
Abstract
We have recently identified and characterized two pseudogenes (HMGA1P6 and HMGA1P7) of the HMGA1 gene, which has a critical role in malignant cell transformation and cancer progression. HMGA1P6 and HMGAP17 act as microRNA decoy for HMGA1 and other cancer-related genes upregulating their protein levels. We have previously shown that they are upregulated in several human carcinomas, and their expression positively correlates with a poor prognosis and an advanced cancer stage. To evaluate in vivo oncogenic activity of HMGA1 pseudogenes, we have generated a HMGA1P7 transgenic mouse line overexpressing this pseudogene. By a mean age of 12 months, about 50% of the transgenic mice developed splenomegaly and accumulation of lymphoid cells in several body compartments. For these mice FACS and immunohistochemical analyses suggested the diagnosis of B-cell lymphoma that was further supported by clonality analyses and RNA expression profile of the pathological tissues of the HMGA1P7 transgenic tissues. Therefore, these results clearly demonstrate the oncogenic activity of HMGA1 pseudogenes in vivo.
Similar content being viewed by others

Introduction
Many evidences indicate that long non-coding RNAs (lncRNAs) are key modulators of different biological phenomena. Given this scenario, it is predictable that deregulated expression and aberrant role of lncRNAs are involved in the development of several diseases including cancer1. Among lncRNAs, pseudogenes, a subgroup of genes that arises from protein-coding genes that have lost the capacity to produce proteins, have been considered for long time as non-functional genomic junk1. However, recent studies have unveiled important functions of pseudogenes in the regulation of the expression of the parental genes. Indeed, the majority of the identified pseudogenes has high sequence homology with their protein-coding parental counterparts, enabling them to take part in post-transcriptional control of their parental genes. The regulation of parental gene relies on several mechanisms: (i) the generation of endogenous short interfering RNAs (siRNAs)2,3; (ii) the engagement of regulatory proteins on the parental gene by pseudogene RNAs to control gene expression and chromatin remodelling4,5; (iii) the ability of the pseudogenes to compete with the parental genes for RNA-binding proteins and the translation machinery6,7,8; (iiii) the ability of pseudogenes to compete with their parental genes for a common pool of shared microRNAs (miRNAs)9 through the high sequence homology of the 3′ Untranslated region (UTR), thus regulating each other expression as competitive endogenous RNAs (ceRNAs)10.
The HMGA protein family includes the HMGA1a, HMGA1b and HMGA2 members11. The first two are coded for by the same gene through an alternative splicing. They have no transcriptional activity per se, but, modifying the chromatin architecture, they are able to positively or negatively regulate the expression of several genes, particularly those involved in cancer progression11,12. Consistently, these proteins are expressed at very low levels in normal adult tissues, but are abundant in almost all the human malignant neoplasms11, and their expression significantly correlates with the capability of cancer cells to metastatize and a patient poor prognosis13,14,15. Moreover, in vitro and in vivo models support a causal role of the HMGA proteins in cell transformation and cancer development11,16,17.
We have recently identified two human HMGA1 processed pseudogenes (HMGA1P6 and HMGA1P7) that are not present in mouse genome. HMGA1P6 and HMGA1P7 can compete with HMGA1 for miRNA binding, leading to the upregulation of HMGA1 cellular levels, thereby enhancing the expression of cell malignant features18,19,20,21,22,23. The overexpression of these HMGA1 pseudogenes (HMGA1Ps) also increases the levels of HMGA2 and other cancer-related genes, such as EZH2 and VEGF, by inhibiting the suppression of their synthesis18. Noteworthy, HMGA1Ps were found overexpressed in several human cancer types supporting their involvement in carcinogenesis18,20,21,22,23. To investigate the role of HMGA1 pseudogenes overexpression in vivo, we generated transgenic mouse model overexpressing HMGA1P7 (HMGA1P7-TG)18,22,23,24. Mouse Embryonic Fibroblasts (MEFs) derived from HMGA1 pseudogene transgenic mice showed a higher growth rate and a later onset of senescence than the wild-type (WT) counterpart18.
Here, we report that HMGA1 pseudogene transgenic mice develop haematological neoplasia characterized by monoclonal B-cell populations, most of them diagnosed as large B-cell lymphoma. These results validate the oncogenic role of the HMGA1 pseudogenes18.
Results
HMGA1P7 transgenic mice develop lymphoproliferative lesions
Transgenic mice carrying the HMGA1P7 gene were generated by the injection of the transgene into C57BL/6N derived-zygotes and, then transferred into pseudo-pregnant as previously described18. The expression of the HMGA1P7 was assessed in lungs, spleens and kidneys explanted from transgenic mice (Fig. 1).

Analysis of HMGA1P7 expression in transgenic mice qRT-PCR analysis of total RNA from lungs, spleens and kidneys of WT (n = 3) and HMGA1P7 (n = 3) transgenic mice. The error bars represent mean ± SD.
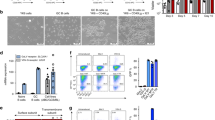
Interestingly, HMGA1P7 mice showed significant increased mortality with respect to the WT mice (Gehan Breslow Wilcoxon test, p < 0.0001) with a mean age of death of about 52 weeks (Fig. 2A). About 50% of 12 months-old HMGA1P7 transgenic mice displayed splenomegaly at necropsy, whereas WT mice showed no relevant alteration in splenic size or weight (Fig. 2B,C). Histological sections of the HMGA1P7- TG spleens showed a clear distinction between the red and the white pulp. In the red pulp multiple foci of extramedullary haematopoiesis, as well as hemosiderin-laden macrophages were frequently observed (Fig. 3A,III). White pulp showed a moderate expansion with some confluent areas and partial loss of normal structures and germinal centers. In some mice, higher magnification showed a diffuse, monotonous lymphoid population composed of medium-to-large rounded cells with scant cytoplasm, round to oval nuclei and single or multiple, prominent nucleoli often adherent to the nuclear membrane (Fig. 3A,IV). Mitotic activity was medium to high (<10 × 10 HPF). Intriguingly, histopathological analyses revealed monotonous lymphoid cells infiltrating liver (≈25%), kidneys (≈25%), lung (≈30%), and pancreas (≈20%) (Fig. 3B). Immunohistochemical analysis of lymphoid component displayed a predominant CD45/B220-positive population intermingled with few, scattered CD3-positive cells (Fig. 3C). Based on morphology and immunophenotype, a diagnosis of large B-cell lymphoma with immunoblastic features was made (human counterpart: DLBCL, immunoblastic variant)25.

HMGA1P7 expression in vivo induces splenomegaly and premature death (A) Survival curve of WT (n = 30) and HMGA1P7 (n = 40) transgenic mice. The survival rate of WT mice was significantly higher than HMGA1P7 transgenic ones (Gehan Breslow Wilcoxon test, p < 0.0001). (B) Representative images of spleens from WT and HMGA1P7 transgenic mice. (C) Spleens from HMGA1P7 (n = 12) transgenic mice were larger than spleens from WT (n = 4) (Mann-Whitney Test, **p < 0.0011). The error bars represent mean ± SD.

HMGA1P7 transgenic mice show a lymphoid malignancy (A) (I and II) Spleen from WT mouse showing normal morphology. (III) Representative image of immunoblastic lymphoma from a HMGA1P7-TG mouse. Expansion and merging of adjacent white pulp areas with loss of normal structures and germinal centers. (IV) A higher magnification shows a monotonous lymphoid population composed of fairly large rounded cells with scant cytoplasm and round to ovalar nuclei with finely dispersed chromatin and inconspicuous nucleoli. Haematoxylin and eosin (Original Magnification 4X for I and III panels, 40X for II and IV panels). (B) Representative images of liver (I), kidney (II) and lung (III) from a WT mouse with no pathological alteration. Monotonous lymphoid cells infiltrating the liver (IV), the kidney (V), the lung (VI) of HMGA1P7-TG mice. At higher magnification (inset), lymphoid population was composed of fairly large rounded cells with scant cytoplasm and round to ovalar nuclei with finely dispersed chromatin and inconspicuous nucleoli. Haematoxylin and eosin (Original Magnification 10X, inset 40X). (C) (I and II) immunohistochemistry of a WT mouse spleen showing normal distribution of CD3 immunolabelled T-cells and CD45/B220 immunolabelled B-cells. (III, IV) Immunohistochemical analysis of HMGA1P7-TG mouse spleen revealed a predominant CD45/B220 neoplastic lymphoid population with fewer scattered CD3-immunolabelled cells (Original Magnification 10X, inset 40X).
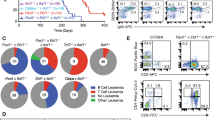
Furthermore, FACScan analysis of lymphocytes isolated from WT or pathological spleens using the CD3, CD19 and NK anti-mouse antibodies confirmed the immunohistochemical data. CD19 population resulted almost doubled, while CD3 population was decreased in HMGA1P7-TG mouse spleens in comparison with WT animals (Fig. 4A).

Mice overexpressing HMGA1P7 develop monoclonal expansion of the CD19 positive population. (A) FACScan analysis of splenic cells isolated from WT (n = 8) and HMGA1P7 (n = 14) transgenic mice using CD19, CD3, and NK1.1 anti-mouse antibodies. The results are reported as the mean of values. The error bars represent mean ± SD; *P < 0.05 **P < 0.01 (t test). (B) Genomic DNA isolated from the spleens of two WT mice and eight HMGA1P7-TG derived-lymphomas were amplified with DSF and JH4 primers. Three expected DJH bands with the spleen DNA were indicated. Within all eight tumor samples, DJH3 or DJH4 bands were predominantly detected.
To investigate the clonal status of the accumulation of the CD19 positive population in HMGA1P7-TG mice, genomic DNAs from TG and WT spleens were analysed. As shown in Fig. 4B, only one dominant PCR product was generated by the amplification of the DNA extracted from the transgenic spleens, whereas DNA derived from a WT spleen yielded three prominent PCR products of 1.0, 0.7 and 0.12 kb, corresponding to DJH2, DJH3 and DJH4 Immunoglobulin (Ig) gene rearrangements, respectively26.
Taken together, these results indicate that HMGA1P7-TG mice lymphoid expansion was monoclonal, therefore further supporting the diagnosis of B-cell lymphoma.
Identification of the genes modulated by HMGA1P7 expression in pathological spleens
Since HMGA1 did not result upregulated by HMGA1P7 overexpression in the analyzed pathological spleens and other mouse tissues (Fig. 5), we compared the transcriptome of spleens derived from HMGA1P7 transgenic mice (n = 2) versus that of WT spleens (n = 2) by RNA-Seq analyses, in order to better understand the mechanisms leading to lymphoid cell proliferation in transgenic mice. The upregulated transcripts included genes involved in inflammation (Ccl24, Il1a, Rgs16, Ccl5)27,28,29,30, in the NFKB pathway and in IL6/JAK/STAT3 and MTOR signalling, in oxidative phosphorylation (Uqcrc1, Ndufa1, Cox5a, Atp5d)31,32,33,34 and targets of MYC, E2F, STAT3, AP1, ATF3. In addition, the spleens of transgenic mice presented a gene expression signature compatible with an induction of senescence (Il13ra2, Il1a, Mmp3, Il1b)35,36,37,38 and immune escape (Pvrl2, Il10, Cd160, Ido1)39,40,41,42. Enrichment of genes downregulated by B cell receptor (BCR) inhibitors in diffuse large B cell lymphomas (DLBCL) was also unveiled by this analysis. Moreover, among the genes that showed a decreased expression in the transgenic spleens we found (i) transcripts down-regulated in post- germinal center (GC) BCL6 dependent B cell lymphomas when compared to MYC driven pre-GC lymphomas (Cnot6l, Sh3kbp1)43,44, (ii) genes repressed by BLIMP1 (Stat6, Zfp36l1)45,46; (iii) genes present in the GC B-cell type (GCB) DLBCL signature (Dtx1,Cux1, Sh3pxd2a, Klhl6)47,48,49 (Fig. 6).

HMGA1 is not upregulated in HMGA1P7-TG explanted-organs. qRT-PCR and Western Blot analyses of HMGA1 expression levels in WT (n = 3) and HMGA1P7 (n = 3) transgenic brain, liver, spleen, lung and kidney organs.

Transcriptome of HMGA1P7-TG derived-lymphomas. Representative GSEA plots illustrating the transcriptional expression signature enrichment in genes upregulated in transgenic spleens compared to normal spleens. Green line, enrichment score; bars in the middle portion of the plots show where the members of the gene set appear in the ranked list of genes. Positive or negative ranking metric indicate respectively correlation or inverse correlation with the profile. NES, normalised enrichment score. P, p-value. FDR, false discovery rate.
Then, we validated the results obtained by RNA-Seq analyses, testing the expression of a panel of deregulated mRNAs in spleens from HMGA1P7 by qRT-PCR (Fig. 7). Among the upregulated genes we chose CCAAT/enhancer-binding protein delta (Cebpd), chemokine (C-C motif) ligand 24 (Ccl24), Bcl-2-like 1 (Bcl2l1), Fos, Interleukin 1 Alpha (Il1a), BTB and CNC homolog 2 (Bach2), one of the downregulated genes. Next, the increased expression levels of Cebpd, Bcl2l1 and Fos were also confirmed by western blot analyses (Fig. 7). Finally, to demonstrate that HMGA1P7 acts through a ceRNA mechanism on the genes deregulated in pathological spleens (Fig. 8A), we inserted downstream of the luciferase open reading frame the 3′-UTRs of these genes. These reporter vectors were transfected into NIH3T3 cells overexpressing or not HMGA1P7. As expected, the luciferase activity was markedly increased in the cells that overexpressed HMGA1P7 (Fig. 8B), confirming the ceRNA action induced by HMGA1P7 on these new targets.

Validation of RNA-Seq analyses on HMGA1P7 spleens. qRT-PCR and Western Blot analyses of selected deregulated genes performed on WT (n = 4) and HMGA1P7 (n = 4) transgenic spleens. The results are reported as the mean of values. The error bars represent mean ± SD; *P < 0.05 (Mann-Whitney Test).

Deregulated genes from RNA-Seq are modulated by HMGA1P7 through a ceRNA mechanism. (A) 3′UTRs of Bcl2l1, Ccl24, Cebpd, Il1a and Fos share several microRNA Responsive Elements with HMGA1P7. (B) The Bcl2l1, Ccl24, Cebpd, Il1a and Fos 3′UTRs were cloned into the pMIR control vector and then transfected in NIH3T3 cells stably overexpressing the control vector or HMGA1P7. The results are reported as the mean of values. Error bars represent mean ± SD; n = 4. *P < 0.05 (Mann-Whitney Test).
Therefore, on the basis of the FACS and immunohistochemical data combined with the RNA-Seq analyses we can assess that the lymphoproliferation in the HMGA1P7 transgenic mice shares transcriptome features with DLBCL of the non-GCB type43,44,45,46,47,48,49.
Discussion
We have previously reported that the overexpression of HMGA1Ps accelerates cell proliferation, by enhancing the G1-S transition, increases cell migration ability, likely raising the levels of HMGA1 and other oncogenic proteins such as HMGA2 and EZH218. Moreover, the MEFs obtained from HMGA1Ps transgenic mice showed a reduced proliferation time and senescence in comparison with the WT MEFs18.
Therefore, the aim of this study was to better characterize the transgenic mice overexpressing the HMGA1P7 to possibly validate its oncogenic activity in vivo. The analysis of HMGA1P7 transgenic mice at 12 months of age, shows that about 50% of these mice developed a pathology characterized by splenomegaly and invasion of lymphoid cells in different anatomical districts. The pathological spleens showed a diffuse and monotonous lymphoid population effacing the splenic parenchyma with the loss of typical structures and germinal centres. Neoplastic lymphoid cells were medium to large, rounded, with scant cytoplasm and round to ovular nuclei with single or multiple prominent nucleoli.
By immunohistochemistry and FACS analyses, we found that the neoplastic cells were respectively positive for CD45/B220 and CD19 proposing a B cell phenotype of the lymphoid cells. Clonality assay on pathological spleens evidenced the clonal expansion of CD19-positive lymphoid population supporting a diagnosis of B cell lymphomas for these lesions. Interestingly, RNA-Seq analyses performed on spleens derived from WT and HMGA1P7 mice revealed a deregulation of several genes, likely due to HMGA1P7-ceRNA activity. The deregulated genes were involved in inflammation pathways such as NFKB pathway, the IL6/JAK/STAT3 and MTOR signalling, the oxidative phosphorylation, and targets of MYC, E2F, STAT3, AP1, ATF3. Moreover, the spleens from HMGA1P7 mice had a gene expression signature compatible with an induction of senescence and immune escape (Il13ra2, Il1a, Mmp3, Il1b, Pvrl2, Il10, Cd160, Ido1)35,36,37,38,39,40,41,42.
Noteworthily, the genes suppressed by BCR inhibitors in DLBCL were found significantly enriched in the pathological tissues of HMGA1P7 mice. In particular, the downregulated genes were enriched of transcripts decreased in post-GC BCL6 dependent B cell lymphomas and present in the GCB DLBCL signature. Therefore, the transcriptome study of the lymphoproliferative lesions in the HMGA1P7 transgenic mice unveils a pathology compatible with DLBCL of the non-GCB type.
Consistently with the ability of the HMGA1Ps to regulate gene transcription by a ceRNA mechanism, bioinformatic analyses demonstrate that several upregulated genes emerged from RNA-Seq data shared the same microRNA Responsive Elements with HMGA1P7 (i.e. Cebpd, Ccl24, Bcl2l1, Fos, Il1a).
Surprisingly, HMGA1 did not result upregulated by HMGA1P7 overexpression in the analysed pathological spleens, suggesting that pseudogene-induced lymphomas were based on other molecular targets already described27,28,29,30,31,32,33,34,35,36,37,38,39,40,41,42,43,44,45,46,47,48,49. Moreover, we did not find any change in HMGA1 expression levels during spleen development of HMGA1P7-transgenic mice (data not shown). However, we cannot exclude the possibility of increased HMGA1 protein levels in a limited cell compartment in the initial steps of lymphomagenesis.
Altogether the data presented here show that deregulated expression of HMGA1P7 pseudogene has oncogenic role also in vivo, thus representing a new class of genes involved in cancer pathology as their upregulation occurs frequently in multiple human cancers50. An oncogenic role for pseudogenes has been already reported. Indeed, mice engineered to overexpress the full-length murine B-Raf pseudogene Braf-rs1 develop an aggressive malignancy resembling human diffuse large B cell lymphoma by ceRNA mechanism that elevates BRAF expression50.
Noteworthy, preliminary studies on a mouse strain overexpressing HMGA1P6 pseudogene show that several mice develop a lymphoid pathology characterized by splenomegaly that resembles that found in HMGA1P7-TG mice.
Therefore, our mouse model confirms the oncogenic potential of pseudogenes and provides compelling support for a causal link between altered pseudogene expression and cancer, mediated by ceRNA mechanism. Studies are in progress to evaluate the expression levels of HMGA1 pseudogenes in human lymphomas. Preliminary results indicate HMGA1P1 overexpression that could contribute to lymphomagenesis by a similar ceRNA mechanism.
Materials and Methods
Transgenic mice
HMGA1P7 overexpressing mice have been previously generated and described18. Animals were housed in IRCCS “G. Pascale” animal facility as previously reported51 (project “Ruolo degli pseudogeni di HMGA1 nel cancro” Cod. 893/2013 approved by Italian Health Ministry on 13/05/2013). The experimental protocols were in complaints with the European Communities Council Directive (63/2010/EEC).
Cell culture and transfections
NIH3T3 cells were maintained in DMEM supplemented with 10% calf serum (Thermofisher, Waltham, MA, USA), glutamine and antibiotics. MycoAlert (Basel, Switzerland) was regularly used to test that cells were not infected with mycoplasma. Cell transfection protocol was reported elsewhere19.
Histology and immunohistochemistry
Light microscopy was performed as previously described52. Definition and classification of lymphoid disease were based on criteria reported elsewhere25.
For immunohistochemistry (IHC), 4-μm-thick sections were processed as previously described24,53. Primary antibodies included rabbit monoclonal to CD3 (ab16669, Abcam, Cambridge, UK) diluted 1: 200 and rabbit polyclonal to CD45/B220 as a marker for B cells54 (ab10558 Abcam, Cambridge, UK) diluted 1:1000.
RNA extraction and qRT-PCR
RNA extraction, RNA reverse transcription and Real-time PCR was performed as described elsewhere55. The following primers were used:
HMGA1P7-Fw 5′-gctccttctcggctcctc-3′HMGA1P7-Rev 5′-gcttgggcctcttttatgg-3′
Hmga1-Fw 5′-ggcagacccaagaaactgg-3′Hmga1-Rev 5′-ggcactgcgagtggtgat-3′
Cebpd-Fw λ5′-cttttaggtggttgccgaag-3′Cebpd-Rev 5′-ggcaacgaggaatcaagttt-3′
Ccl24-Fw 5′-gcagcatctgtcccaagg-3′Ccl24-Rev 5′-gcagcttggggtcagtaca-3′
G6pd-Fw 5′-cagcggcaactaaactcaga-3′G6pd-Rev 5′-ttccctcaggatcccacac-3′
Bcl2l1-Fw 5′-tgaccacctagagccttgga-3′Bcl2l1-Rev 5′-gctgcattgttcccgtaga-3′
Fos-Fw 5′-gggacagcctttcctactacc-3′Fos-Rev 5′-agatctgcgcaaaagtccg-3′
Il1a-Fw 5′-ttggttaaatgacctgcaaca-3′Il1a-Rev 5′-gagcgctcacgaacagttg-3′
Bach2-Fw 5′-gcagacagtgagtcgtgtcc-3′Bach2-Rev 5′-gttcctgggaaggtctgtga-3′
Flow cytometric analysis (FACS)
For FACS analyses, spleens were collected from WT and transgenic mice, hard-pressed through a stainless-steel mesh, resuspended in PBS and then in Red Blood Lysing Buffer (Sigma-Aldrich, Saint Louis, MI, US) for 3 min. After two washes in PBS, lymphocytes (5 × 105) were set in 96-well round-bottom dishes.
Monoclonal antibodies utilized: NK-FITC (clone # PK146), CD3 APC-H7 (clone # 560176), CD19 PE/cy7 (clone # HIB19). All antibodies were from BioLegend (San Diego, CA, US).
Analysis of the clonality of lymphomas
Genomic DNA was extracted from fresh spleens through Phenol/Chloroform/Isoamyl Alcohol Extraction (Thermofisher, Waltham, MA, USA). The obtained DNAs were utilized as PCR templates with DSF and JH4 primers that recognize mouse DNA DJ rearrangement26.
DSF primer: 5′-AGGGATCCTTGTGAAGGGATCTACTACTGTG-3′;
JH4 primer: 5′-AAAGACCTCCAGAGGCCATTCTTACC-3′.
RNA-Seq analyses
Genomix4life S.R.L. (Baronissi, Salerno, Italy) performed the next generation sequencing analysis, including samples quality control and Bioinformatics analysis. Following the producer’s guidelines, indexed libraries were obtained from 500 ng/ea RNA through TruSeq Stranded total RNA Sample Prep Kit (Illumina, San Diego, CA, USA). The libraries quantification was performed by the Agilent 2100 Bioanalyzer (Agilent Technologies, Santa Clara, CA, USA) and Qubit fluorometer (Thermofisher, Waltham, MA, USA), then combined in order that every index-tagged sample was in equimolar amounts, with 2 nM pooled samples final concentration. Illumina HiSeq. 2500 System (Illumina, San Diego, CA, USA) sequenced the pooled samples with a format of 2 × 100 paired-end at 8 pmol final concentration.
FastQC tool56 was utilized for the quality control analysis of the generated raw sequence files (.fastq files). Cutadapt was used in order to eliminate the adapter sequences. Paired-end reads were mapped using STAR (version 2.5.2b)57 on reference genome assembly mm10 acquired from Ensembl58,59. The quantification of transcripts expressed for each replicate of the sequenced samples was performed using HTSeq-Count algorithm60. The differential expression analysis was performed through DESeq. 261.
Gene Set Enrichment Analysis (GSEA) was used for functional annotation on pre-ranked lists using the MSigDB 5.262, the SignatureDB collection63 and genesets obtained from different publications64,65, applying false discovery rate (FDR) values <0.05 as threshold.
Luciferase assay
Dual-luciferase reporter assays were performed as previously described18.
Plasmids
For the mCebpd (NM_007679), mCcl24 (NM_019577), mBcl2l1 (NM_009743), mFos (NM_010234) and mIl1a (NM_010554) 3′UTR luciferase reporter constructs, the 3′UTR sequences were amplified by using the following primers:
mCebpd 3′utr-Fw 5′-gcagagctcagaattctgcctttctactaagatactggttg-3′
mCebpd 3′utr-Rv 5′-gcgatcgcttgaattcttagtgttctgggagctgcc-3′
mCcl24 3′utr-Fw 5′-gcagagctcagaattcccgcctctcctctgtccc-3′
mCcl24 3′utr-Rv 5′-gcgatcgcttgaattcacatcctggcagcaagagg-3′
mBcl2l1 3′utr-Fw 5′-gcagagctcagaattcgagcctctcgggaatgcttttc-3′
mBcl2l1 3′utr-Rv 5′-gcgatcgcttgaattccgcacagcaagccagcag-3′
mFos 3′utr-Fw 5′-gcagagctcagaattcgaatgttctgacattaacagttttc-3′
mFos 3′utr-Rv 5′-gcgatcgcttgaattcttcaacttaaatgcttttattgac-3′
mIl1a 3′utr-Fw 5′-gcagagctcagaattccaaaatgccagttgagtagga-3′
mIl1a 3′utr-Rv 5′-gcgatcgcttgaattcaggagactacatctaactgaccac-3′
The amplified fragments were cloned into pMirTarget vector (OriGene, Rockville, MD, USA) using In-Fusion HD Cloning kit (Takara Bio, Mountain View, CA, USA). HMGA1P7 overexpressing vector was previously described18.
Western blot
Western blot analyses were performed as previously described66. The primary antibodies used were: anti-Cebpd #7077 (ProSci, Poway, CA, USA); anti-Fos sc-166940 (Santa Cruz Biotechnology, Dallas, TX, USA); anti-Bcl2l1 #2762 (Cell Signaling, Danvers, MA, USA). Antibody against HMGA1 protein was described elsewhere67.
Statistical analysis
Two-sided unpaired Student’s t tests and Mann-Whitney tests were utilized to analyse data (GraphPad Prism, GraphPad Software, Inc.). P < 0.05 values were taking into account as statistically significant. The mean values +/− s.d were obtained from three or more separate experiments. GraphPad Prism, GraphPad Software, Inc. was used to obtain regression analyses and correlation coefficients.
Change history
11 June 2021
A Correction to this paper has been published: https://doi.org/10.1038/s41598-021-92173-4
References
Poliseno, L. Pseudogenes: newly discovered players in human cancer. Sci. Signal. 5, re5, https://doi.org/10.1126/scisignal.2002858 (2012).
Tam, O. H. et al. Pseudogene-derived small interfering RNAs regulate gene expression in mouse oocytes. Nature 453, 534–538, https://doi.org/10.1038/nature06904 (2008).
Watanabe, T. et al. Endogenous siRNAs from naturally formed dsRNAs regulate transcripts in mouse oocytes. Nature 453, 539–543, https://doi.org/10.1038/nature06908 (2008).
Hawkins, P. G. & Morris, K. V. Transcriptional regulation of Oct4 by a long non-coding RNA antisense to Oct4-pseudogene 5. Transcription 1, 165–175, https://doi.org/10.4161/trns.1.3.13332 (2010).
Johnsson, P., Lipovich, L., Grander, D. & Morris, K. V. Evolutionary conservation of long non-coding RNAs; sequence, structure, function. Biochim. Biophys. Acta 1840, 1063–1071, https://doi.org/10.1016/j.bbagen.2013.10.035 (2014).
Bier, A. et al. Connexin43 pseudogene in breast cancer cells offers a novel therapeutic target. Mol. Cancer Ther. 8, 786–793, https://doi.org/10.1158/1535-7163.MCT-08-0930 (2009).
Chiefari, E. et al. Pseudogene-mediated posttranscriptional silencing of HMGA1 can result in insulin resistance and type 2 diabetes. Nat. Commun. 1, 40, https://doi.org/10.1038/ncomms1040 (2010).
Han, Y. J., Ma, S. F., Yourek, G., Park, Y. D. & Garcia, J. G. A transcribed pseudogene of MYLK promotes cell proliferation. FASEB J. 25, 2305–2312, https://doi.org/10.1096/fj.10-177808 (2011).
Poliseno, L. et al. A coding-independent function of gene and pseudogene mRNAs regulates tumour biology. Nature 465, 1033–1038, https://doi.org/10.1038/nature09144 (2010).
Salmena, L., Poliseno, L., Tay, Y., Kats, L. & Pandolfi, P. P. A ceRNA hypothesis: the Rosetta Stone of a hidden RNA language? Cell 146, 353–358, https://doi.org/10.1016/j.cell.2011.07.014 (2011).
Fusco, A. & Fedele, M. Roles of HMGA proteins in cancer. Nat. Rev. Cancer 7, 899–910, https://doi.org/10.1038/nrc2271 (2007).
Sgarra, R. et al. High Mobility Group A (HMGA) proteins: Molecular instigators of breast cancer onset and progression. Biochim. Biophys. Acta Rev. Cancer 1869, 216–229, https://doi.org/10.1016/j.bbcan.2018.03.001 (2018).
Pegoraro, S. et al. A novel HMGA1-CCNE2-YAP axis regulates breast cancer aggressiveness. Oncotarget 6, 19087–19101, https://doi.org/10.18632/oncotarget.4236 (2015).
Pegoraro, S. et al. HMGA1 promotes metastatic processes in basal-like breast cancer regulating EMT and stemness. Oncotarget 4, 1293–1308, https://doi.org/10.18632/oncotarget.1136 (2013).
Gorbounov, M. et al. High mobility group A1 (HMGA1) protein and gene expression correlate with ER-negativity and poor outcomes in breast cancer. Breast Cancer Res. Treat. 179, 25–35, https://doi.org/10.1007/s10549-019-05419-1 (2020).
Roy, S. et al. HMGA1 overexpression correlates with relapse in childhood B-lineage acute lymphoblastic leukemia. Leuk. Lymphoma 54, 2565–2567, https://doi.org/10.3109/10428194.2013.782610 (2013).
Xu, Y. et al. The HMG-I oncogene causes highly penetrant, aggressive lymphoid malignancy in transgenic mice and is overexpressed in human leukemia. Cancer Res. 64, 3371–3375, https://doi.org/10.1158/0008-5472.CAN-04-0044 (2004).
Esposito, F. et al. HMGA1 pseudogenes as candidate proto-oncogenic competitive endogenous RNAs. Oncotarget 5, 8341–8354, https://doi.org/10.18632/oncotarget.2202 (2014).
Esposito, F., De Martino, M., Forzati, F. & Fusco, A. HMGA1-pseudogene overexpression contributes to cancer progression. Cell Cycle 13, 3636–3639, https://doi.org/10.4161/15384101.2014.974440 (2014).
Esposito, F. et al. HMGA1-pseudogene expression is induced in human pituitary tumors. Cell Cycle 14, 1471–1475, https://doi.org/10.1080/15384101.2015.1021520 (2015).
De Martino, M., Forzati, F., Arra, C., Fusco, A. & Esposito, F. HMGA1-pseudogenes and cancer. Oncotarget 7, 28724–28735, https://doi.org/10.18632/oncotarget.7427 (2016).
De Martino, M. et al. HMGA1P7-pseudogene regulates H19 and Igf2 expression by a competitive endogenous RNA mechanism. Sci. Rep. 6, 37622, https://doi.org/10.1038/srep37622 (2016).
De Martino, M. et al. The HMGA1 Pseudogene 7 Induces miR-483 and miR-675 Upregulation by Activating Egr1 through a ceRNA Mechanism. Genes (Basel) 8, https://doi.org/10.3390/genes8110330 (2017).
De Biase, D. et al. Characterization of inflammatory infiltrate of ulcerative dermatitis in C57BL/6NCrl-Tg(HMGA1P6)1Pg mice. Lab Anim, 23677218815718, https://doi.org/10.1177/0023677218815718 (2018).
Morse, H. C. III. et al. Bethesda proposals for classification of lymphoid neoplasms in mice. Blood 100, 246–258 (2002).
Chang, Y., Paige, C. J. & Wu, G. E. Enumeration and characterization of DJH structures in mouse fetal liver. EMBO J. 11, 1891–1899 (1992).
Menzies-Gow, A. et al. Eotaxin (CCL11) and eotaxin-2 (CCL24) induce recruitment of eosinophils, basophils, neutrophils, and macrophages as well as features of early- and late-phase allergic reactions following cutaneous injection in human atopic and nonatopic volunteers. J. Immunol. 169, 2712–2718 (2002).
Calvano, S. E. et al. A network-based analysis of systemic inflammation in humans. Nature 437, 1032–1037, https://doi.org/10.1038/nature03985 (2005).
Lippert, E. et al. Role of regulator of G protein signaling 16 in inflammation-induced T lymphocyte migration and activation. J. Immunol. 171, 1542–1555 (2003).
Medzhitov, R. & Horng, T. Transcriptional control of the inflammatory response. Nat. Rev. Immunol. 9, 692–703, https://doi.org/10.1038/nri2634 (2009).
Kunej, T. et al. Functional UQCRC1 polymorphisms affect promoter activity and body lipid accumulation. Obesity 15, 2896–2901, https://doi.org/10.1038/oby.2007.344 (2007).
Aguilera-Aguirre, L. et al. Mitochondrial dysfunction increases allergic airway inflammation. J. Immunol. 183, 5379–5387, https://doi.org/10.4049/jimmunol.0900228 (2009).
Gong, Y. Y. et al. Hypermethylation of Cox5a promoter is associated with mitochondrial dysfunction in skeletal muscle of high fat diet-induced insulin resistant rats. PLoS One 9, e113784, https://doi.org/10.1371/journal.pone.0113784 (2014).
Liu, H. et al. Identification of a new Mpl-interacting protein, Atp5d. Mol. Cell Biochem. 391, 85–94, https://doi.org/10.1007/s11010-014-1990-x (2014).
Sattiraju, A. et al. IL13RA2 targeted alpha particle therapy against glioblastomas. Oncotarget 8, 42997–43007, https://doi.org/10.18632/oncotarget.17792 (2017).
Laberge, R. M. et al. MTOR regulates the pro-tumorigenic senescence-associated secretory phenotype by promoting IL1A translation. Nat. Cell Biol. 17, 1049–1061, https://doi.org/10.1038/ncb3195 (2015).
Young, A. R. & Narita, M. SASP reflects senescence. EMBO Rep. 10, 228–230, https://doi.org/10.1038/embor.2009.22 (2009).
Salama, R., Sadaie, M., Hoare, M. & Narita, M. Cellular senescence and its effector programs. Genes. Dev. 28, 99–114, https://doi.org/10.1101/gad.235184.113 (2014).
Stamm, H. et al. Immune checkpoints PVR and PVRL2 are prognostic markers in AML and their blockade represents a new therapeutic option. Oncogene 37, 5269–5280, https://doi.org/10.1038/s41388-018-0288-y (2018).
Couper, K. N., Blount, D. G. & Riley, E. M. IL-10: the master regulator of immunity to infection. J. Immunol. 180, 5771–5777 (2008).
Cai, G. & Freeman, G. J. The CD160, BTLA, LIGHT/HVEM pathway: a bidirectional switch regulating T-cell activation. Immunol. Rev. 229, 244–258, https://doi.org/10.1111/j.1600-065X.2009.00783.x (2009).
Mbongue, J. C. et al. The Role of Indoleamine 2, 3-Dioxygenase in Immune Suppression and Autoimmunity. Vaccines 3, 703–729, https://doi.org/10.3390/vaccines3030703 (2015).
Luo, X., Shi, F., Qiu, H., Tong, Y. & Gao, X. Identification of potential key genes associated with diffuse large B-cell lymphoma based on microarray gene expression profiling. Neoplasma 64, 824–833, https://doi.org/10.4149/neo_2017_603 (2017).
Chen, J. et al. Dysregulated CXCR4 expression promotes lymphoma cell survival and independently predicts disease progression in germinal center B-cell-like diffuse large B-cell lymphoma. Oncotarget 6, 5597–5614, https://doi.org/10.18632/oncotarget.3343 (2015).
Wang, L. et al. Blimp-1 induced by IL-4 plays a critical role in suppressing IL-2 production in activated CD4 T cells. J. Immunol. 181, 5249–5256 (2008).
Nasir, A. et al. ZFP36L1 negatively regulates plasmacytoid differentiation of BCL1 cells by targeting BLIMP1 mRNA. PLoS One 7, e52187, https://doi.org/10.1371/journal.pone.0052187 (2012).
Gupta-Rossi, N. et al. Specific over-expression of deltex and a new Kelch-like protein in human germinal center B cells. Mol. Immunol. 39, 791–799 (2003).
McNerney, M. E. et al. CUX1 is a haploinsufficient tumor suppressor gene on chromosome 7 frequently inactivated in acute myeloid leukemia. Blood 121, 975–983, https://doi.org/10.1182/blood-2012-04-426965 (2013).
Xu-Monette, Z. Y. et al. Clinical and Biologic Significance of MYC Genetic Mutations in De Novo Diffuse Large B-cell Lymphoma. Clin. Cancer Res. 22, 3593–3605, https://doi.org/10.1158/1078-0432.CCR-15-2296 (2016).
Karreth, F. A. et al. The BRAF pseudogene functions as a competitive endogenous RNA and induces lymphoma in vivo. Cell 161, 319–332, https://doi.org/10.1016/j.cell.2015.02.043 (2015).
Fedele, M. et al. HMGA2 cooperates with either p27(kip1) deficiency or Cdk4(R24C) mutation in pituitary tumorigenesis. Cell Cycle 17, 580–588, https://doi.org/10.1080/15384101.2017.1403682 (2018).
Fedele, M. et al. Overexpression of the HMGA2 gene in transgenic mice leads to the onset of pituitary adenomas. Oncogene 21, 3190–3198, https://doi.org/10.1038/sj.onc.1205428 (2002).
Palumbo, A. Jr. et al. UBE2C is overexpressed in ESCC tissues and its abrogation attenuates the malignant phenotype of ESCC cell lines. Oncotarget 7, 65876–65887, https://doi.org/10.18632/oncotarget.11674 (2016).
Bleesing, J. J. & Fleisher, T. A. Human B cells express a CD45 isoform that is similar to murine B220 and is downregulated with acquisition of the memory B-cell marker CD27. Cytometry B Clin. Cytom. 51, 1–8, https://doi.org/10.1002/cyto.b.10007 (2003).
Forzati, F. et al. miR-155 is positively regulated by CBX7 in mouse embryonic fibroblasts and colon carcinomas, and targets the KRAS oncogene. BMC Cancer 17, 170, https://doi.org/10.1186/s12885-017-3158-z (2017).
Dobin, A. et al. STAR: ultrafast universal RNA-seq aligner. Bioinformatics 29, 15–21, https://doi.org/10.1093/bioinformatics/bts635 (2013).
Hubbard, T. et al. The Ensembl genome database project. Nucleic Acids Res. 30, 38–41 (2002).
Anders, S., Pyl, P. T. & Huber, W. HTSeq–a Python framework to work with high-throughput sequencing data. Bioinformatics 31, 166–169, https://doi.org/10.1093/bioinformatics/btu638 (2015).
Love, M. I., Huber, W. & Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq. 2. Genome Biol. 15, 550, https://doi.org/10.1186/s13059-014-0550-8 (2014).
Subramanian, A. et al. Gene set enrichment analysis: a knowledge-based approach for interpreting genome-wide expression profiles. Proc. Natl Acad. Sci. USA 102, 15545–15550, https://doi.org/10.1073/pnas.0506580102 (2005).
Shaffer, A. L. et al. A library of gene expression signatures to illuminate normal and pathological lymphoid biology. Immunol. Rev. 210, 67–85, https://doi.org/10.1111/j.0105-2896.2006.00373.x (2006).
Tarantelli, C. et al. PQR309 Is a Novel Dual PI3K/mTOR Inhibitor with Preclinical Antitumor Activity in Lymphomas as a Single Agent and in Combination Therapy. Clin. Cancer Res. 24, 120–129, https://doi.org/10.1158/1078-0432.CCR-17-1041 (2018).
Edgar, R., Domrachev, M. & Lash, A. E. Gene Expression Omnibus: NCBI gene expression and hybridization array data repository. Nucleic Acids Res. 30, 207–210 (2002).
Nicolau-Neto, P. et al. UBE2C Is a Transcriptional Target of the Cell Cycle Regulator FOXM1. Genes (Basel) 9, https://doi.org/10.3390/genes9040188 (2018).
Puca, F. et al. HMGA1 negatively regulates NUMB expression at transcriptional and post transcriptional level in glioblastoma stem cells. Cell Cycle 18, 1446–1457, https://doi.org/10.1080/15384101.2019.1618541 (2019).
Acknowledgements
This work was supported by grants from: PNR-CNR Aging Program 2012–2014, CNR Flagship Projects (Epigenomics-EPIGEN), Associazione Italiana per la Ricerca sul Cancro (AIRC IG 11477). M.D.M is recipient of a Fellowship from “Programma Valere Plus”, University of Campania “L. Vanvitelli”, Caserta, Italy. The Authors sincerely thank Mrs. Mariarosaria Montagna for her research support.
Author information
Authors and Affiliations
Contributions
M.D.M., F.F., A.F. and F.E. made substantial contributions to conception and design of the study, as well as analysis and interpretation of data and drafting of the article; S.C.C., D.D.B., O.P. and M.P. significantly contributed to the pathological evaluation of specimens and revision of the paper; M.D.M. realized FACS analyses; E.G. contributed to data interpretation; F.B. performed the bioinformatic analyses of the RNA-Seq data, giving substantial contributions to the interpretation of data and drafting of the article; G.P., C.A., A.B., G.D.V. and P.C. contributed to conception of the study and critically revised the paper.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
De Martino, M., De Biase, D., Forzati, F. et al. HMGA1-pseudogene7 transgenic mice develop B cell lymphomas. Sci Rep 10, 7057 (2020). https://doi.org/10.1038/s41598-020-62974-0
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-020-62974-0
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
