
Abstract
Extracellular matrix in solid tumors has emerged as a specific, stable, and abundant target for affinity-guided delivery of anticancer drugs. Here we describe the homing peptide that interacts with the C-isoform of Tenascin-C (TNC-C) upregulated in malignant tissues. TNC-C binding PL3 peptide (amino acid sequence: AGRGRLVR) was identified by in vitro biopanning on recombinant TNC-C. Besides TNC-C, PL3 interacts via its C-end Rule (CendR) motif with cell-and tissue penetration receptor neuropilin-1 (NRP-1). Functionalization of iron oxide nanoworms (NWs) and metallic silver nanoparticles (AgNPs) with PL3 peptide increased tropism of systemic nanoparticles towards glioblastoma (GBM) and prostate carcinoma xenograft lesions in nude mice (eight and five-fold respectively). Treatment of glioma-bearing mice with proapoptotic PL3-guided NWs improved the survival of the mice, whereas treatment with untargeted particles had no effect. PL3-coated nanoparticles were found to accumulate in TNC-C and NRP-1-positive areas in clinical tumor samples, suggesting a translational relevance. The systemic tumor-targeting properties and binding of PL3-NPs to the clinical tumor sections, suggest that the PL3 peptide may have applications as a targeting moiety for the selective delivery of imaging and therapeutic agents to solid tumors.
Similar content being viewed by others

Introduction
The microenvironment in solid tumors is shaped by concerted actions of tumor cells and cells in tumor stroma (endothelial cells, immune cells, and fibroblasts). All these cells deposit and remodel tumor extracellular matrix (ECM) – a complex meshwork of structural proteins and bioactive compounds. In addition to the structural role, the tumor ECM modulates invasion and distant spreading of tumor cells, tumor angiogenic status, and immune microenvironment, thereby plays a critical role(s) in tumor initiation, progression, and metastasis1. The molecular composition of tumor ECM reflects its type, origin, and physiological status2. Tumor ECM is rich in components expressed in developing and/or regenerating tissues that are typically absent or expressed at a lower level in quiescent healthy adult tissues and organs3. Tenascin C (TNC) is an ECM glycoprotein that is upregulated during normal tissue repair and in many human malignancies and plays important roles in tumor neovascularization, modulation of adhesion, and local and distant motility of malignant cells, and modulation of tumor immune status4. Several affinity ligands of TNC have been developed for targeting payloads to solid tumors, such as G11 single-chain antibody5, aptamers6, FH peptide7, and PL1 peptide8. TNC targeting antibodies are in phase I/II/III clinical trials as affinity targeting modules for cytotoxic or immunomodulatory payloads for therapy of solid tumors including metastatic renal cell carcinoma, metastatic melanoma, pancreatic carcinoma, acute myeloid leukemia, sarcoma and breast carcinoma, lung cancer, colorectal cancer, and glioma4. Importantly, malignant transformation is not only able to increase the expression of tenascin-C, but also influence its qualitative composition by modulating the splicing to result in different proportion of its isoforms. In particular, 91-amino acid alternatively spliced type-III homology domain of Tenascin-C FnIII C (TNC-C) has a tumor-restricted expression pattern5,8,9. The TNC-C is abundant in the glial tumors of the CNS, as well as in various carcinomas10,11,12. Overexpression of TNC-C correlates with tumor grade and is associated with poor prognosis12,13.
Peptides have several advantages over antibodies as tumor-targeting vehicles, including small size (and thus straightforward synthesis, eliminating the need for antibody-protein engineering), low immunogenicity, moderate cost, biocompatibility, and low-micromolar affinity range that circumvents the affinity site barrier14,15,16. Small peptides as targeting elements are particularly relevant in the context of nanomaterials, where even small molecule targeting ligands with weak affinity can through multivalent interactions significantly enhance target-specific avidity (by up to 4 orders of magnitude), and thus the affinity of the final material is readily tunable17.
Here we set out to develop peptidic affinity ligands for targeting TNC-C in solid tumors. Using peptide phage display, we identified an octameric homing peptide, PL3, that interacts with TNC-C in vitro and in vivo. In addition, PL3 peptide was able to interact with cell- and tissue penetration receptor NRP-1. Systemic PL3-guided nanoparticles accumulated in tumor xenografts implanted in mice. These nanoparticles were useful for tumor detection, imaging and served as a tumor-seeking carrier for a proapoptotic peptide anticancer payload. Our study suggests applications for PL3-targeted compounds and nanoparticles for improved detection and therapy of solid tumors.
Materials and Methods
Materials
Phosphate-buffered saline (PBS) was purchased from Lonza (Verviers, Belgium). K3[Fe(CN)6], HCl, isopropanol, Triton-X, Tween-20, CHCl3, MeOH, Isopropyl β-D-1-thiogalactopyranoside (IPTG) and dimethylformamide (DMF) were purchased from Sigma-Aldrich (Munich, Germany). Cys-fluorescein (FAM)-PL3 and Cys-FAM peptides with 6-aminohexanoic acid spacer were purchased from TAG Copenhagen (Denmark). PL3-D(KLAKLAK)2 and D(KLAKLAK)2 peptides were synthesized using Fmoc/t-Bu chemistry on a microwave-assisted automated peptide synthesizer (Liberty; CEM Corporation, Matthews, NC, USA). High-performance liquid chromatography (HPLC) was used to purify the peptides with 0.1% trifluoroacetic acid (TFA) in acetonitrile-water mixtures to 90%–95% purity. The peptides were validated by quadrupole time-of-flight (Q-TOF) mass spectral analysis18. CF647 amine dye was purchased from Biotium (Hayward, CA, USA).
Cell lines and experimental animals
The U87-MG (human glioblastoma, HTB-14) cells, PC3 (prostate carcinoma, CRL1435) cells, PPC1 (primary prostate carcinoma-1) cells were purchased from ATCC. Murine WT-GBM glioblastoma cells were a kind gift from Gabriele Bergers (UCSF, USA) and P3, P8, P13 stem cell-like, P22, NCH421K cells were a kind gift from Rolf Bjerkvig, (University of Bergen, Norway), and M21 melanoma cells were the gift of David Cheresh (USA). Cells and tumors were prepared as described previously19,20,21.
Athymic nude mice (Hsd/Athymic Fox1 nu Harlan) were purchased from Envigo (Netherlands) and maintained under standard housing conditions of the Animal Facility of the Institute of Biomedicine and Translational Medicine, University of Tartu (Tartu, Estonia). For orthotropic GBM tumor models, we used NCH421K, P13, and P3 stem cell-like, WT-GBM cells. The individual GBM cells around 2–3 × 105 in 3 μL PBS were intracranially implanted into mice brain 2 mm right and 1 mm anterior to the bregma. U87-MG and PC3 subcutaneous models were induced by injecting 2–9 × 106 cells in 100 µl PBS subcutaneously into the right flank of 11–15 week old male and female nude mice. All animal experiments were performed in accordance with the procedures approved by the Committee of Animal Experimentation of Estonian Ministry of Agriculture, permits #42 and #48.
Peptide phage biopanning
For biopanning on recombinant human TNC-C, we used NNK-encoded CX7C and X7 peptide T7 phage libraries with diversity ~5 × 108. Throughout our screens, the selected phages were amplified following plate amplification protocol22. The 1st and 4th rounds of biopanning were performed on TNC-C immobilized on Costar 96-Well ELISA plate (#3590, Corning Life Sciences, Tewksbury, MA, USA). Briefly, the multiwell plates were coated with 20 µg/ml recombinant TNC-C in 100 µl of PBS overnight at 4 °C, followed by blocking with 1% bovine serum albumin (BSA) in PBS overnight at 4 °C. The phage library (5 × 108 pfu in 100 µl of PBS-BSA) was incubated overnight at 4 °C, followed by 6 washes with PBS containing 1% BSA and 0.1% Tween-20 (washing buffer) to remove background phages, and by phage rescue and amplification in E. coli strain BLT5403 (Novagen, EMD Biosciences, MA, USA)8. The subsequent rounds of selection were performed on Ni-NTA Magnetic Agarose Beads (QIAGEN, Hilden, Germany) coated with His-6X tagged TNC-C (30 µg/10 µl beads) at room temperature for 1 h in 400 µl of PBS. The TNC-C beads were washed 3 times with washing buffer, followed by incubation with phages in (5 × 108 pfu in 100 µl in washing buffer) at room temperature for 1 h. The background phages were removed by rinsing 6 times with washing buffer, and the bound phages were eluted with 1 ml of PBS containing 500 mM Imidazole and 0.1% NP40.
The eluted phages were titered and amplified for a next round of selection. After 5 rounds of selection, peptide-encoding DNA from a set of 48 phage clones was subjected to Sanger sequencing of peptide-encoding phage DNA18,22. For cell-free binding studies with individual phage clones were incubated with Ni-NTA magnetic beads coated with hexahistidine-tagged TNC-C as above. RPARPAR phage binding to NRP-1-coated beads was used as a positive control23. Phage clones displaying heptaglycine peptide (GGGGGGG, G7), or insertless phage clones were used as negative controls.
Fluorescence polarization assay
Fluorescence anisotropy (FA) saturation binding experiments were set up as described previously24,25. The experiments were carried out in Dulbecco’s Phosphate Buffer Saline (Sigma-Aldrrich, Cat# D8662) with the addition of 0.1% Pluronic F-127 (Sigma-Aldrrich, Cat#P2443) in a final volume of 100 μl using 96‐well half area, flat‐bottom polystyrene NBS multiwell plates (Corning, Cat# 3686). The different concentrations of proteins (0–112 µM NRP1 or 0–275 µM TNC-C) were added to a fixed concentration (0.66 µM) of FAM-Cys-PL3 fluorescent ligand (KJ Ross-Petersen aps). The total and non‐specific binding was measured in the absence or in the presence of a 500 µM Biotin-Ahx-PL3 (KJ Ross-Petersen aps) respectively, after 24 h incubation at 25 °C in the dark, sealed with moisture barrier (4Titude, Cat# 4ti-0516/96). The concentration of fluorescent ligand and proteins in-stock solutions was determined by absorbance (for FAM-PL3 ε495 = 75000 M−1 cm−1, for NRP1 ε280 = 67630 M−1 cm−1 and TNC-C ε280 = 8480 M−1 cm−1 were used). The measurements were performed at 25 °C on a Synergy NEO (BioTek) microplate reader using an optical module with an excitation filter at 485 nm (slit 20 nm), emission filter at 528 nm (slit 20 nm) and polarizing beam splitting for dual-channel detection. Dual emission detection mode allows simultaneous recording of intensities that are parallel (I||) and perpendicular (I⊥) to the plane of excitation light. Sensitivities of channels (G factor) were calibrated with gain adjustment of the photomultiplier tubes using fluorescein (1 µM reference solution, Lambert Instruments) as a standard. The fluorescence anisotropy values were calculated as parameters FA from the equation X: FA = (I||−G·I⊥)/(I|| + 2·I⊥). The binding affinity was estimated by global fitting of the data as in25. This simultaneous fitting of total and non‐specific binding data takes into account the ligand depletion by both binding processes.
Nanoparticle synthesis and functionalization
The iron oxide nanoworms (NWs) were prepared according to a published protocol by8,26,27. The aminated NWs were PEGylated using maleimide-5K-PEG-NH. Peptides were coupled to NWs through a thioether bond between the thiol group of a cysteine residue added to the N-terminus of the peptide. The concentration of the NWs was determined by measuring the absorbance of NWs at 400 nm with a NanoDrop 2000c spectrophotometer (Thermo Scientific)8,27. Silver nanoparticles (AgNPs) were synthesized and functionalized as described28, CF647- N-hydroxysuccinimide-dye (NHS-dye) was conjugated to the PEG terminal amine groups, and biotinylated peptides were coated on the NeutrAvidin (NA) on the surface of the AgNPs. Transmission electron microscopy (TEM, Tecnai 10, Philips, Netherlands) was used to image the NPs and DLS (Zetasizer Nano ZS, Malvern Instruments, UK) was used to assess the zeta potential, polydispersity, and size of nanoparticles.
In vivo play-off phage auditioning
In vivo play-off was used for internally controlled and competitive systemic phage homing studies in mice bearing tumor xenografts. Phages displaying the candidate TNC-C binding peptides and control peptides were individually amplified and purified by precipitation with PEG-8000 (Sigma-Aldrich, St. Louis, MO, USA), followed by CsCl2 gradient ultracentrifugation and dialysis. The phages were pooled at equimolar representation (Table 1 for details) and injected iv at 1 × 1010 pfu (in 200 µl PBS) in tumor-bearing mice, circulated for 2 h, followed by anesthesia and intracardial perfusion with DMEM. The tumors and organs were collected in lysogeny broth (LB) medium containing 1% NP40, and the tissues were homogenized using a hand-held homogenizer. Phages in the tissue lysate were amplified in E.coli, purified by precipitation with PEG-8000, and DNA was extracted using a DNA extraction kit (High Pure PCR Template Preparation Kit; Roche, Basel, Switzerland). Next-generation sequencing of phage genomic DNA with Ion Torrent system (Thermo Fisher Scientific, Waltham, MA, USA) was used to evaluate the representation of each peptide-phage in the input mixture, in tumors and control organs. The FASTQ data from Ion Torrent were processed using a custom python script that identified the barcodes, constant flanking residues, and extracted the reads of the correct length.
In vivo fluorescence imaging with the IVIS Spectrum system
PL3 peptide tumor homing in mice bearing U87-MG xenografts was assessed by in vivo fluorescence imaging using the IVIS Spectrum imaging system (PerkinElmer, Waltham, MA). AgNPs coated with CF647 dye-labeled neutravidin were functionalized with biotinylated PL3 peptide (PL3-AgNP) or biotin (AgNP). AgNPs were iv injected, and the mice were placed in a dark chamber under isoflurane anesthesia for imaging (parameters: specific excitation filter, 650 nm; emission filter, 665 nm; auto exposure time; binning, medium; the field of view, 12; f/stop, 2). The images were captured at pre-injection (0 h) and 5 h post-injection. The signal was expressed in total Radiant Efficiency [p/s]/[µW/cm²]. The regions of interest (ROIs) were drawn for the whole tumor after tissue background correction, and radiant efficiency signal was quantitated. An automated spectral unmixing algorithm was used, images were analyzed by using Living Image 4.4 software (Caliper Life Sciences, Hopkinton, MA). Three animals per experimental group were analyzed. For the receptor blocking studies, PL3-AgNP injection was preceded by systemic pre-injection of blocking TNC-C and/or NRP1 antibodies (30 µg/mouse) 15 min prior to injection of the AgNPs.
Cell binding and internalization experiments
U87-MG, PPC1, and M21 cells cultured on glass coverslips were incubated with CF555-labeled AgNPs at 37 °C for 1 h, followed by washing with medium to remove background AgNPs. Etching solution freshly diluted from 0.2 M stock solutions of Na2S2O3 and K3Fe(CN)6 to 10 mM working concentration in PBS was applied to cells for 3 min, followed by PBS washes. The cells on coverslips were fixed with −20 °C methanol for 1 min. The cell membrane was visualized by staining with Alexa Fluor 488-labeled wheat germ agglutinin (WGA) at 1:1000 at RT for 1 h. Nuclei were stained with 4′,6-diamidino-2-phenylindole (DAPI, Molecular Probes) at 1 µg/mL. Fluoromount-G (Electron Microscopy Sciences) medium was used to mount the coverslips on microscope slides for confocal imaging.
Tumor-targeted delivery and biodistribution studies
FAM-labeled PL3 peptide coupled NWs or control FAM-NWs (7.5 mg /kg) in PBS were injected into the tail vein of mice bearing subcutaneous U87-MG, PC3, and orthotropic WT-GBM tumors. After 5 h circulation, the mice were perfused with 20 ml PBS/DMEM under deep anesthesia and tumors and control organs were collected. Tissues were ex vivo imaged using an Illuminatool Bright Light System LT-9900 (Lightools Research, Encinitas, CA, USA) and snap-frozen. The tissues were cryosectioned at 8–10 µm and mounted on a Superfrost plus slides. The sections were equilibrated at RT and fixed with 4% paraformaldehyde at RT for 20 min, or with methanol at −20 °C for 1 min. The immunostaining was performed with the following primary antibodies; rabbit anti-fluorescein IgG fragment (cat. no. A889, Thermo Fisher Scientific, MA, USA), rat anti-mouse CD31 (BD Biosciences, CA, USA), and in-house prepared CF647 or CF546-labeled single-chain ScFV G11. Secondary antibodies were Alexa 488 goat anti-rabbit IgG, Alexa 647 goat anti-rat IgG, and Alexa 546 goat anti-mouse IgG (all Invitrogen, CA, USA). Nuclei were counterstained with DAPI at 1 μg/ml. The coverslips were mounted on glass slides using Fluoromount-G (Electron Microscopy Sciences, PA, USA), imaged using confocal microscopy (Olympus FV1200MPE, Hamburg, Germany), and analyzed using the FV10 –ASW4.2 viewer, Imaris, and Fiji ImageJ software tools8,27.
Experimental therapy
Tumors were induced by s.c. injection of 8 ×106 U87-MG cells into the right dorsal flank of 18–20-week old male nude mice (weight 38 ± 5 g). When the tumor volume reached 100 ± 20 mm3, the mice were assigned randomly into 4 groups (N = 6 per group). For experimental therapy, the mice were treated with i.v. injections of 100 μl of PBS, FAM-D[KLAKLAK]2-NWs, FAM-PL3-NWs, and FAM-PL3-D[KLAKLAK]2-NWs every other day for 10 injections. Every 2 days the body weight and animal well-being (behavior, appearance, grooming) were assessed, and the tumor volume was measured using a digital caliper. The tumor volume was calculated with this formula: Volume (V) (mm3) = [length × (width)2]/2. The study was terminated when the tumor volume reached 2000 mm3 (or >20% body weight). At the end of the study, the mice were perfused, followed by tumors, and control organs were excised and snap-frozen for histological studies. Tumor volume, Kaplan–Meier survival and body weight curves were calculated for each group using the GraphPad Prism 6 software with p values <0.05 were considered significant.
Overlay assay on clinical tumor samples
All methods for the use of human samples were carried out in accordance with relevant guidelines and regulations. The collection and use of human samples was approved by the Research Ethics Committee of the University of Tartu, Estonia (permit #243/T-27). Freshly excised human samples were obtained during surgeries from Department of Neurosurgery, Tartu University Hospital, Estonia. The informed consent was obtained from all patients. The tissue samples were snap-frozen in liquid nitrogen, cryosectioned at 10 μm, fixed with methanol, and permeabilized with PBST buffer (1X Phosphate-Buffered Saline, 0.1% Tween 20), followed by incubation with blocking buffer containing 5% BSA, 5% goat serum, 5% FBS in PBST. For overlay, the sections were incubated with 20 μg/slide PL3-NW and NW overnight at 4 °C. The sections were washed and blocked with a blocking buffer, followed by immunostaining using rabbit anti-fluorescein primary antibodies and detection with the Alexa-488 anti-rabbit and mouse anti-TNC-C antibody and detection with the 647 goat anti-mouse IgG8.
Statistical analysis
Prism 6 software was used for statistical analysis. The results were presented as mean with error bars indicating ±SEM. For a comparison of 2 groups, we used a comparison using an unpaired t-test and multiple groups ANOVA test. P < 0.05 was considered significant and P-values were indicted as follows: *P ≤ 0.05, **P ≤ 0.01, ***P ≤ 0.001 and ****P ≤ 0.0001.
Results
Identification of a TNC-C binding peptide
TNC-C was expressed in E. coli and purified using affinity chromatography on Ni-NTA matrix (Fig. S1). For identification of TNC-C–interacting peptides, 5-round biopanning with of T7 CX7C peptide phage libraries was performed, using as the target his-tagged TNC-C coated on polystyrene multiwell plates in rounds 1 and 4, and TNC-C immobilized on Ni-NTA magnetic beads in rounds 2, 3 and 5 (Fig. S2). Selection on TNC-C in multiwell plates was included to avoid enrichment for histidine-containing peptides on the Ni-NTA beads. By round 5, ~1000-fold enrichment in the binding of the selected phage pool to TNC-C was observed (Fig. 1A). Sanger sequencing of a peptide-encoding region of the genome of 38 random phage clones from round 4 demonstrated a shift away from original cyclic CX7C library configuration due to frameshifts in the peptide-encoding region (Table. S1) and overrepresentation of the linear peptides containing the following motifs: RGRLXR (7 repeats), RGRLR (18 repeats), and RLXR (12 repeats)(Table S1). Next, we created a panel of T7 phage clones displaying peptides derived from the phage clone 5 (Table S1) that contains the longest enriched motif, RGRLXR, and characterized their binding to TNC-C (Fig. S3).

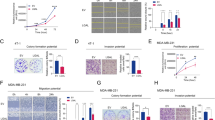
Identification and cell-free binding of PL3 peptide. (A) Selection of CX7C T7 phage library on immobilized TNC-C resulted in ~1000-fold increase in phage binding in round 5. Binding is expressed fold heptaglycine (G7) control phage. (B) Structural requirements for binding of PL3 peptide to TNC-C and NRP-1. T7-displayed PL3 peptide AGRGRLVR was subjected to alanine scanning mutagenesis (alanine substitutions indicated by underlined bold). Phage binding to TNC-C and NRP-1 is expressed as percent binding of parental PL3 phage. Values represent mean ± standard deviation from 3 independent experiments. (C,D) Saturation curve for the binding of FAM-Cys-PL3 to TNC-C and NRP1. FAM-Cys-PL3 (0.66 µM) was incubated with different concentrations of proteins in the absence (total binding, open circles) or presence (nonspecific binding, filled circles) of 0.5 mM Biotin-Ahx-PL3. After a 24 h incubation at 25 °C, FA values were calculated according to FA = (I | | − G·I⊥)/ (I | | + 2·I⊥) and fitted globally. The data represents the mean ± the standard deviation of 3 independent experiments.
For evaluation of systemic tumor homing of the candidate TNC-C-targeting peptides, we used in vivo phage play-off auditioning22. Tumor-bearing mice were injected i.v. with a phage pool of equally represented candidate TNC-C binding and control phages, followed by circulation, perfusion, and assessment of representation of phage clones in malignant tissue (4 glioma models and 1 prostate cancer model) and in normal brain tissue (Table 1). We found that a T7 clone displaying AGRGRLVR octapeptide was overrepresented in tumor tissue across models tested. In the following studies, we focused on AGRGRLVR peptide that we codenamed PL3.
Fluorescent anisotropy is widely used for solution-based characterization of interaction of small fluorescent ligands with larger partners and is thus well suited for studies of binding of FAM-Cys-PL3 (mW:1.46 kDa) to NRP1 b1b2 domain (mW:37.8 kD) and TNC-C (mW: 12.3 kDa). Binding of FAM-PL3 to both targets was saturable, although at different levels, likely due to different rotational mobility of bound ligand due to the ~3 fold difference in molecular weight of TNC-C and NRP-1. The Kd values were obtained by global fitting data to a binding isotherm, assuming single binding site with ligand depletion25. For PL3/NRP-1 interaction, the Kd was 1.1 ± 0.2 μM (Fig. 1D) - similar to published Kd values for the C-terminal arginine-displaying CendR peptide with the b1b2 domain of NRP-129. For PL3/TNC-C, the fluorescent anisotropy assay and calculations yielded Kd of 51 ± 19 μM (Fig. 1C). However, we observed dequenching of FAM signal after tryptic digestion of PL3/TNC-C complex (Fig. S17), suggesting involvement of more than one binding sites and thus, a lower Kd. The detailed mode of interaction of PL3 with TNC-C is a subject for follow-up studies.
PL3 peptide interacts with recombinant TNC-C and NRP-1 and is taken up by cultured tumor cells
We next studied the binding of alanine-substituted PL3-derivative peptide-phages to determine the amino acids important for peptide binding. The substitution of arginine or leucine residues in PL3 peptide resulted in decreased binding to recombinant TNC-C (Fig. 1B). In contrast, alanine substitutions of glycine and valine had a minimal effect. In contrast to robust interaction with TNC-C, the PL3 phage did not bind to fibronectin EDB-domain, a protein with similar size and negative surface charge as TNC-C (Fig. S4). PL3 peptide includes a C-terminal RXXR motif known to target NRP-1 cell and tissue penetration receptor23 overexpressed in most solid tumors. PL-3 phage bound to recombinant b1b2 domain of NRP-1 ~200 fold more than heptaglycine control phage (Fig. 1B). Interaction with NRP-1 was dependent on the presence of C-terminal arginine, as the phage with terminal R > A PL3-derivative peptide showed a dramatically reduced binding (Fig. 1B).
Many cultured tumor cell lines overexpress NRP-1 and internalize peptides with active CendR motif. We next studied the uptake of CF555-labeled PL3-AgNPs in TNC-C and NRP-1-positive U87-MG glioma cells, NRP-1-positive PPC1 prostate carcinoma cells, and in TNC-C and NRP-1-negative M21 melanoma cells (Fig. S15). The dye-labeled AgNP cores are well suited for fluorescence imaging as the AgNP increases the brightness of the surface-bound dye by about an order of magnitude by plasmonic enhancement. The AgNPs labeled with CF647 dye were coated with neutravidin (NA) and PEGylated as described previously, creating a stable colloid ready for coating with biotinylated peptides. The TEM and DLS showed that particles had an average size of 66.9 ± 27.6 nm, and zeta potential of −5.09 ± 0.19 mV in PBS (Fig. S5). After 1 h incubation with U87-MG and PPC1 cells, PL3-AgNPs were robustly endocytosed with perinuclear accumulation, whereas control nanoparticles showed only a background binding (Fig. 2A,B). In contrast, NRP-1-negative M21 cells showed a low background-like uptake with both PL3 and control AgNPs. Extracellular AgNPs can be removed by treatment with a mild biocompatible hexacyanoferrate/thiosulfate redox-based destain solution so that only internalized AgNP signal remains30. In U87-MG and PPC1 cells, etching resulted in a modest decrease of the cellular PL3-AgNP signal (Figs. 2B, S13), suggesting that most of the cell-associated particles were internalized and protected from etching by cellular membrane. The uptake of PL3-AgNPs in U87-MG cells was NRP-1-dependent, as incubation with function-blocking anti-NRP-1 antibody abolished PL3-AgNP signal in U87-MG cells treated with an etching solution to remove extracellular AgNPs (Fig. S14). These experiments showed that PL3 peptide interacts with TNC-C and NRP-1 and that PL3-functionalized nanoparticles are taken up by NRP-1-positive cultured cells.

PL3-functionalized AgNPs are taken up in NRP-1-positive PPC1 cells. PL3-AgNPs, or control AgNP particles (37 µL of 100 O.D. stock solution) labeled with CF555 fluorophore were incubated with PPC1 prostate carcinoma, U87-MG glioma, or M21 melanoma cells for 1 h, washed, treated with an optional hexacyanoferrate/thiosulfate redox-based etching solution to dissolve extracellular particles, and processed for confocal imaging. (A) Confocal imaging of cells incubated with peptide-targeted vs. control AgNPs. Note robust uptake of PL3-AgNPs (red) in NRP-1-positive PPC1 and U87-MG (and not NRP-1 negative M21) cells (high-magnification images are on the right of PL1-AgNP images). Control particles did not bind to the cells independent of their NRP-1 expression status. Scale bar: 20 µm (main images), and 2 µm (insets). (B) Quantitation of binding and internalization of CF555-labeled particles using ImageJ for samples from (A) and at least 3 independent experiments were carried out for individual conditions. Error bars: mean ± SD (N = 6), P-values were determined using unpaired Student’s t-test (ns, P > 0.05; ***p ≤ 0.001; ****p < 0.0001).
PL3-functionalized nanoparticles accumulate in malignant lesions
Systemic in vivo phage play-off studies showed that PL3 peptide-displaying phage outperforms other TNC-C binding candidate peptide phages in tumor homing. We next studied the effect of functionalization with synthetic PL3 peptide on in vivo tumor tropism of two classes of synthetic nanoparticles, iron oxide NWs and AgNPs.
We first tested the effect of PL3 functionalization on tumor homing of dextran-coated PEGylated paramagnetic NWs – a dual-use model nanoplatform that can be used as a carrier for drugs and for MR imaging due to intrinsic T2 contrast properties8,26. NWs with an average size of 88.8 ± 5 nm and zeta potential of −7.8 ± 2 mV (Fig. S6) were coated with FAM-labeled PL3 peptide or FAM-Cys control with no significant effect on particle size or surface charge. NWs were i.v. administered at 7.5 mg/kg NWs in mice bearing prostate cancer xenografts (PC3 s.c. tumors), or gliomas (s.c. U87-MG and orthotopic WT-GBM), both known to overexpress TNC-C and NRP-18,27. After 5 h circulation, accumulation of PL3-NWs, but not control NWs, was observed in all 3 tumor models tested (Fig. 3A–C). Confocal analysis demonstrated that whereas the PL3-NW homing was mainly overlapping or associated with CD31-positive vascular structures (Fig. 3A–C, arrows), in some regions, the PL3-NWs extravasated and accumulated in the tumor parenchyma (Fig. 3A–C, arrowheads). In control organs, PL3-NWs and control NWs showed similar background (Fig. S7). Macroscopic imaging confirmed preferential tumor accumulation of the PL3-NWs in U87-MG tumor mice (Fig. 4A,D). In tumor tissue PL3-NWs were found in the areas positive for TNC-C and NRP-1 immunoreactivities (Fig. 4B,C,E,F).

Systemic PL3-NWs accumulate in solid tumors. PL3-NWs or control FAM-NWs were injected i.v. at 7.5 mg/kg into mice bearing s.c. U87-MG glioblastoma (A), orthotopic WT-GBM glioblastoma (B), or s.c. PC3 prostate carcinoma (C). Five h later, the animals were intracardially perfused with DMEM/BSA and the tumors and control organs were snap-frozen, sectioned, immunostained with rabbit anti-FAM (green) and rat anti-CD31 (red) antibodies, counterstained with DAPI (blue) and imaged by confocal microscopy. Arrows point to PL3-NWs in the CD31-positive vessels and arrowheads point to extravasated PL3-NWs in the tumor parenchyma. Insets: confocal images without DAPI channel. Bar charts: quantitation of homing of NWs by quantitative analysis of FAM fluorescence in tissue sections by Fiji ImageJ. Error bars: mean ± SEM (N = 3–6). Scale bars: 100 μm. P-values were calculated using unpaired Student’s t-test (ns p > 0.05; *p < 0.05; **p ≤ 0.01; ***p ≤ 0.001).

PL3-NWs in tumor localize at areas positive for TNC-C and NRP1 immunoreactivities. PL3-NWs (A–C, upper row), or control NWs (D–F, lower row) were i.v.-administered at 7.5 mg/kg into mice bearing s.c. U87-MG glioblastoma. Five h later, the animals were intracardially perfused with DMEM, and tissues were collected for ex vivo macroscopic imaging and fluorescence microscopy. (A,D) Ex vivo macroscopic Illumatool imaging of PL3 and control NWs in green channel. Tu: tumor; Br: brain, Ki: kidney; Spl: spleen; Li: liver; Lu: lung; He: heart. The images are representative of 3 independent experiments. (B,C,E,F) Confocal imaging of NWs, TNC-C, and NRP-1 in tumor cryosections. PL3-NWs colocalize with perivascular TNC-C and NRP1 (arrows), whereas FAM-NWs show only background accumulation. Tumor tissues were stained with rabbit anti-FAM (green), anti-TNC-C ScFv G11 (red), and rabbit anti-NRP1 (red) antibodies. Arrows point to colocalization of PL3-NW with TNC-C and NRP1. Scale bar, 100 µm. The images are representative of 3 independent experiments.
We also studied the binding of PL3 and control NWs to the clinical glioma. The NWs were overlaid on cryosections of GBM, washed, and subjected to confocal imaging. PL3-NWs showed co-localization with TNC-C-positive structures in tumor perivascular space and parenchyma (Figs. S8,9). For TNC-C detection we used an in-house monoclonal antibody, and as a specificity
We also tested the effect of PL3 coating on tumor homing of near-infrared dye-labeled AgNPs. Intravital imaging with IVIS Spectrum after 5 h circulation showed that the PL3-CF647-AgNPs accumulated in U87-MG lesions at ~10 fold more than control CF647-AgNPs (Fig. 5A,B). Tumor homing of PL3-AgNPs was confirmed by confocal imaging (Fig. 5C). Coadministration of PL3-AgNPs with blocking rabbit polyclonal antibodies against either TNC-C or NRP-1 resulted in a decrease in tumor homing, and a cocktail of both antibodies almost completely inhibited the tumor accumulation (Fig. 5C,D). These data show that PL3 functionalization results in specific tumor homing of different classes of nanoparticles.

Systemic PL3-AgNPs home to U87-MG tumors. (A) In vivo imaging of U87-MG s.c. tumor-bearing mice injected with Alexa-647-labeled PL3-AgNPs (upper row), or with non-targeted AgNPs (lower row). The fluorescence imaging was performed using IVIS Lumina Imaging System at pre-injection and at 5 h post-injection. Three mice per group were i.v. injected with AgNPs; images after spectral un-mixing are shown. Note elevated tumor Alexa-647 signal in mice injected with PL3-AgNPs at 5 h post-injection. (B) Quantification of tumor Alexa-647AgNP fluorescence at pre-injection and at 5 h time points. The signal is expressed as Average Radiant Efficiency [p/s]/[µW/cm²]. Error bars: mean ± SEM (N = 3); P-values were determined using two-way ANOVA Fisher’s LSD test (ns, P > 0.05; ****P ≤ 0.0001). (C) The PL3-AgNP green signal was quantified from representative images using Fiji ImageJ. Error bars, mean ± SEM (N = 3 mice per group); scale bars: 20 μm; p-value was determined using Student unpaired t-test, two-tailed; ***p ≤ 0.001; ****p < 0.0001. (D) Effect of TNC-C and NRP-1 antibodies on tumor accumulation of PL3-AgNPs. PL3-AgNPs alone, or in combination with anti-TNC-C or anti-NRP1 antibodies, or a cocktail of both antibodies were i.v injected into mice bearing U87-MG xenograft tumors. Mice were perfused through the heart with PBS/DMEM 5 h after injection and organs were collected for cryosectioning and confocal microscopy. Colors: PL3-AgNPs (green), CD31-positive blood vessels (red) and DAPI (blue).control, we confirmed that preincubation of the antibody with recombinant TNC-C resulted in reduced staining (Fig. S10).
PL3-guided proapoptotic NWs have anti-glioma activity
We next tested the effect of PL3-functionalization on anticancer efficacy of nanoparticles loaded with proapoptotic D(KLAKLAK)2 peptide. For these studies we used s.c. U87-MG tumor model in order to monitor tumor size, rather than survival, as the endpoint. Starting on day 36 after tumor induction (when the tumors had reached 100 mm3), the tumor mice were treated with i.v. PL3-D(KLAKLAK)2-NWs, D(KLAKLAK)2-NWs, PL3-NWs, or PBS for 10 injections every other day, and tumor volume and survival of the mice were recorded (Fig. 6A). Over the treatment, the volume of tumors in PBS, PL3-NW, and D(KLAKLAK)2 –NW-treated mice increased rapidly. In contrast, tumor growth in the D(KLAKLAK)2 -PL3-treated group was significantly delayed (Fig. 6B). The median survival of PBS, D(KLAKLAK)2-NW, PL3-NW, and PL3-D(KLAKLAK)2-NW groups was 55, 58, 54, and 70 days, respectively. In the D(KLAKLAK)2 -PL3 group, 50% of mice had prolonged survival compared to the animals in the other treatment groups (Figs. 6C and S11). Immunostaining of post-treatment tumor tissue with anti-CD31 antibody to visualize tumor vasculature showed that compared to controls PL3-D(KLAKLAK)2-NW-treated tumors had a significant reduction in CD31-positive area and cleaved Caspase-3 showed a significant increase in apoptotic cells (Figs. 6D, S12, S15).

Experimental therapy with PL3-D(KLAKLAK)2-NWs suppresses glioblastoma growth. (A) Experimental design of the tumor treatment study. Treatment of mice bearing s.c. U87-MG tumors with systemic injections of NWs were initiated on day 36 when the tumors had reached 100 mm3 volume; groups of 6 randomized mice were treated every other day for 10 injections. The body weight, survival, behavior, and tumor volume were monitored every two days until the end of the treatment. (B) Tumor size dynamics of mice treated with 5 mg/kg of D(KLAKLAK)2-NW, PL3-NW and PL3-D(KLAKLAK)2-NW, or control PBS (N = 6 mice/group). The tumor volume was measured with a digital caliper and calculated using the formula: Volume (V) (mm3) = [length × (width × 2)]/2. Data were analyzed with 2-way ANOVA and log-rank test. Error bars: mean ± SEM; *P < 0.05. (C) Kaplan-Meier survival analysis. At the endpoint of the study the mice were sacrificed by perfusion, and organs and tumors were collected. Tumor volume, Kaplan–Meier survival curve and body weight curve were calculated for each group using the GraphPad Prism 6 software with p values <0.05 considered significant. (D) Treatment with PL3-D(KLAKLAK)2-NWs resulted in decreased tumor vascularization. Cryosections of tumors were stained with vascular endothelial marker CD31 to visualize tumor blood vessels using confocal microscopy. Red: blood vessels stained by the anti-CD31 antibody; blue: nuclei stained with DAPI. The images are representative of 3 independent tumors.
Discussion
Most affinity-based precision delivery strategies target receptors on the surface of tumor cells. This approach, while clearly proven useful, has limitations, as it relies on targeting a limiting number of systemically accessible receptors on genetically unstable malignant cells31. Compared to surface antigens on tumor cells, tumor-associated ECM represents an abundant and stable target with low shedding32. ECM directed affinity targeting strategies may allow simultaneous targeting of both malignant tumor cells and tumor support cells (fibroblasts, immune- and vascular cells) and be beneficial for improving treatment efficacies. TNC, an ECM component expressed at the invasive tumor front and in the angiomatrix, is predictive of adverse outcomes13 and provides specific targeting opportunities due to precisely controlled expression of its multiple structurally and functionally different isoforms4,11. Here, we report the development of an affinity ligand, octameric PL3 peptide, that targets C-domain of TNC and also interacts with NRP-1, a pleiotropic hub receptor upregulated in angiogenic sites and in malignant tissues involved in regulation of vascular permeability. Systemic PL3 phage nanoparticles and two types of synthetic PL3-guided nanocarriers home to solid tumors implanted in mice and bind to receptor-positive regions in clinical tumor samples. For therapeutic nanoparticles, PL3 functionalization improves their anticancer activity. Our study suggests potential uses for PL3 guided compounds and particles for improved detection and therapy of solid tumors.
Over the years, several preclinical and clinical-stage affinity ligands for TNC have been developed, such as a aptamers33, FHK peptide7, a bispecific PL1 peptide that in addition to TNC-C also targets fibronectin extra domain B8, TNC-C-targeting single-chain G11 antibody34, and TNC-C/D targeting monoclonal antibody 81C635. These and other ECM-reactive affinity ligands have proven useful for tumor delivery of extracellularly-acting anticancer payloads such as cytokines/growth factors, or payloads with intrinsic internalizing ability, such as proapoptotic D(KLKLAK)2 peptide nanoparticles, or cell-permeable cytotoxic compounds3,32,36. However, a challenge for these and other ECM-directed systemic compounds is that they can only reach extravascular tumor tissue passively, through the increased leakiness of aberrant tumor microvasculature, a phenomenon known as enhanced permeability and retention (EPR) effect37. EPR shows extensive inter- and intratumoral variability, thus decreasing the utility of the EPR-based targeting strategies38. During systemic in vivo play-off experiments using a panel of phage clones derived from screen on recombinant TNC-C in tumor mice, we observed that a phage clone displaying PL3 peptide with C-terminal RLVR motif outperformed other phage clones that had shown better TNC-C binding under cell-free conditions. C-terminal RLVR of PL3 corresponds to RXXR CendR consensus motif that, when exposed at the C-terminus, interacts with b1 domain of NRP-1 to trigger a trans-tissue pathway that mediates exit from the blood vessels and extravascular transport through tumor tissue23,39. Indeed, we observed specific binding of PL3 nanoparticles to recombinant NRP-1 b1b2 domain and not to b1b2 with mutated CendR binding pocket, and binding and internalization of the PL3 nanoparticles in NRP-1-positive PPC1 cells and no interaction with NRP-1-negative M21 melanoma cells. In vivo, NPs, including biological bacteriophage nanoparticles, are because of their size particularly prone to be excluded from difficult-to-access parts of tumors, and PL3 functionalization may mitigate this problem. PL3 peptide functionalization results in a small reproducible increase in uptake of NWs in the liver. The liver, along with other organs of the reticuloendothelial system, plays a central role in the nonselective clearance of NPs, primarily due to scavenger and phagocytic functions of liver Kupffer cells40,41. PL3-dependent increase in liver accumulation may be due to NRP-1 binding-ability of the peptide, as NRP1 has been detected in hepatic stellate cells and liver sinusoidal endothelial cells42,43. A potential strategy to mitigate NRP-1-dependent accumulation of PL3-targeted compounds in nontarget organs may be to render its CendR motif proteolytically activatable, as we have done with urokinase dependent uCendR3 peptide44. Numerous studies document the ability of CendR peptides to specifically increase the accumulation of a variety of anticancer therapeutics, such as chemotherapeutic agents, antibodies and NPs, in tumors45,46,47,48,49,50,51. For PL3 peptide, combination of CendR activity with the tumor ECM homing function may increase accessibility of TNC-C deeper in tumor parenchyma than would be possible under the conditions of physiological EPR with simple docking-based affinity targeting. In 2016, a chimeric 19-mer chimeric peptide composed of TNC-A-D domain binding peptide and tLyP-1 tumor penetrating peptide52 that targets NRP-1 on tumor cells was reported to allow anti-glioma drug delivery via NRP-1- and TNC-mediated specific penetration of nanoparticles into glioma parenchyma53. Compared to that peptide, the 8-amino acid PL3 peptide we have identified has the advantage of being smaller, and hence less likely to be immunogenic and simpler to produce and develop towards clinical applications. In both these peptides, the CendR motif is C-terminally exposed and does not need on-site proteolytic activation, as seen for several other tumor penetrating peptides. NRP-1 is expressed in the endothelial cells in non-malignant sites, albeit at lower levels than observed in tumors, and systemic peptides with C-terminally exposed CendR motif show increased background, in particular in first-met vascular beds, lung, and heart23,52. We have reported on development of tumor penetrating peptides containing cryptic CendR motifs that are cleavage-activated by a tumor-associated serine protease, urokinase-type plasminogen activator for tumor-specific extravasation and tissue penetration44,54. Our ongoing studies will explore homing and activation of PL3-derived cryptic CendR peptides with added C-terminal residues compatible with CendR motif activation of the peptide in tumor microenvironment. We hypothesize that this strategy may add additional layer of tumor selectivity to further improve the tumor homing/background ratio for PL3 and other ECM-targeted peptides.
The mode of interaction of PL3 peptide with TNC-C remains to be determined in follow-up studies. At neutral pH, the net charge of PL3 peptide is +3, whereas TNC-C is negatively charged (−5). Whereas electrostatic interactions may contribute to peptide-receptor interaction, the binding is likely to involve other mechanisms also, as no binding of PL3 phage was observed to Fn-EDB (charge −10 at pH 7). Ongoing in silico modeling combined with site-directed mutagenesis studies will map the binding site of PL3 on TNC-C. This information can be used to develop PL3 derivatives containing non-natural amino acids or low molecular weight peptidomimetic compounds of improved binding and plasma stability.
In summary, this study describes the identification of an 8-amino acid homing peptide, PL3, that interacts with TNC-C and with the cell- and tissue penetration receptor NRP-1. Systemic PL3-guided nanoparticles accumulated in tumor xenografts implanted in mice. The PL3-guided nanoparticles were useful for tumor detection, imaging and served as a tumor-seeking carrier for a proapoptotic peptide anticancer payload. Our study warrants future studies to study applications for PL3-targeted compounds and nanoparticles for improved detection and therapy of solid tumors.
References
Pickup, M. W., Mouw, J. K. & Weaver, V. M. The extracellular matrix modulates the hallmarks of cancer. EMBO Rep. 15, 1243–1253 (2014).
Lu, P., Weaver, V. M. & Werb, Z. The extracellular matrix: A dynamic niche in cancer progression. J. Cell Biol. 196, 395–406 (2012).
Venning, F. A., Wullkopf, L. & Erler, J. T. Targeting ECM Disrupts Cancer Progression. Front. Oncol. 5, 224 (2015).
Spenlé, C. et al. Tenascin-C: Exploitation and collateral damage in cancer management. Cell Adh. Migr. 9, 141–53 (2015).
Silacci, M. et al. Human monoclonal antibodies to domain C of tenascin-C selectively target solid tumors in vivo. Protein Eng. Des. Sel. 19, 471–478 (2006).
Daniels, D. A., Chen, H., Hicke, B. J., Swiderek, K. M. & Gold, L. A tenascin-C aptamer identified by tumor cell SELEX: systematic evolution of ligands by exponential enrichment. Proc. Natl. Acad. Sci. USA 100, 15416–15421 (2003).
Kim, M. Y. et al. Selection and characterization of tenascin C targeting peptide. Mol. Cells 33, 71–77 (2012).
Lingasamy, P. et al. Bi-specific tenascin-C and fibronectin targeted peptide for solid tumor delivery. Biomaterials 219, 119373 (2019).
Schliemann, C. & Neri, D. Antibody-based targeting of the tumor vasculature. Biochim. Biophys. Acta 1776, 175–92 (2007).
Leins, A. et al. Expression of tenascin-C in various human brain tumors and its relevance for survival in patients with astrocytoma. Cancer 98, 2430–2439 (2003).
Brösicke, N., Van Landeghem, F. K. H., Scheffler, B. & Faissner, A. Tenascin-C is expressed by human glioma in vivo and shows a strong association with tumor blood vessels. Cell Tissue Res. 354, 409–430 (2013).
Carnemolla, B. et al. Identification of a glioblastoma-associated tenascin-C isoform by a high affinity recombinant antibody. Am. J. Pathol. 154, 1345–52 (1999).
Orend, G., Saupe, F. & Schwenzer, A. The extracellular matrix and cancer: regulation of tumor cell biology by tenascin-C. (2012).
Zhao, N., Qin, Y., Liu, H. & Cheng, Z. Tumor-Targeting Peptides: Ligands for Molecular Imaging and Therapy. Anticancer. Agents Med. Chem. 18, 74–86 (2018).
Fosgerau, K. & Hoffmann, T. Peptide therapeutics: Current status and future directions. Drug Discov. Today 20, 122–128 (2015).
Albericio, F. & Kruger, H. G. Therapeutic peptides revisited. Future Med. Chem. 4, 1527–1531 (2012).
Ruoslahti, E. Peptides as targeting elements and tissue penetration devices for nanoparticles. Adv. Mater. 24, 3747–3756 (2012).
Ikemoto, H. et al. Hyaluronan-binding peptide for targeting peritoneal carcinomatosis. Tumor Biol. 39, 1010428317701628 (2017).
Bougnaud, S. et al. Molecular crosstalk between tumour and brain parenchyma instructs histopathological features in glioblastoma. Oncotarget 5, (2016).
Talasila, K. M. et al. EGFR wild-type amplification and activation promote invasion and development of glioblastoma independent of angiogenesis. Acta Neuropathol. 125, 683–98 (2013).
Keunen, O. et al. Anti-VEGF treatment reduces blood supply and increases tumor cell invasion in glioblastoma. Proc. Natl. Acad. Sci. USA 108, 3749–54 (2011).
Teesalu, T., Sugahara, K. N. & Ruoslahti, E. Mapping of vascular ZIP codes by phage display. Methods Enzymol. 503, 35–56 (2012).
Teesalu, T., Sugahara, K. N., Kotamraju, V. R. & Ruoslahti, E. C-end rule peptides mediate neuropilin-1-dependent cell, vascular, and tissue penetration. Proc. Natl. Acad. Sci. USA 106, 16157–62 (2009).
Veiksina, S. et al. Homogeneous fluorescence anisotropy-based assay for characterization of ligand binding dynamics to GPCRs in budded baculoviruses: The case of Cy3B-NDP-a-MSH binding to MC4 receptors. In G Protein-Coupled Receptor Screening Assays: Methods and Protocols 37–50, https://doi.org/10.1007/978-1-4939-2336-6_3 (Springer New York, 2015).
Veiksina, S., Kopanchuk, S. & Rinken, A. Budded baculoviruses as a tool for a homogeneous fluorescence anisotropy-based assay of ligand binding to G protein-coupled receptors: The case of melanocortin 4 receptors. Biochim. Biophys. Acta - Biomembr. 1838, 372–381 (2014).
Park, J.-H. et al. Magnetic Iron Oxide Nanoworms for Tumor Targeting and Imaging. Adv. Mater. 20, 1630–1635 (2008).
Säälik, P. et al. Peptide-guided nanoparticles for glioblastoma targeting. J. Control. Release 308, 109–118 (2019).
Willmore, A.-M. A. et al. Targeted silver nanoparticles for ratiometric cell phenotyping. Nanoscale 8, 9096–9101 (2016).
Wang, J. et al. Selection of phage-displayed peptides on live adherent cells in microfluidic channels. Proc. Natl. Acad. Sci. USA 108, 6909–6914 (2011).
Braun, G. B. et al. Etchable plasmonic nanoparticle probes to image and quantify cellular internalization. Nat. Mater. 13, 904–11 (2014).
Hussain, S., Rodriguez-Fernandez, M., Braun, G. B., Doyle, F. J. & Ruoslahti, E. Quantity and accessibility for specific targeting of receptors in tumours. Sci. Rep. 4, 5232 (2014).
Raavé, R., van Kuppevelt, T. H. & Daamen, W. F. Chemotherapeutic drug delivery by tumoral extracellular matrix targeting. J. Control. Release 274, 1–8 (2018).
Hicke, B. J. et al. Tenascin-C aptamers are generated using tumor cells and purified protein. J. Biol. Chem. 276, 48644–54 (2001).
Mårlind, J. et al. Antibody-mediated delivery of interleukin-2 to the stroma of breast cancer strongly enhances the potency of chemotherapy. Clin. Cancer Res., https://doi.org/10.1158/1078-0432.CCR-07-5041 (2008).
Lee, Y. S. et al. Therapeutic Efficacy of Antiglioma Mesenchymal Extracellular Matrix 131I-Radiolabeled Murine Monoclonal Antibody in a Human Glioma Xenograft Model. Cancer Res. 48, 584–588 (1988).
Fernandes, C., Suares, D. & Yergeri, M. C. Tumor Microenvironment Targeted Nanotherapy. Front. Pharmacol. 9, 1230 (2018).
Maeda, H. Macromolecular therapeutics in cancer treatment: The EPR effect and beyond. Journal of Controlled Release 164, 138–144 (2012).
Prabhakar, U. et al. Challenges and key considerations of the enhanced permeability and retention effect for nanomedicine drug delivery in oncology. Cancer Res. 73, 2412–7 (2013).
Teesalu, T., Sugahara, K. N. & Ruoslahti, E. Tumor-Penetrating Peptides. Front. Oncol. 3, 216 (2013).
Longmire, M., Choyke, P. L. & Kobayashi, H. Clearance properties of nano-sized particles and molecules as imaging agents: Considerations and caveats. Nanomedicine 3, 703–717 (2008).
Tsoi, K. M. et al. Mechanism of hard nanomaterial clearance by the liver. Nat. Mater. 15, 1212–1221 (2016).
Zhuang, P.-Y. et al. Peritumoral Neuropilin-1 and VEGF receptor-2 expression increases time to recurrence in hepatocellular carcinoma patients undergoing curative hepatectomy. Oncotarget 5, (2014).
Bergé, M. et al. Neuropilin-1 is upregulated in hepatocellular carcinoma and contributes to tumour growth and vascular remodelling. J. Hepatol. 55, 866–875 (2011).
Braun, G. B. et al. Urokinase-controlled tumor penetrating peptide. J. Control. Release 232, 188–195 (2016).
Sugahara, K. N. et al. Tissue-Penetrating Delivery of Compounds and Nanoparticles into Tumors. Cancer Cell 16, 510–520 (2009).
Sugahara, K. N. et al. Coadministration of a tumor-penetrating peptide enhances the efficacy of cancer drugs. Science 328, 1031–5 (2010).
Gu, G. et al. The influence of the penetrating peptide iRGD on the effect of paclitaxel-loaded MT1-AF7p-conjugated nanoparticles on glioma cells. Biomaterials 34, 5138–5148 (2013).
Akashi, Y. et al. Anticancer effects of gemcitabine are enhanced by co-administered iRGD peptide in murine pancreatic cancer models that overexpressed neuropilin-1. Br. J. Cancer 110, 1481–1487 (2014).
Sha, H. et al. Tumor-penetrating peptide fused EGFR single-domain antibody enhances cancer drug penetration into 3D multicellular spheroids and facilitates effective gastric cancer therapy. J. Control. Release 200, 188–200 (2015).
Wang, K. et al. Tumor penetrability and anti-angiogenesis using iRGD-mediated delivery of doxorubicin-polymer conjugates. Biomaterials 35, 8735–8747 (2014).
Schmithals, C. et al. Improving drug penetrability with iRGD leverages the therapeutic response to sorafenib and doxorubicin in hepatocellular carcinoma. Cancer Res. 75, 3147–3154 (2015).
Roth, L. et al. Transtumoral targeting enabled by a novel neuropilin-binding peptide. Oncogene 31, 3754–3763 (2012).
Kang, T. et al. Synergistic targeting tenascin C and neuropilin-1 for specific penetration of nanoparticles for anti-glioblastoma treatment. Biomaterials 101, 60–75 (2016).
Sharma, S. et al. Tumor-Penetrating Nanosystem Strongly Suppresses Breast Tumor Growth. Nano Lett. 17, 1356–1364 (2017).
Acknowledgements
We thank Gabriele Bergers for WT GBM cells, Erkki Juronen for help with ÄKTA Protein Purification System, Rein Laiverik for help with the characterization of the NWs. This work was supported by the European Regional Development Fund (Project No. 2014–2020.4.01.15–0012), by EMBO Installation grant #2344 (to T. Teesalu), European Research Council grants GBMDDS and GLIOGUIDE from European Regional Development Fund (to T. Teesalu), Wellcome Trust International Fellowship WT095077MA (to T. Teesalu), and Norwegian-Estonian collaboration grant EMP181 (to T. Teesalu). We also acknowledge the support of Estonian Research Council (grant PRG230 and EAG79 to T. Teesalu) and Estonian Research Council grants (IUT20‐17 and PSG230 to S. Kopanchuk).
Author information
Authors and Affiliations
Contributions
Prakash Lingasamy – coordinated the study, performed the experiments, drafted the manuscript. Allan Tobi – prepared and characterized the nanoparticles. Kaarel Kurm – performed ion-torrent next-generation sequencing. Sergei Kopanchuk – performed and interpreted the fluorescence polarization assay. Aleksander Sudakov – wrote Python script for next generation sequencing data analysis. Markko Salumäe – participated in phage biopanning. Tõnu Rätsep – provided access to clinical samples and data. Toomas Asser – provided access to clinical samples and data. Rolf Bjerkvig – provided support in GBM modeling, participated in data interpretation. Tambet Teesalu – concepted and supervised the study, wrote the manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supporting Information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Lingasamy, P., Tobi, A., Kurm, K. et al. Tumor-penetrating peptide for systemic targeting of Tenascin-C. Sci Rep 10, 5809 (2020). https://doi.org/10.1038/s41598-020-62760-y
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-020-62760-y
This article is cited by
-
Magnetic transferrin nanoparticles (MTNs) assay as a novel isolation approach for exosomal biomarkers in neurological diseases
Biomaterials Research (2023)
-
Tumor Homing Peptides as Fusion Partners of Therapeutic Proteins for Efficient Delivery to Cancer Cells
Biotechnology and Bioprocess Engineering (2023)
-
The matrix in cancer
Nature Reviews Cancer (2021)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
