outlier time points with 3rd order spline fitting to clean neighboring points) and filtered using a 5th order Butterworth filter (0.01–0.15 Hz)45.
Meta-state computation
Dynamic functional network connectivity (dFNC) was obtained using the dynamic FNC toolbox implemented in GIFT48. dFNC was calculated using a sliding-window approach to estimate functional connectivity between ICNs for each segment. Segments were defined with a tapered window convolving a rectangle (width = 30, TRs = 60 s) with a Gaussian (σ = 3 TRs) and slides in steps of 1 TR. A LASSO approach with L1 regularization (100 repetitions) was used to compute the covariance between the independent component (IC) time-courses. For each subject, the optimal regularization parameter (λ) obtained using cross-validation was defined, as previously published29,49. To decompose dFNC into connectivity patterns (CPs, meta-states), the sICA approach was applied, considering a number of CPs equal to 5, in line with previous work on meta-states in dynamic brain connectivity, to have a good balance to take into account complex linearly additive effects and to retain richly featured basis pattern31,33,34. As previously described, the time-courses were discretized (to work over a more tractable space) into 8 bins (positive and negative quartiles) and each timepoint was ended into a meta-state33. The time-courses for sICA CPs were derived from the regression of each subject’s dFNC information at each time window on the group of sICA CPs.
In line with meta-states calculation, each subject will have a weighted probability to be in more than one state, and this time-varying distribution of probability represents the methodological underpinning to obtain effective measures of meta-state dynamic fluidity.
For this purpose, four indexes of connectivity dynamism were herein explored: i) the number of distinct meta-states the subjects occupied during their scans (meta-state number); ii) the number of times that subjects switched from one meta-state to another (meta-state changes); iii) the largest distance between two meta-states that subjects occupied (meta-state span); and iv) the overall distance travelled by each subject through the state space (the sum of the L1 distances between successive meta-states, i.e., meta-state total distance)33. Moreover, to further test the robustness of the approach on the present data we also considered different model dimensionality (using a number of CPs from 4 to 8): for each number of CPs, the statistical comparisons between professional chess players and beginner chess players were performed.
Statistical analysis
Comparisons of demographic and clinical characteristics among groups (professional chess players vs. beginner chess players) were assessed by Mann-Whitney U test for continuous variables and χ2 test for categorical variables. A general linear model (GLM) considering gender and FD-P as nuisance variables was applied to study dFNC in the two groups. Partial Pearson’s correlation analysis was used to assess the relationship between the meta-state measures (meta-state number, meta-state changes, meta-state span and meta-state total distance) and the total amount of time spent by professional chess players for training, corrected for gender and FD. Statistical analyses were performed by using SPSS software (IBM SPSS Statistics 22.0, Chicago, USA) and statistical significance was set at p < 0.05, considering correction for multiple comparisons (Benjamini-Hochberg False Discovery Rate-FDR correction) for indexes of connectivity dynamism50.
Results
Considering the original group39 of 29 professional chess players and 29 beginner chess players, 20 subjects were excluded for technical reasons, namely 2 beginner chess players for different MRI protocol (number of slices), 1 professional chess player for excessive absolute motion, 7 beginner chess players and 10 professional chess players for excessive relative motion (FD > 0.2 mm). Thus, 38 subjects (18 professional chess players and 20 beginner chess players) were considered (Table 1).
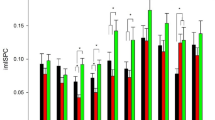
We considered five connectivity patterns (CPs) of dFNC, which are reported in Fig. 1. dFNC was expressed as a weighted sum of the discretized five-dimensional CPs, considering that 85 = 32,768 distinct five-dimensional meta-states in our signed quartile discretization were present, with a mean number of occupied meta-states in the overall group of 72.1 ± 10.8 (0.22% of the total). Professional chess players showed greater dynamic fluidity, as they occupied a higher number of meta-states (i.e., meta-state numbers, p = 0.043 FDR-corrected) and changed from one meta-state to another more often (i.e., meta-state changes, p = 0.043 FDR-corrected) than beginner chess players (see Table 2 and Supplementary Fig. 2). Furthermore, professional chess players operated over an increased dynamic range with increased meta-state total distance (p = 0.0004 FDR-corrected), as they travelled more overall distance, between successive meta-states, through the state space than beginner chess players (see Table 2 and Supplementary Fig. 2). We did not find a statistically significant difference in meta-state span between groups. Considering different numbers of CPs (ranging from 4 to 8) the statistical differences between professional and beginner chess players were quite stable, in particular for meta-state total distance (see Supplementary Table 1 for details). In Fig. 2, meta-state dynamics through time, meta-state numbers, meta-state change points, and meta-state total distance in a representative beginner chess player (on the left) and in a representative professional chess player (on the right) were reported. The representative professional chess player showed a greater brain dynamism, as compared to the beginner chess player (panel A), as suggested by the more complex pattern in the former subject, with a higher number of realized meta-states (panel B), meta-state changes (panel C), and greater travelled overall distance (panel D).

The five connectivity patterns (CPs) resulting from the dynamic Functional Network Connectivity (dFNC) analysis. The five correlation matrices are reported, in which each square represents one of the 37 considered network components. The colors of each CP represent the direction and the strength of the relationship between each dFNC pair and time-course of the CP (red: positive correlation of the time-course; blue: negative correlation of the time-course).

Meta-state dynamics through time, meta-state numbers, meta-state changes, and meta-state total distance in a representative beginner chess player and a representative professional chess player. Meta-state dynamics through time (panel A), meta-state numbers (panel B), meta-state change points (panel C), and meta-state total distance (panel D) in a representative beginner chess player (left column) and in a representative professional chess player (right column). The colorbar represents the strength of probability to be in each meta-state. For panel A and B Y-axis represents the five connectivity patterns (CPs), from 1 to 5 and X-axis represented the time-indexed meta-states (seconds, after time-course discretization in quartiles). For panel C Y-axis represents the distance of each changepoint, whereas the sum of all the blue dots represents the cumulative number of changepoints for a given subject. Finally, for panel D the total cumulative distance traveled (summed L1 distance between successive meta-states) in the state space is reported on Y-axis. y = years; y.o.= years old.
No significant correlation between time spent by professional chess players for daily training and meta-state measures (partial Pearson’s correlation analysis corrected for gender and FD-P: meta-state number: r = 0.150, p = 0.580; meta-state changes: r = 0.195, p = 0.469; meta-state span: r = 0.295, p = 0.268; meta-state total distance: r = 0.055, p = 0.839) was found.
In the whole group no significant correlation between education and meta state measures (partial Pearson’s correlation analysis corrected for gender, chess group and FD-P; meta-state number: r = 0.028, p = 0.872; meta-state changes: r = 0.067, p = 0.661; meta-state span: r = −0.023, p = 0.895; meta-state total distance: r = 0.195, p = 0.261)) was found.
Discussion
Different board games have been taken into consideration as potential tools to reinforce/strength cognitive abilities in either healthy or diseased brain51,52,53,54. Among all, the game of chess has been assumed as a paradigm in the field, especially because of its high diffusion worldwide with a significant number of professional players55. Several brain hotspots have been demonstrated to be selectively strengthen throughout practice of chess playing, mainly in the basal ganglia, the fusiform gyrus, the default mode network and the attention network8,9,17. This reflects the specific abilities employed by professional players/grand masters, based on the simulation and planning of the next best move using previously acquired chess pattern schemes56.
Our study tries to move forward, with the idea that regardless of the specific involved brain hubs/networks8,9,17, chess playing can potentially modulate whole-brain activity, shaping the spontaneous and time-varying fluctuation of brain signal22,35,37,57. In fact, compared to beginners, professional chess players demonstrated enhanced global dynamic fluidity (with a higher number of occupied meta-states, an increased number of changes from one meta-state to another), operating over an extended dynamic range (increased meta-state total distance travelled between successive meta-states). This is in line with a previous study demonstrating an increased small-world topology in chess players, reflecting an optimized cost efficiency of information processing as well as an optimal global network organization as a result of cognitive training10. It is interesting to note that static and dynamic brain connectivity are intimately linked, the fine-grained and multi-level organization of the brain based on the spontaneous fluctuation of brain signal21,58,59. Considering the theoretic framework of meta-states, these findings suggest that chess playing does not merely modulate brain areas strictly involved in chess practice (such as visuo-attentional network or frontal executive functions) but is able to shape whole brain functioning at different spatial and temporal scales (as suggested in particular by meta-state total distance modulation). Plasticity induced by practice produces multiple structural change in the brain such as myelin reorganization and formation, dendritic branching and synaptogenesis60 which boost the effectivity in the neural communication. This process may maximize neuronal tuning, resulting in magnifying transmission and information capacity to a critical optimal point61. From this point of view, dynamic functional connectivity and its related measures could be more effective in detecting this functional reorganization.
Furthermore, the impact on brain fluidity and dynamism (as measured by meta-state indexes) is not directly related to the total amount of time spent by chess players in training as well as it is not related to education. Even if the lack of a significant correlation could be related to the small sample size, it might be hypothesized an add-on modulating effect of chess playing, along with the well-established role of education, on cognitive reserve mechanisms62,63,64. Thus, the increased global brain dynamic fluidity observed in professional chess players could provide a direct evidence of neuroplasticity mechanisms related to long-term skill acquisition, potentially representing cognitive reserve enhancement65,66,67,68,69,70. In this view, chess playing may be considered a proxy measure of cognitive reserve along with education, occupation attainment or leisure activities71,72,73 and in some way may protect the brain from physiological or pathological aging74.
We acknowledge that this study entails some limits. First, we considered a small number of subjects and further studies are warranted to validate the present findings, considering different level of confounders (age, educational level, etc.). Moreover, impact of head movement on dynamic functional connectivity needs to be further addressed, although in this study we applied strict head motion parameters to exclude this possible bias.
In conclusion, this study demonstrated that chess playing may ameliorate whole brain functioning and increase whole brain dynamic fluidity. Further studies (collecting data with more time-points as well as a higher spatial resolution and applying different dynamic connectivity methodological approaches75,76, in different populations and in other board games) may further show how the brain can react and modify itself through the continuous practice of high-level activities with long-term skill acquisition77.
References
Wenger, E., Brozzoli, C., Lindenberger, U. & Lovden, M. Expansion and Renormalization of Human Brain Structure During Skill Acquisition. Trends Cogn. Sci. 21, 930–939, https://doi.org/10.1016/j.tics.2017.09.008 (2017).
DeFelipe, J. Brain plasticity and mental processes: Cajal again. Nat. Rev. Neurosci. 7, 811–817, https://doi.org/10.1038/nrn2005 (2006).
Chapman, S. B. et al. Neural mechanisms of brain plasticity with complex cognitive training in healthy seniors. Cereb. Cortex 25, 396–405, https://doi.org/10.1093/cercor/bht234 (2015).
Cramer, S. C. et al. Harnessing neuroplasticity for clinical applications. Brain 134, 1591–1609, https://doi.org/10.1093/brain/awr039 (2011).
May, A. Experience-dependent structural plasticity in the adult human brain. Trends Cogn. Sci. 15, 475–482, https://doi.org/10.1016/j.tics.2011.08.002 (2011).
Lewis, C. M., Baldassarre, A., Committeri, G., Romani, G. L. & Corbetta, M. Learning sculpts the spontaneous activity of the resting human brain. Proc. Natl Acad. Sci. USA 106, 17558–17563, https://doi.org/10.1073/pnas.0902455106 (2009).
Wan, X. et al. The neural basis of intuitive best next-move generation in board game experts. Science 331, 341–346, https://doi.org/10.1126/science.1194732 (2011).
Duan, X. et al. Reduced caudate volume and enhanced striatal-DMN integration in chess experts. Neuroimage 60, 1280–1286, https://doi.org/10.1016/j.neuroimage.2012.01.047 (2012).
Duan, X. et al. Large-scale brain networks in board game experts: insights from a domain-related task and task-free resting state. PLoS One 7, e32532, https://doi.org/10.1371/journal.pone.0032532 (2012).
Duan, X. et al. Functional organization of intrinsic connectivity networks in Chinese-chess experts. Brain Res. 1558, 33–43, https://doi.org/10.1016/j.brainres.2014.02.033 (2014).
Powell, J. L., Grossi, D., Corcoran, R., Gobet, F. & Garcia-Finana, M. The neural correlates of theory of mind and their role during empathy and the game of chess: A functional magnetic resonance imaging study. Neuroscience 355, 149–160, https://doi.org/10.1016/j.neuroscience.2017.04.042 (2017).
Villafaina, S., Collado-Mateo, D., Cano-Plasencia, R., Gusi, N. & Fuentes, J. P. Electroencephalographic response of chess players in decision-making processes under time pressure. Physiol. Behav. 198, 140–143, https://doi.org/10.1016/j.physbeh.2018.10.017 (2019).
Aciego, R., Garcia, L. & Betancort, M. The benefits of chess for the intellectual and social-emotional enrichment in schoolchildren. Span. J. Psychol. 15, 551–559 (2012).
van der Maas, H. L. & Wagenmakers, E. J. A psychometric analysis of chess expertise. Am. J. Psychol. 118, 29–60 (2005).
Bilalic, M., Langner, R., Ulrich, R. & Grodd, W. Many faces of expertise: fusiform face area in chess experts and novices. J. Neurosci. 31, 10206–10214, https://doi.org/10.1523/jneurosci.5727-10.2011 (2011).
Rennig, J., Bilalic, M., Huberle, E., Karnath, H. O. & Himmelbach, M. The temporo-parietal junction contributes to global gestalt perception-evidence from studies in chess experts. Front. Hum. Neurosci. 7, 513, https://doi.org/10.3389/fnhum.2013.00513 (2013).
Song, L., Peng, Q., Liu, S. & Wang, J. Changed hub and functional connectivity patterns of the posterior fusiform gyrus in chess experts. Neurotherapeutics, https://doi.org/10.1007/s11682-018-0020-0 (2019).
Langner, R., Eickhoff, S. B. & Bilalic, M. A network view on brain regions involved in experts’ object and pattern recognition: Implications for the neural mechanisms of skilled visual perception. Brain Cogn. 131, 74–86, https://doi.org/10.1016/j.bandc.2018.09.007 (2019).
Damoiseaux, J. S. et al. Consistent resting-state networks across healthy subjects. Proc. Natl Acad. Sci. USA 103, 13848–13853, https://doi.org/10.1073/pnas.0601417103 (2006).
De Luca, M., Beckmann, C. F., De Stefano, N., Matthews, P. M. & Smith, S. M. fMRI resting state networks define distinct modes of long-distance interactions in the human brain. Neuroimage 29, 1359–1367, https://doi.org/10.1016/j.neuroimage.2005.08.035 (2006).
Liu, J., Liao, X., Xia, M. & He, Y. Chronnectome fingerprinting: Identifying individuals and predicting higher cognitive functions using dynamic brain connectivity patterns. 39, 902–915, https://doi.org/10.1002/hbm.23890 (2018).
Calhoun, V. D., Miller, R., Pearlson, G. & Adali, T. The chronnectome: time-varying connectivity networks as the next frontier in fMRI data discovery. Neuron 84, 262–274, https://doi.org/10.1016/j.neuron.2014.10.015 (2014).
Canolty, R. T. & Knight, R. T. The functional role of cross-frequency coupling. Trends Cogn. Sci. 14, 506–515, https://doi.org/10.1016/j.tics.2010.09.001 (2010).
Chang, C. & Glover, G. H. Time-frequency dynamics of resting-state brain connectivity measured with fMRI. Neuroimage 50, 81–98, https://doi.org/10.1016/j.neuroimage.2009.12.011 (2010).
Fries, P. A mechanism for cognitive dynamics: neuronal communication through neuronal coherence. Trends Cogn. Sci. 9, 474–480, https://doi.org/10.1016/j.tics.2005.08.011 (2005).
Hillebrand, A. et al. Direction of information flow in large-scale resting-state networks is frequency-dependent. Proc. Natl Acad. Sci. USA 113, 3867–3872, https://doi.org/10.1073/pnas.1515657113 (2016).
Sakoglu, U. et al. A method for evaluating dynamic functional network connectivity and task-modulation: application to schizophrenia. Magma 23, 351–366, https://doi.org/10.1007/s10334-010-0197-8 (2010).
Keilholz, S., Caballero-Gaudes, C., Bandettini, P., Deco, G. & Calhoun, V. Time-Resolved Resting-State Functional Magnetic Resonance Imaging Analysis: Current Status, Challenges, and New Directions. Brain Connect. 7, 465–481, https://doi.org/10.1089/brain.2017.0543 (2017).
Allen, E. A. et al. Tracking whole-brain connectivity dynamics in the resting state. Cereb. Cortex 24, 663–676, https://doi.org/10.1093/cercor/bhs352 (2014).
Marusak, H. A. et al. Dynamic functional connectivity of neurocognitive networks in children. Hum. Brain Mapp. 38, 97–108, https://doi.org/10.1002/hbm.23346 (2017).
Miller, R. L. et al. Higher Dimensional Meta-State Analysis Reveals Reduced Resting fMRI Connectivity Dynamism in Schizophrenia Patients. PLoS One 11, e0149849, https://doi.org/10.1371/journal.pone.0149849 (2016).
Leonardi, N., Shirer, W. R. & Greicius, M. D. & Van De Ville, D. Disentangling dynamic networks: Separated and joint expressions of functional connectivity patterns in time. Hum. Brain Mapp. 35, 5984–5995, https://doi.org/10.1002/hbm.22599 (2014).
Miller, R. L., Yaesoubi, M. & Calhoun, V. D. Higher dimensional analysis shows reduced dynamism of time-varying network connectivity in schizophrenia patients. Conf. Proc. IEEE Eng. Med. Biol. Soc. 2014, 3837–3840, https://doi.org/10.1109/embc.2014.6944460 (2014).
Premi, E. et al. The inner fluctuations of the brain in presymptomatic Frontotemporal Dementia: The chronnectome fingerprint. Neuroimage 189, 645–654, https://doi.org/10.1016/j.neuroimage.2019.01.080 (2019).
Linaro, D., Ocker, G. K., Doiron, B. & Giugliano, M. Correlation transfer by layer 5 cortical neurons under recreated synaptic inputs in vitro. J. Neurosci. 39, 7648–7663, https://doi.org/10.1523/JNEUROSCI.3169-18.2019 (2019).
Doiron, B. & Litwin-Kumar, A. The mechanics of state-dependent neural correlations. Nat. Neurosci. 19, 383–393, https://doi.org/10.1038/nn.4242 (2016).
Preti, M. G., Bolton, T. A. & Van De Ville, D. The dynamic functional connectome: State-of-the-art and perspectives. Sci. Rep. 160, 41–54, https://doi.org/10.1038/s41598-017-12993-1 (2017).
Preti, M. G. & Van De Ville, D. Dyn. Funct. connectivity high. Spat. Resolut. Reveal. long-range Interact. fine-scale organization. 7, 12773, https://doi.org/10.1038/s41598-017-12993-1 (2017).
Li, K. et al. A multimodal MRI dataset of professional chess players. Sci. Data 2, 150044, https://doi.org/10.1038/sdata.2015.44 (2015).
Yan, C. G., Wang, X. D., Zuo, X. N. & Zang, Y. F. DPABI: Data Processing & Analysis for (Resting-State) Brain Imaging. Neuroinformatics 14, 339–351, https://doi.org/10.1007/s12021-016-9299-4 (2016).
Parkes, L., Fulcher, B., Yucel, M. & Fornito, A. An evaluation of the efficacy, reliability, and sensitivity of motion correction strategies for resting-state functional MRI. Neuroimage 171, 415–436, https://doi.org/10.1016/j.neuroimage.2017.12.073 (2018).
Power, J. D., Barnes, K. A., Snyder, A. Z., Schlaggar, B. L. & Petersen, S. E. Spurious but systematic correlations in functional connectivity MRI networks arise from subject motion. Neuroimage 59, 2142–2154, https://doi.org/10.1016/j.neuroimage.2011.10.018 (2012).
Calhoun, V. D., Adali, T., Pearlson, G. D. & Pekar, J. J. A method for making group inferences from functional MRI data using independent component analysis. Hum. Brain Mapp. 14, 140–151 (2001).
Wang, C., Ong, J. L., Patanaik, A., Zhou, J. & Chee, M. W. Spontaneous eyelid closures link vigilance fluctuation with fMRI dynamic connectivity states. Proc. Natl Acad. Sci. USA 113, 9653–9658, https://doi.org/10.1073/pnas.1523980113 (2016).
Abrol, A., Chaze, C., Damaraju, E. & Calhoun, V. D. The chronnectome: Evaluating replicability of dynamic connectivity patterns in 7500 resting fMRI datasets. Conf. Proc. IEEE Eng. Med. Biol. Soc. 2016, 5571–5574, https://doi.org/10.1109/embc.2016.7591989 (2016).
Abrol, A. et al. Replicability of time-varying connectivity patterns in large resting state fMRI samples. Neuroimage 163, 160–176, https://doi.org/10.1016/j.neuroimage.2017.09.020 (2017).
Salman, M. S. et al. Group ICA for identifying biomarkers in schizophrenia: ‘Adaptive’ networks via spatially constrained ICA show more sensitivity to group differences than spatio-temporal regression. Neuroimage Clin. 22, 101747, https://doi.org/10.1016/j.nicl.2019.101747 (2019).
Damaraju, E. et al. Dynamic functional connectivity analysis reveals transient states of dysconnectivity in schizophrenia. Neuroimage Clin. 5, 298–308, https://doi.org/10.1016/j.nicl.2014.07.003 (2014).
Friedman, J., Hastie, T. & Tibshirani, R. Sparse inverse covariance estimation with the graphical lasso. Biostatistics 9, 432–441, https://doi.org/10.1093/biostatistics/kxm045 (2008).
Benjamini, Y. & Hochberg, Y. Controlling the false discovery rate: a practical and powerful approach to multiple testing. J. Roy. Statist. Soc. Ser. B 57, 289–300 (1995).
Dartigues, J. F. et al. Playing board games, cognitive decline and dementia: a French population-based cohort study. BMJ Open. 3, e002998, https://doi.org/10.1136/bmjopen-2013-002998 (2013).
Nakao, M. Special series on “effects of board games on health education and promotion” board games as a promising tool for health promotion: a review of recent literature. Biopsychosoc. Med. 13, 5, https://doi.org/10.1186/s13030-019-0146-3 (2019).
Hampshire, A., Sandrone, S. & Hellyer, P. J. A Large-Scale, Cross-Sectional Investigation Into the Efficacy of Brain Training. Front. Hum. Neurosci. 13, 221, https://doi.org/10.3389/fnhum.2019.00221 (2019).
Lin, Q., Cao, Y. & Gao, J. The impacts of a GO-game (Chinese chess) intervention on Alzheimer disease in a Northeast Chinese population. Front. Aging Neurosci. 7, 163, https://doi.org/10.3389/fnagi.2015.00163 (2015).
Vaci, N. & Bilalic, M. Chess databases as a research vehicle in psychology: Modeling large data. Behav. Res. Methods 49, 1227–1240, https://doi.org/10.3758/s13428-016-0782-5 (2017).
Gong, Y., Ericsson, K. A. & Moxley, J. H. Recall of briefly presented chess positions and its relation to chess skill. PLoS One 10, e0118756, https://doi.org/10.1371/journal.pone.0118756 (2015).
Lord, L. D., Stevner, A. B., Deco, G. & Kringelbach, M. L. Understanding principles of integration and segregation using whole-brain computational connectomics: implications for neuropsychiatric disorders. Philos Trans A Math Phys Eng Sci 375, https://doi.org/10.1098/rsta.2016.0283 (2017).
Xia, Y. et al. Tracking the dynamic functional connectivity structure of the human brain across the adult lifespan. Hum. Brain Mapp. 40, 717–728, https://doi.org/10.1002/hbm.24385 (2019).
Ma, Z. & Zhang, N. Temporal transitions of spontaneous brain activity. Elife 7, https://doi.org/10.7554/eLife.33562 (2018).
Zatorre, R. J., Fields, R. D. & Johansen-Berg, H. Plasticity in gray and white: neuroimaging changes in brain structure during learning. Nat. Neurosci. 15, 528–536, https://doi.org/10.1038/nn.3045 (2012).
Ma, Z., Turrigiano, G. G., Wessel, R. & Hengen, K. B. Cortical Circuit Dynamics Are Homeostatically Tuned to Criticality In Vivo. Neuron, https://doi.org/10.1016/j.neuron.2019.08.031 (2019).
Stern, Y. et al. Whitepaper: Defining and investigating cognitive reserve, brain reserve, and brain maintenance. Alzheimers Dement, https://doi.org/10.1016/j.jalz.2018.07.219 (2018).
Borroni, B., Premi, E., Bozzali, M. & Padovani, A. Reserve mechanisms in neurodegenerative diseases: from bench to bedside and back again. Curr. Med. Chem. 19, 6112–6118 (2012).
Marques, P. et al. The functional connectome of cognitive reserve. Hum. Brain Mapp. 37, 3310–3322, https://doi.org/10.1002/hbm.23242 (2016).
Rosenberg, A. et al. Multidomain lifestyle intervention benefits a large elderly population at risk for cognitive decline and dementia regardless of baseline characteristics: The FINGER trial. Alzheimers Dement. 14, 263–270, https://doi.org/10.1016/j.jalz.2017.09.006 (2018).
Groot, C. et al. The effect of physical activity on cognitive function in patients with dementia: A meta-analysis of randomized control trials. Ageing Res. Rev. 25, 13–23, https://doi.org/10.1016/j.arr.2015.11.005 (2016).
Vauzour, D. et al. Nutrition for the ageing brain: Towards evidence for an optimal diet. Ageing Res. Rev. 35, 222–240, https://doi.org/10.1016/j.arr.2016.09.010 (2017).
Perani, D. et al. The impact of bilingualism on brain reserve and metabolic connectivity in Alzheimer’s dementia. Proc. Natl Acad. Sci. USA 114, 1690–1695, https://doi.org/10.1073/pnas.1610909114 (2017).
Biasutti, M. & Mangiacotti, A. Assessing a cognitive music training for older participants: a randomised controlled trial. Int. J. Geriatr. Psychiatry 33, 271–278, https://doi.org/10.1002/gps.4721 (2018).
Hars, M., Herrmann, F. R., Gold, G., Rizzoli, R. & Trombetti, A. Effect of music-based multitask training on cognition and mood in older adults. Age Ageing 43, 196–200, https://doi.org/10.1093/ageing/aft163 (2014).
Stern, Y. Cognitive reserve in ageing and Alzheimer’s disease. Lancet Neurol. 11, 1006–1012, https://doi.org/10.1016/s1474-4422(12)70191-6 (2012).
Ihle, A., Oris, M., Fagot, D., Maggiori, C. & Kliegel, M. The association of educational attainment, cognitive level of job, and leisure activities during the course of adulthood with cognitive performance in old age: the role of openness to experience. Int. Psychogeriatr. 28, 733–740, https://doi.org/10.1017/s1041610215001933 (2016).
Foubert-Samier, A. et al. Education, occupation, leisure activities, and brain reserve: a population-based study. Neurobiol. Aging 33(423), e415–425, https://doi.org/10.1016/j.neurobiolaging.2010.09.023 (2012).
Altschul, D. M. & Deary, I. J. Playing analog games is associated with reduced declines in cognitive function: a 68 year longitudinal cohort study. J Gerontol B Psychol Sci Soc Sci, https://doi.org/10.1093/geronb/gbz149 (2019).
Iraji, A. et al. The spatial chronnectome reveals a dynamic interplay between functional segregation and integration. Hum. Brain Mapp. 40, 3058–3077, https://doi.org/10.1002/hbm.24580 (2019).
Iraji, A. et al. Spatial dynamics within and between brain functional domains: A hierarchical approach to study time-varying brain function. Hum. Brain Mapp. 40, 1969–1986, https://doi.org/10.1002/hbm.24505 (2019).
Vaci, N. et al. The joint influence of intelligence and practice on skill development throughout the life span. Proc. Natl Acad. Sci. USA 116, 18363–18369, https://doi.org/10.1073/pnas.1819086116 (2019).
Author information
Authors and Affiliations
Contributions
E.P. design of the work, statistical analysis and interpretation of the data, draft of the work. S.G. statistical analysis and interpretation of the data, draft of the work. M.D. statistical analysis and interpretation of the data, draft of the work. A.G. design of the work, revision of the manuscript. V.D.C. interpretation of the data, revision of the manuscript. AI: interpretation of the data, revision of the manuscript. Q.G. revision of the manuscript. K.L. revision of the manuscript. F.C. revision of the manuscript. R.G. revision of the manuscript. A.P. revision of the manuscript. B.B. interpretation of the data, draft of the work. M.M. revision of the manuscript.
Corresponding author
Ethics declarations
Competing interests
Authors have no competing interests as defined by Nature Research, or other interests that might be perceived to influence the results and/or discussion reported in this paper. M.D. is supported by an ERC Consolidator Grant 2017 (LIGHTUP, project # 772953, P.I. Prof. Marco Tamietto).
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Premi, E., Gazzina, S., Diano, M. et al. Enhanced dynamic functional connectivity (whole-brain chronnectome) in chess experts. Sci Rep 10, 7051 (2020). https://doi.org/10.1038/s41598-020-63984-8
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-020-63984-8
This article is cited by
-
Prediction of dyslexia and attention deficit and hyperactivity disorder prediction using ensemble classifier model
International Journal of System Assurance Engineering and Management (2022)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
