
Abstract
Acoustic communication in the form of courtship and mating songs are often involved in reproductive isolation between species of Diptera, such as Drosophila, mosquitoes and sand flies. The patterns of courtship songs in New World sand fly species evolve quickly under sexual selection; and therefore, represent an important trait that can be used as a marker to study the evolution of species complexes and may aid identification of sibling species with a complex. The ability to identify vector species within species complexes is of critical importance for effective and efficient vector control programs. Species-specific song patterns seems to contribute to reproductive isolation in New World sand fly species, suggesting that auditory communication signals may be widespread among these important vectors of leishmaniasis. The main goal of the present study was to characterize the copulatory courtship song of Phlebotomus argentipes, an important vector of visceral leishmaniasis in the Old World. Ph. argentipes males produce acoustic signals during copulation and two types of songs were observed. The one we called primary song is a ‘pulse song’ with similar length and amplitude to the previously observed ‘P1’ pattern recorded in Brazilian populations of Lu. longipalpis s.l. The secondary song has ‘sine song’ characteristics and is quite different from any song produced by New World species. The discovery of this copulation courtship songs in Ph. argentipes supports the possibility that acoustic communication in sandflies might be more widespread than previously thought, including Old World species. Our results highlight the importance of further research on acoustic communication in the Ph. argentipes species complex and other Old World vectors of leishmaniasis.
Similar content being viewed by others

Introduction
Acoustic signaling represents one of several methods of insect communication and can be used as a defense mechanism in male-male competition and for male-female intra-specific recognition1,2. When associated with mating behavior, songs are frequently under sexual selection and thus can diverge quickly3,4,5. In Drosophila species, differences in acoustic signals are often associated with pre-mating reproductive isolation and represent sexual traits that result in restricted gene flow between closely related species6. Moreover, acoustic communication studies have played a key role in the identification of cryptic sibling species, and therefore, can provide species-specific traits for taxonomic studies when song are associated with reproductive success7,8.
Lutzomyia longipalpis s.l. Lutz & Neiva 1912 is known to consist of a number of cryptic species that are morphologically indistinguishable from each other9. Males of this species produce acoustic signals by flapping their wings. Usually acoustic signals associated with reproductive behavior are produced during pre-mating courtship, as in most Drosophila species, and these signals are important for reproductive success1. Unlike Drosophila, males of Lu. longipalpis s.l produce songs after copulation has started, e.g. once the male genital clasps the female genitalia8,10,11,12. Although not very common, copulatory courtship has been reported in some insect groups13,14,15,16,17. In the case of Lu. longipalpis s.l, copulatory courtship songs are likely to be involved in insemination success as many mating experiments have demonstrated high levels of insemination failure following copulation between sibling species that produce different songs8,10,11,12.
Two types of song have been observed in the Lu. longipalpis species complex: a “primary” song which is a pulse song produced by all males during copulation and a “secondary” song composed of low amplitude polycyclic pulses with variable intervals that are produced by some (but not all) males. The primary copulatory courtship song varies considerably among Brazilian populations of Lu. longipalpis s.l with three patterns already identified: Burst-type, Pulse-type and Mix-type18. The song variation observed suggests that the Lu. longipalpis species complex in Brazil consists of at least six cryptic species19.
Males from other New World sandfly species also produce acoustic signals. The main vector of visceral leishmaniasis (VL) in the Central-West region of Brazil is Lu. cruzi Mangabeira 1938. In this species, which belongs to the Lu. longipalpis species complex20, males produce a Burst-type copulation song similar to the Burst-type songs produced by some of the Lu. longipalpis s.l. sibling species19.
Copulation songs have also been found in sandfly vectors of American cutaneous leishmaniasis (ACL). Males of Lutzomyia migonei França 1920 produce short ‘trains’ which consist of a small number of pulses with each train exhibiting a short inter-pulse interval. This pattern is different from the Pulse songs observed in the Lu. longipalpis species complex21. Moreover, pulse songs consisting of short trains were also recorded in males of Lu. (Nyssomyia) intermedia Lutz & Neiva 1912, one of the main vectors of ACL. Unlike the species mentioned previously, Lu. intermedia males produce pre-copulatory courtship acoustic signals similar to those observed in several species of Drosophila22,23. These observations suggest that acoustic communication is widespread in New World sand flies.
In South East Asia, Phlebotomus (Euphlebotomus) argentipes (Diptera: Culicidae) Annandale & Brunette 1908, is the vector of the Protist Leishmania donovani (Kinetoplastida: Trypanosomatidae)24. It is thought that member from the Phlebotomus and Lutzomyia genera have separated approximately 200 MYA25. Both the Old World Ph. argentipes and the New World Lu. longipalpis species display lekking and pheromone communication behavior as part of their mating behavioral repertoire and males have been observed in the field displaying wing fanning26,27,28,29. Our aim, therefore, was to determine if acoustic signaling may also take place in the Old World leishmaniasis vector, Ph. argentipes.
Results and Discussion
It is known that Ph. argentipes males, like Lu. longipalpis s.l. males, do not produce a courtship song similar to those that have been observed in Drosophila species22,30. Lutzomyia intermedia is the only sand fly species that is known to produce a song during courtship.
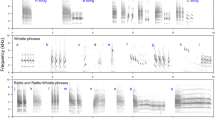
The study presented here shows that Ph. argentipes males produce acoustic signals during copulation, which have similarities to the Lu. longipalpis s.l., Lu. migonei and Lu. cruzi copulation songs19,20,21,31. Only males were observed to produce songs. They produced two types of song: a primary song which is a pulse song, and a secondary song which has sine song characteristics (Fig. 1). Pulse songs consist of trains of uni- or polycyclic sound pulses and sine songs are continuous, humming-like sounds32. Figure 2 shows the song spectrograms of both types of songs (audio file:Additional File 1).

Copulatory courtship song of Ph. argentipes. (A) General view of song trace showing primary followed by a secondary song trace, (B) Trace of a pulse-type primary song and (C) Trace of a sine song of the secondary song. Figures (B,C) show 1 second of song.

Song spectrogram of copulatory courtship song of Ph. argentipes. Pulses with similar length and amplitude compose the primary song and multiple harmonics with five different frequencies are evident in the secondary song.
The primary song is composed of pulses with similar length and amplitude and shares the qualitative properties of the Pulse-type song observed in some Brazilian populations of Lu. longipalpis s.l, e.g. the P1 pattern recorded previously in populations from Jacobina, Jequié and Cavunge (Bahia State). Each train is composed of 43 to 83 pulses (mean 58 ± 3.6), with a mean inter-pulse interval (IPI) of 54.9 ms (SEM ± 4.21), a length of 3.1 s (SEM ± 0.29) and a mean of frequency of 246.8 Hz (SEM ± 15.44) (Additional File 2; Fig. 1B).
The secondary song follows immediately after the primary song and has a mean frequency of 313.1 Hz (SEM ± 7.36) and lasts approximately 2.3 s (SEM ± 0.21) presenting multiple harmonics that resemble the flight sound observed in several mosquito species33,34 (Additional File 2; Fig. 1C; Additional File 3). The Ph. argentipes secondary song presents more differences when compared to the New World species counterpart. Some male Lu. longipalpis s.l. also produce a secondary song18,31, however, the pattern is quite different to the one observed in Ph. argentipes. The Lu. longipalpis s.l. secondary song is more of a pulse-like song with polycyclic pulses that are flanked by two primary songs, and it is not produced by every male. On the other hand, the secondary song of Ph. argentipes was produced by every male that we examined (n = 13). Both the primary song and secondary song are produced only once in each copulation interaction, unlike Lu. longipalpis s.l., where males can produce each song multiple times during the same copulation sequence31.
Sexual signaling controls the exchange of sensory information between partners and plays a direct role in divergence and speciation22,35. For example, in the Drosophila montium species subgroup the sine song frequency was suggested to be a cue indicating differences among sibling species36. In the Lu. longipalpis species complex the same function may be performed by the pulse and burst pattern songs18,19,31. Phlebotomus argentipes s.s. belongs to a species complex and although there are two nominotypical members, Ph. annandalei Sinton 1923 and Ph. glaucus Mitra & Roy 1953, which can be distinguished by morphological characters37, the full extent of the species complex is unclear. It would be interesting to analyze the songs of males from populations of the known species complex members as well as within Ph. argentipes s.s., particularly in areas of VL transmission, to evaluate the possibilities that acoustic signals are involved in reproductive isolation within this species.
The study of acoustic communication in vector insects, such as mosquitoes and sandflies, can provide a useful tool in vector control programs, such as the potential to design sound traps or for the assessment of male mating competitiveness in relation to control based on modified male release programs in the field.
Conclusions
Our results show that Ph. argentipes males produce copulatory courtship songs. Two types of patterns are observed, a primary song similar to P1 subtype previously described in a sibling species of the Lu. longipalpis species complex, and a subsequent secondary sine song that has not been seen previously in Lu. longipalpis. Our analysis represents the first report of the acoustic signals produced during copulation in Ph. argentipes and supports the idea that acoustic communication might be widespread in sandflies, including the Old World species. Future study is required to identify song patterns in other putative members of the Ph. argentipes species complex and to determine whether copulatory courtship song is important for sexual communication in Old World sandflies.
Methods
The Ph. argentipes specimens used in this study were obtained from a colony maintained at Keele University, UK, for more than 40 generations at 27 °C, 95% RH, under a 12:12 light:dark photocycle. The colony originated from wild-stock collected near Pune, India, on the east side of India, in a region where there is no visceral leishmaniasis, also known as kala-azar. Recordings were performed according to Souza et al.11. A virgin male and a virgin female between 2–7 days old were placed inside a small square acrylic chamber (1 cm × 1,5 cm × 0,5 cm) for up to 5 min at 25 ± 1 °C. Recordings were made during afternoon hours until dusk using an INSECTABOX38 and a Sony Hi8CCD-TRV65 video camera and Panasonic DMRES10 DVD recorder. Songs were analyzed using DataView version 10.3.0 (Heitler W.J. University of St Andrews, Scotland). Four parameters were measured: (1) IPI, inter-pulse interval; (2) NP, pulses per train; (3) TL, train length; and (4) Freq, the carrier frequency of the pulse train. An Additional file shows the parameters per each individual used for the mean value estimates [see Additional File 2]. The song spectrogram was obtained from Raven version 2.039.
References
Ewing, A. W. Arthropod Bioacoustics. Neurobiology and Behavior. Comstock, Cornell University Press, Ithaca, New York; p. 260 (1989).
Drosopoulos, S. & Claridge, M. F. Insect sounds and communication: physiology, behavior, ecology and evolution (contemporary topics in entomology), CRC press, Boca Raton, p. 552 (2005).
Lande, R. Models of speciation by sexual selection on polygenic traits. Proc. Nat. Acad. Sci. USA 78, 3721–3725 (1981).
Gleason, J. M. & Ritchie, M. G. Evolution of courtship song and reproductive isolation in the Drosophila willistoni species complex: do sexual signals diverge the most quickly? Evolution. 52, 1493–1500, https://doi.org/10.1111/j.1558-5646.1998.tb02031.x (1998).
Ritchie, M. G. Sexual selection and speciation. Annu. Rev. Ecol. Evol. Syst. 38, 79–102, https://doi.org/10.1146/annurev.ecolsys.38.091206.095733 (2007).
Gleason, J. M. Mutations and natural genetic variation in the courtship song of Drosophila. Behav. Genet. 35, 265–277, https://doi.org/10.1007/s10519-005-3219-y (2005).
Stone, C. M., Tuten, H. C. & Dobson, S. L. Determinants of male Aedes aegypti and Aedes polynesiensis (Diptera: Culicidae) response to sound: efficacy and considerations for use of sound traps in the field. J. Med. Entomol. 50(4), 723–730 (2013).
Vigoder, F. M., Ritchie, M. G., Gibson, G. & Peixoto, A. A. Acoustic communication in insect disease vectors. Mem. Inst. Oswaldo Cruz. 108(1), 26–33, https://doi.org/10.1590/0074-0276130390 (2013).
Souza, N. A., Brazil, R. P. & Araki, A. S. The current status of the Lutzomyia longipalpis (Diptera: Psychodidae: Phlebotominae) species complex. Mem. Inst. Oswaldo Cruz. 112(3), 161–174, https://doi.org/10.1590/0074-02760160463 (2017).
Souza, N. A., Ward, R. D., Hamilton, J. G. C., Kyriacou, C. P. & Peixoto, A. A. Copulation songs in three siblings of Lutzomyia longipalpis (Diptera: Psychodidae). Trans. R. Soc. Trop. Med. Hyg. 96, 102–103 (2002a).
Souza, N. A. et al. Analysis of the copulatory courtship songs of Lutzomyia longipalpis in six populations from Brazil. J. Med. Entomol. 41(5), 906–913, https://doi.org/10.1603/0022-2585-41.5.906 (2004).
Souza, N. A., Andrade-Coelho, C. A., Vigoder, F. M., Ward, R. D. & Peixoto, A. A. Reproductive isolation between sympatric and allopatric Brazilian populations of Lutzomyia longipalpis s.l. (Diptera: Psychodidae). Mem. Inst. Oswaldo Cruz. 103, 216–219 (2008).
Wulff, N. C., Schöneich, S. & Lehmann, G. U. C. Female perception of copulatory courtship by male titillators in a bushcricket. Proc. Biol. Sci. Aug 15, 285(1884), https://doi.org/10.1098/rspb.2018.1235 (2018).
Wulff, N. C., van de Kamp, T., Dos Santos Rolo, T., Baumbach, T. & Lehmann, G. U. Copulatory courtship by internal genitalia in bushcrickets. Sci. Rep. Feb 7(7), 42345, https://doi.org/10.1038/srep42345 (2017).
Wen, S. Y. et al. Copulatory courtship behavior and sine song as a mate recognition cue in Drosophila lini and its sibling species. Zoolog. Sci. Jul. 28(7), 469–75, https://doi.org/10.2108/zsj.28.469 (2011).
Chen, C. C. et al. Courtship patterns in the Drosophila montium species subgroup: repeated loss of precopulatory courtship? Zoolog. Sci. Dec 30(12), 1056–62 (2013).
Edvardsson, M. & Arnqvist, G. Copulatory courtship and cryptic female choice in red flour beetles Tribolium castaneum. Proc. Biol. Sci. Mar 22(267(1443)), 559–63 (2000).
Araki, A. S. et al. Molecular and behavioral differentiation among Brazilian populations of Lutzomyia longipalpis (Diptera: Psychodidae: Phlebotominae). PLoS Negl. Trop. Dis. 3, e365, https://doi.org/10.1371/journal.pntd.0000365 (2009).
Vigoder, F. M. et al. Phenotypic differentiation in love song traits among sibling species of the Lutzomyia longipalpis complex in Brazil. Parasit. Vectors. 8, 290, https://doi.org/10.1186/s13071-015-0900-8 (2015).
Vigoder, F. M. et al. Lovesongs and period gene polymorphisms indicate Lutzomyia cruzi (Mangabeira, 1938) as a sibling species of the Lutzomyia longipalpis (Lutz and Neiva, 1912) complex. Infect. Genet. Evol. 10(6), 734–9, https://doi.org/10.1016/j.meegid.2010.05.004 (2010a).
Vigoder, F. M., Souza, N. A. & Peixoto, A. A. Copulatory courtship song in Lutzomyia migonei (Diptera: Psychodidae). Mem. Inst. Oswaldo Cruz. 105(8), 1065–1067, https://doi.org/10.1590/S0074-02762010000800020 (2010b).
Vigoder, F. M., Souza, N. A. & Peixoto, A. A. Acoustic signals in the sand fly Lutzomyia (Nyssomyia) intermedia (Diptera: Psychodidae). Parasit. Vectors. 4, 76, https://doi.org/10.1186/1756-3305-4-76 (2011).
Greenspan, R. J. & Ferveur, J. F. Courtship in Drososphila. Annu. Rev. Genet. 34, 205–232, https://doi.org/10.1146/annurev.genet.34.1.205 (2000).
Maroli, M., Feliciangeli, M. D., Bichaud, L., Charrel, R. N. & Grandoni, L. Phlebotomine sandflies and the spreading of leishmaniases and other diseases of public health concern. Med. Vet. Entomol. 27, 123–147, https://doi.org/10.1111/j.1365-2915.2012.01034.x (2013).
Akhoundi, M. et al. A historical overview of the classification, evolution, and dispersion of Leishmania parasites and sandflies. PLoS Negl. Trop. Dis. 10(3), e0004349, https://doi.org/10.1371/journal.pntd.0004349 (2016).
Bray, D. P. & Hamilton, J. G. C. Courtship behavior in the sandfly Lutzomyia longipalpis, the New World vector of visceral leishmaniasis. Med. Vet. Entomol. 2, 332–338, https://doi.org/10.1111/j.1365-2915.2007.00700.x (2007).
Bray, D. P. et al. Multi-modal analysis of courtship behavior in the Old World leishmaniasis vector Phlebotomus argentipes. PLoS Negl. Trop. Dis. 8(12), e3316, https://doi.org/10.1371/journal.pntd.0003316 (2014).
Lane, R. P., Pile, M. M. & Amerasinghe, F. P. Anthropophagy and aggregation behavior of the sandfly Phlebotomus argentipes in Sri Lanka. Med. Vet. Entomol. 4(1), 79–88 (1990).
Morrison, A. C. et al. Nocturnal activity patterns of Lutzomyia longipalpis (Diptera, Psychodidae) at an endemic focus of visceral leishmaniasis in Colombia. J. Med. Entomol. 32, 605–617 (1995).
Suvanto, L., Hoikkala, A. & Liimatainen, J. O. Secondary courtship songs and inhibitory songs of Drosophila virilis-group males. Behav. Genetics. 24(1), 85–94 (1994).
Souza, N. A. et al. Analysis of the copulatory courtship songs of Lutzomyia longipalpis in six populations from Brazil. J. Med. Entomol. 41(5), 906–913, https://doi.org/10.1603/0022-2585-41.5.906 (2004).
Von Schilcher, F. The function of pulse song and sine song in the courtship of Drosophila melanogaster. Anim. Behav. 24, 622–625 (1976).
Warren, B., Gibson, G. & Russell, I. J. Sex recognition through midflight mating duets in Culex mosquitoes is mediated by acoustic distortion. Curr. Biol. 19(6), 485–91, https://doi.org/10.1016/j.cub.2009.01.059 (2009).
Cator, L. J., Arthur, B. J., Harrington, L. C. & Hoy, R. R. Harmonic convergence in the love songs of the dengue vector mosquito. Science. 323, 1077–1079, https://doi.org/10.1126/science.1166541 (2009).
Kirkpatrick, M. Sexual selection and the evolution of female choice. Evolution. 36, 1–12 (1982).
Wen, S. et al. Copulatory courtship behavior and sine song as a mate recognition cue in Drosophila lini and its sibling species. Zoolog. Sci. 28(7), 469–75, https://doi.org/10.2108/zsj.28.469 (2011).
Ilango, K. A taxonomic reassessment of the Phlebotomus argentipes species complex (Diptera: Psychodidae: Phlebotomine). J. Med. Entomol. 47(1), 1–15 (2010).
Gorczyca, M. & Hall, J. C. The INSECTAVOX, an integrated device for recording and amplifying courtship songs of Drosophila. Drosophila Inform. Serv. 66, 157–160 (1987).
Charif, R. A., Ponirakis, D. W. & Krein, T. P. Raven Lite 2.0. Cornell Laboratory of Ornithology, Ithaca, NY, www.birds.cornell.edu/raven (2006).
Acknowledgements
We would like to thank Dra. Rafaela Bruno for laboratory supplies, Dr. João Gesto for support in figure design, Kashinath Gosh, Ann Underhill and Khatijah Yaman for collected and maintaining the Phlebotomus argentipes colony at Keele University. We would also like to thank Dra. Nicola Cook and two anonymous reviewers for helpful comments on the manuscript. Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES-PNPD), Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq), Fundação Oswaldo Cruz, the Malaysian Government and the Wellcome Trust.
Author information
Authors and Affiliations
Contributions
A.S.A. and F.M.V. performed the experiments and analyzed the data. A.S.A. prepared figures. A.S.A., R.P.B., J.G.C.H. and F.M.V. wrote the main manuscript text. All authors reviewed the manuscript.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Araki, A.S., Brazil, R.P., Hamilton, J.G.C. et al. Characterization of copulatory courtship song in the Old World sand fly species Phlebotomus argentipes. Sci Rep 10, 5116 (2020). https://doi.org/10.1038/s41598-020-61867-6
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-020-61867-6
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
