
Abstract
Hearing and balance rely on the transduction of mechanical stimuli arising from sound waves or head movements into electrochemical signals. This archetypal mechanoelectrical transduction process occurs in the hair-cell stereocilia of the inner ear, which experience continuous oscillations driven by undulations in the endolymph in which they are immersed. The filamentous structures called tip links, formed by an intertwined thread composed of an heterotypic complex of cadherin 23 and protocadherin 15 ectodomain dimers, connect each stereocilium to the tip of the lower sterocilium, and must maintain their integrity against continuous stimulatory deflections. By using single molecule force spectroscopy, here we demonstrate that in contrast to the case of classical cadherins, tip-link cadherins are mechanoresilient structures even at the exceptionally low Ca2+ concentration of the endolymph. We also show that the D101G deafness point mutation in cadherin 23, which affects a Ca2+ coordination site, exhibits an altered mechanical phenotype at the physiological Ca2+ concentration. Our results show a remarkable case of functional adaptation of a protein’s nanomechanics to extremely low Ca2+ concentrations and pave the way to a full understanding of the mechanotransduction mechanism mediated by auditory cadherins.
Similar content being viewed by others


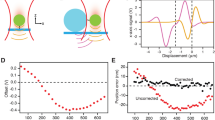
Introduction
Hearing and balance perception in vertebrates is considered as one of the most evident mechanotransduction processes1. In particular, the hair cells of the inner ear contain a bundle of ascending stereocilia that are deflected by forces produced by undulations that propagate in the endolymph arising from sound waves or head movements2,3,4,5,6,7. These deflections produced by mechanical tension promote the rapid opening of the so-called mechano-electrical transduction (MET) channels and the subsequent depolarization of hair cells8,9,10. This strongly coupled mechano-electrical transduction process requires robust extracellular structures, not only to transmit the tension from the upper to the lower stereocilium leading to the opening of the MET channel, but also to maintain the bundle integrity after continuous mechanical stimulations (Fig. 1a). Among other connectors between the stereocilia, the so-called tip-link, a long intertwined filament formed by a helical dimer of cadherin 23 (CDH23) in the apical end bound head-to-head to a helical dimer of protocadherin 15 (PCDH15) in the basal end, is particularly relevant for the proper gating process and coupling of the MET response2,4,5,6,7.

Nanomechanics of the full ectodomain from tip-link cadherins. (a) Schematic cartoon of the connections between sterocilia from the hair cells of the inner ear. Tip links are formed by a trans interaction of a single cis dimer of CDH23 (magenta) and a cis dimer of PCDH15 (green). The location of the MET channel is indicated. Lateral connections (indicated with blue horizontal lines) between sterocilia are schematically drawn. (b) Typical experimental setup in AFM-SMFS. (c) Nanomechanics of full-length CDH23 in different Ca2+ conditions. Although we were able to stretch almost 1 μm in z direction, no more than 15 peaks were obtained in single-molecule recordings. (d) Detailed view of the proximal region of a force-extension recording at 1 mM Ca2+ showing calcium rivets17 and EC domain unfolding peaks, which are delimited by the WLC fit (in green). (e) Nanomechanics of full-length PCDH15 in the mentioned Ca2+ conditions. The calcium rivets are marked in the histograms with filled bars (c,e). In (c) and (e) a high number of non-monomeric recordings were obtained, which were discarded following the criteria explained in Methods (Fig. S6c,d). Data is presented from low (top) to high (bottom) Ca2+ concentrations (c,e).
Both CDH23 and PCDH15 belong to the Cadherin Superfamily, a large Ca2+-binding protein family responsible for cell-cell adhesion11. These are single transmembrane proteins with CDH23 containing 27 extracellular cadherin (EC) domains and PCDH15 containing 11 EC domains. It has been shown that Ca2+ binding to the tip link is critical for the proper mechanotransduction process in hair cells, since in the absence of this cation the tip-link filament becomes dismantled and signal transduction gets interrupted8,12. However, this effect is reversible once extracellular Ca2+ levels are restored, then the tip-link filament is rebuilt13. Interestingly, several point mutations related with decreased Ca2+ binding have also been linked to inherited forms of deafness in humans and rodents, which appear to compromise mechanotransduction14,15,16. We have previously shown the critical role of Ca2+ binding in the rigidification and mechanical response of cadherin domains from classical cadherins, and that this binding is also responsible for an additional element of mechanical resistance (extramodular, i.e., outside the mechanical clamp of the EC module), termed the “calcium rivet”17. Furthermore, we also showed that17 the mechanical stability of classical cadherins is significantly reduced at the exceptionally low Ca2+ concentration of the endolymph that bathes the tip-link (20–40 μM free Ca2+)18,19. Still, the tip link is stably formed at 50 μM free Ca2+ but if the level of Ca2+ is reduced, then the length of the filament increases, probably due to a loss of rigidity or EC domain unfolding20. Therefore, tip-link cadherins must show particular Ca2+ binding properties compared to classical cadherins, despite the high sequence identity between their Ca2+-binding regions and their similar folds21,22. However, the intrinsic mechanical properties of this critical mechanotransducer have not been experimentally characterized to date.
Here we report the mechanical properties of the tip-link cadherins and demonstrate that, unlike classical cadherins17, these proteins are remarkably mechanostable structures at the low physiologically relevant Ca2+ concentration of the endolymph, in agreement with their pivotal role in mechanotransduction. Furthermore, we also show that a single point mutation related to DFNB12 hereditary deafness, which impairs Ca2+ coordination22, affects the nanomechanics of the domains, but only at physiological Ca2+ levels. Taken together, our data provide a necessary molecular insight into this important mechanotransduction system, setting the limits of its proper mechanical response.
Results
Nanomechanics of tip-link cadherins
Since tip-link cadherins are crucial components of the mechanotransduction machinery of sound stimuli1, their mechanical stability is a critical parameter that will determine their proper function. Moreover, because of their long, multimodular structure and in vivo geometry4, tip-link cadherins are ideal candidates for single-molecule force spectroscopy (SMFS)23. Interestingly, as previously demonstrated for classical cadherins17, we can characterize the role of Ca2+ coordination modulating the robustness of tip-link cadherins, which is a critical parameter based on the already known effects of Ca2+ binding in tip-link formation and rigidity8,12,22.
We have produced full-length murine CDH23 and PCDH15 ectodomains in mammalian cells and analysed their mechanical stability at the single molecule level (Fig. 1). Upon mechanical stretching (Fig. 1b), the full-length CDH23 ectodomain showed the characteristic saw-tooth pattern typical of multi-domain protein unfolding, where each individual force peak represents the unravelling of an individual domain (Fig. 1c). The magnitude of the peak results from the rupture of the main resistance barrier of the domain to mechanical unfolding (the so-called mechanical clamp)23, which in cadherin EC domains corresponds to a patch of hydrogen bonds between two parallel β-strands that secure the domain17 (Fig. 2a). In addition, several force peaks of the recordings are preceded by smaller amplitude peaks with a short increase in contour length (ΔLc; Fig. 1d) that originates from the rupture of the Ca2+ coordination complexes17 and disappear at sub-μM Ca2+ concentrations (1 mM EGTA, Supplementary Table 1). This additional resistance element to mechanical unfolding known as the calcium rivet, which is also present in classical cadherins upon Ca2+ coordination, ensures that the EC domain remains folded when subjected to low range forces17. Interestingly, the calcium rivets may break collectively at low loading forces, meaning that several calcium rivets may appear before the first domain unfolding peaks in the recording. After an EC unfolding event and a short relaxation time until the next unfolding event (0.1 s due to the feedback latency of the AFM piezoelectric device), those rivets ruptured at low forces, which are located between EC domains that remain folded, could reassemble giving rise to additional rivets preceding the following force peaks in the recording (Fig. 1d). Therefore, the more Ca2+ coordination complexes the protein is able to form, the more evident (with larger ΔLc) and abundant the calcium rivets will appear in the force recordings. Indeed, the calcium rivets observed for CDH23 (with 27 EC domains, Fig. 1b–d) are more obvious than those shown by classical cadherins (consisting of just 5 EC domains17). However, since the rupture of Ca2+ coordination networks is a stochastic and cooperative process, not all the EC domain unfolding peaks are preceded by calcium rivets.

DFNB12 hereditary deafness-associated D101G mutation shows mechanical differences. (a) Schematic representation of the pFS + EC1–2 construction. The fusion protein pFS-1 was explained in detail elsewhere25. The inset at the bottom shows the atomic structure of CDH23 EC1–2D101G domains22, showing the coordinated Ca2+ ions as cyan spheres, and the residues involved in coordination in bonds representation. Also of note is the characteristic N-terminal Ca2+ ion present in tip-link cadherins. The two mechanical determinants showed by these proteins (the mechanical clamps from the EC domains and the calcium rivet17) are indicated. The atomic structures shown in this Figure were depicted using VMD 1.8.641. (b) WT CDH23 EC1–2 domains show canalized unfolding in the low Ca2+ concentration present in the endolymph. Several representative force-extension recordings are shown, with the ubiquitin (U in a) peaks in grey, the N2B spacer in light grey (belonging to the N2B spring from human cardiac titin, which acts as a spacer from the noisy proximal region of the recordings25), and the EC peaks in the corresponding color. The fitting of the WLC model37 is shown in green, enabling the observation of the unfolding decanalization of the EC domains in EGTA (top recording), as well as the clear deviation of the calcium rivet from the WLC fitting to the main unfolding peaks (in lighter colors in the spectra). (c) F histograms of WT EC1–2. The F values for the rupture of the calcium rivets are shown in solid bars in the histograms. (d) F histograms of EC1–2D101G. In EGTA the values are comparable to those of decanalized EC domains at 30 μM Ca2+. (e) Unlike the WT, EC1–2D101G showed a high unfolding decanalization in a physiological Ca2+ concentration (~30 μM Ca2+). For simplicity, only data coming from EC domains are plotted in the histograms.
Thus, as observed for classical cadherins, CDH23 at saturating Ca2+ concentrations showed a “canalized”17 mechanical unfolding, consisting of discrete EC domain unfolding events preceded by the rupture of calcium rivets (Fig. 1c,d). However, although the mechanical stability of CDH23 was comparable to that of classical cadherins at saturating Ca2+ conditions (Supplementary Table 117), significant differences in the mechanical behaviour of CDH23 are observed at physiological Ca2+ concentrations. Specifically, in contrast to classical cadherins, which show “decanalized” (i.e. disorganized) unfolding already at 100 μM Ca2+ (Fig. S1a–c17), CDH23 presented a canalized unfolding in the very low Ca2+ concentrations of the endolymph (≈30 μM)18,19, meaning that calcium rivets are present at extraordinarily low Ca2+ levels showing comparable mechanostabilities (Fig. 1c,d and Supplementary Table 117).
Similarly, full-length PCDH15 also showed a canalized mechanical behaviour at 30 μM Ca2+ (Fig. 1e, Supplementary Table 2). Therefore, although there is a wide range of Ca2+ affinity values across their whole ectodomains22,24, tip-link cadherins contain novel high affinity Ca2+ binding sites (Fig. S1d) that enable them to remain resistant to mechanical unfolding even if their Ca2+ binding sites are partially saturated at physiological conditions21,22. Interestingly, in the case of PCDH15, which contains 11 EC domains4, no more than 10 EC peaks were observed per SMFS recording (Fig. 1e); probably the missing EC was denatured or covalently clamped through a disulphide bond7, obscured by the noisy proximal region of the recordings25, or showed decanalized unfolding because of lack of Ca2+ coordination in that particular region24. The first peak of the recording corresponds to the detachment of the cantilever tip from the substrate or, alternatively, from the rupture of unspecific interactions from the tip. It is common that the proximal region of the recordings gets obscured by these noisy, non-specific interactions when the protein is detached from the tip or the substrate. The final peak(s) of the recordings corresponds to the detachment of the protein from either the tip or the substrate, which marks the end of the single-molecule experiment23.
A deafness-associated mutation shows a mechanical phenotype
Since tip-link cadherins showed differential Ca2+ modulation of their mechanical properties, we aimed to analyze specific domains in different Ca2+ concentrations in order to characterize the possible mechanical effect of deafness-related mutations (Figs 2 and S215,26,27). To fully characterize the nanomechanics of the EC modules, we used the pFS-1 polyprotein strategy as a single-molecule marker (Fig. 2a)25. We analyzed many individual EC modules as recombinant proteins (Fig. S2) and, although all the individual EC constructs seemed properly folded (in the different Ca2+ concentrations tested) in a conformation rich in β-structure as measured by CD spectroscopy (Figs S3 and S4), we found only a single construct that enabled a full mechanical characterization: pFS + ECCDH231–2. Using this construct, we additionally characterized the D101G deafness-related mutation (Fig. 2), which is known to affect this CDH23 region’s conformation and dynamics, as a consequence of a reduced Ca2+ binding22. Indeed, the mutation shows a clear mechanical phenotype, but only at low physiological Ca2+ concentrations (Supplementary Table 3).
Specifically, both EC1–2 WT and its D101G variant showed mechanical stabilities comparable to those of the full-length CDH23 at 1.0 mM Ca2+ concentration (Figs 1 and 2 and Supplementary Tables 1 and 3). This observation indicates that the flanking pFS-1 domains (Fig. 2a) do not alter the mechanical properties of the EC modules under study, and therefore are not the source of the altered nanomechanics observed for other EC domains shown in this study (Figs S1 and S2). Remarkably, D101G mutation caused a mechanical phenotype on CDH23 EC1–2 reducing the mechanostability and de-canalizing the unfolding, which begins to be apparent at 300 μM Ca2+ and becomes more evident at 30 μM Ca2+ (Fig. 2c–e and Supplementary Table 3). Thus, it seemed critical to analyse its mechanical properties in physiological conditions. At 300 μM Ca2+, 88% of EC1–2D101G molecules (36 out of 41) showed canalized unfolding of both EC domains, while at 30 μM Ca2+ this percentage decreased dramatically to 31% (14 out of 45 molecules; Supplementary Table 3). Remarkably, no molecules with decanalized unfolding were observed in these conditions for EC1–2 WT, indicating that Ca2+ binding to the wild type protein strengthens the EC domains against mechanical unfolding at physiological conditions (Fig. 2e). In contrast, D101G reduces Ca2+ binding22, compromising the mechanical response of this region of CDH23 (Fig. 2d,e). Indeed, since the proportion of the canalized and decanalized unfolding pathways changes when varying the Ca2+ concentration, these results strongly suggest that the sensitivity to Ca2+ concentration is altered in the deafness mutant. As a confirmation that a decanalized mechanical unfolding is due to impaired Ca2+ binding, both EC1–2 WT and EC1–2D101G showed comparable properties in the absence of Ca2+ (sub-μM Ca2+ concentration) (Fig. 2b–e and Supplementary Table 3).
More notably than in full-length ectodomains (Fig. 1c,e), the average force values of unfolding of EC1–2 domains lower with decreasing Ca2+ concentrations, likely because EC1–2 Ca2+ affinity remains in the low range of the average affinity of the whole CDH23 ectodomain, as observed for classical cadherins28 (Fig. 2d). As mentioned, this effect is more evident for EC1–2D101G. At 30 μM Ca2+, EC1–2D101G domains show, on average, 103 ± 63 pN of resistance to unfolding, while the calcium rivet breaks at 129 ± 35 pN (Supplementary Table 3). This result is surprising since the calcium rivets are assumed to work as sacrificial mechanical resistance elements that prevent the unfolding of EC domains, breaking before the main force peak of domain unfolding and showing lower forces17. However, only 5 out of 36 molecules showed a calcium rivet, which in turn also show the highest forces values within the distribution for EC unfolding at this condition. Therefore, the F value indicated for EC domain unfolding in Supplementary Table 3 contains the large contribution of EC domains that do not show calcium rivet and manifest lower mechanical stabilities, which lowers the overall F value.
Discussion
Channel gating in the inner ear is an extremely sensitive mechanotransduction system in which activation has been recorded with only 3 pN of applied tension and relative displacements of 1 nm or deflections of 1°, with <10 μs of time response29,30,31,32. Therefore, it is of paramount importance to determine the consequences of differential ion binding on its mechanical response to understand this system’s molecular bases and how Ca+2 binding defects uncouple the transmission of mechanical tension to channel gating, producing deafness3. We have shown that tip-link cadherins present a canalized mechanical unfolding at the low Ca2+ concentrations of the endolymph, in contrast to the behavior of classical cadherins, which show decanalized mechanical unfolding at those concentrations17 (Fig. S1a–c). These different properties can be explained as resulting from the differences in coordination affinities displayed by the distinctive Ca2+ binding sites present in tip-link cadherins21,22 (Fig. S1d). In general, it has been proposed that the specific Ca2+-binding affinities of the two most distal EC domains in CDH23 are about 2 orders of magnitude higher than those seen in the equivalent domains of classical cadherins22,28. Therefore, it follows that tip-link cadherins have developed a tighter regulation of their function by Ca2+ binding. Moreover, since they are assumed to function just as force transmitters, Ca2+ binding is expected to strongly determine their mechanical properties. Nonetheless, despite the high conservation of Ca2+ coordination sites among cadherins (Fig. S1d), there are remarkable differences in binding affinities22. Moreover, D101G mutation largely affects the binding affinities in all binding sites between EC1–2 domains22, which can be explained in terms of altered rigidity of this interdomain region in the mutant22, because of the increased entropy of Gly in the unfolded state. Therefore, a partially rigid interdomain linker is necessary for a proper Ca2+ coordination, which in turn provides additional rigidity, as if Ca2+ coordination stabilizes particular conformations, via a conformational selection mechanism24,33. In the future, identifying individual EC domains in the SMFS recordings obtained from full-length ectodomains would be particularly relevant for studying the mechanical effects of deafness-related mutations in the context of the whole protein.
We have previously proposed the cadherin ectodomain as a “Ca2+ switch”, because the mechanical properties dramatically change when the Ca2+ concentration is depleted17. However, since the main physiological role of cadherin ectodomains is to maintain strong extracellular connections, the EC domains must remain intact at the range of forces supported by these connections. Although the tip link is known to be sensitive to extremely low forces29, it is also subjected to continuous mechanical stimuli (Fig. 3). Therefore, for the proper functioning of the tip-link as a force transmitter, tip-link cadherin ectodomains seem to have acquired tighter Ca2+ affinity22, and have preserved the calcium rivet at the low physiological Ca2+ concentration, suggesting that the calcium rivet is an important mechanical component that contributes with additional mechanical resistance to impede the unraveling of the domain upon stretching in vivo. D101G shows a reduced number of calcium rivets at physiological Ca2+ concentration (Fig. 2) and thus the mutated domains are more exposed to an eventual unfolding event upon stretching, which will uncouple the mechanical transduction system leading to the lack of electrical response to sound stimuli (Fig. 3). Nonetheless, in order to demonstrate the relevance of the calcium rivet and the mechanical phenotype in disease, the mutated domain should unfold before the unbinding of the tip-link. Thus, a single tip-link should resist forces above ∼50 pN29, which is the statistical value where many EC D101G domains unfold at the applied loading rate (Fig. 2d).

Explanatory model for the mechanical effect of the D101G mutation. After a stimulus, the WT modules will remain folded since they are able to coordinate Ca2+ in all the sites and remain canalized. Thus, they will transmit the tension generated by undulations in the endolymph opening the coupled MET channels and generating the hearing response. However, in the case of DFNB12 hereditary deafness-related D101G mutation, the mutated modules will be unable to coordinate Ca2+. After a stimulus, they will show decanalized unfolding and will unravel showing no resistance. The tension will not be transmitted to the lower sterocilium, likely uncoupling the mechanism of channel gating and subsequent hearing response, causing deafness. After a continuous stimulation, the reduced mechanical stability of the mutated modules can cause structural defects in the module that could lead to dismantling of the tip link and therefore, the possible interruption of the mechanotransduction circuit.
It must be noted that, statistically, fewer calcium rivet events were observed at the 30 μM physiological Ca2+ concentration (particularly for PCDH15, Fig. S5). This is probably because not all the coordination sites were occupied by Ca2+ 22. Indeed, the mechanical effect observed for the D101G mutant at 30 μM Ca2+ (Fig. 2) might be due to incomplete Ca2+ coordination. Altogether, our results suggest that when Ca2+ is released, WT structures of both CDH23 and PCDH15 fragments are affected, showing differences in secondary structure elements (Figs S3 and S4)33 and lower mechanostability (Supplementary Tables 1–3). Interestingly, we were able to observe the reversibility of the “canalization” of domain unfolding upon Ca2+ binding (Fig. 1), in line with the reversible Ca2+-dependent plasticity of tip-links observed in vivo13. Furthermore, upon mutation of these proteins, the modules appear destabilized and their mechanical properties get altered showing a lowered mechanostability (Supplementary Table 3).
A decrease in Ca2+ concentration causes dismantling of the tip-links20. Thus, there is clear correlation between Ca2+ coordination, conformational constraints22, mechanical stability and tip-link formation, and these properties might determine the proper mechanotransduction in the hair cells3. This indicates that slight changes in the mechanical properties of the EC domains can cause long-range effects in the whole tip-link, finally leading to the formation of a less robust complex and the concomitant dysfunction of the mechanotransduction process (Fig. 3)24. The effect of other deafness-related mutations in complex formation has been described3,22,24. However, our study represents the first experimental evidence of altered mechanical properties of tip-link cadherins at physiological Ca2+ concentrations. Because tip-link cadherins play a critical mechanical role in the mechanotransduction process of sound stimuli, the study of their mechanical properties is of paramount importance to understand the molecular mechanisms underlying the transduction of sound and movement stimuli into electrochemical signals. Moreover, the mechanical phenotype of the disease-related mutation studied here is only evident at the low physiological Ca2+ concentration, highlighting the importance of characterizing the mechanical properties of tip-link cadherins at their real physiological conditions, rather than using the standard 1.0 mM Ca2+ concentration to emulate Ca2+ content in extracellular environment.
Methods
Protein expression and purification of CDH23 and PCDH15
The His-tagged full murine CDH23 and PCDH15 ectodomains (UniProtKB code Q99PF4 and Q99PJ1, respectively) were expressed in HEK293 cells and purified according to4. Cells were cultured in DMEM media supplemented with 10% serum, 1% penicillin/streptomycin and 20 mM HEPES (Gibco). Cell transfection was performed using TurboTransfect (ThermoScientific). Twenty-four hours after transfection, the medium was replaced with F-12 Nutrient Mix (without serum, Sigma). Cells were incubated 48 hours in this medium, when recombinant proteins were overexpressed and secreted to the extracellular media. Supernatants were collected and filtered in Amicon 10-kDa cutoff filters (Millipore) and buffer exchanged to 50 mM TrisHCl/150 mM NaCl/5 mM imidazole [pH 7.5], followed by a Ni2+-affinity purification using Histrap HP columns (GE Healthcare). Proteins were concentrated using Amicon filters and buffer exchanged to 50 mM TrisHCl/150 mM NaCl/1 mM CaCl2 [pH 7.5]. Protein purity was monitored by SDS-PAGE and Western blotting using anti-Histag antibodies (both proteins contain a C-terminal Histag, see4). Proper morphology of the full-length ectodomains was assessed by imaging AFM (see below). Pure proteins were stored at 4 °C at a concentration of ~0.2–0.5 mg/ml.
Cloning of EC domains
We used the pFS-1 vector, containing ubiquitin repeats as single-molecule markers, to characterize the individual CDH23 and PCDH15 ectodomains (EC) at the single-molecule level25. The first two EC domains from CDH23 were analyzed in both the wild type form and containing the D101G mutation (located between both domains)15 in pFS-1. An additional construct containing EC6–8 from CDH23 was built to analyze the E737V mutation (located between EC7-8)27 but the pFS + EC6–8 fusion protein precluded the proper folding of EC6–8 domains (Fig. S2a). In addition, since R139G and G267D mutations are present in PCDH15 EC1–226, the first two EC domains from PCDH15 were cloned onto pFS-1, both in WT and mutated form. Since the G267D mutation is located close to EC2 C-terminus, similar constructs but containing the first three ECs from PCDH15 were also produced for comparative purposes. None of these EC domains from PCDH15 seemed properly folded when fused to pFS-1, and therefore their nanomechanical properties could not be analyzed (Fig. S2b,c).
The same combinations of domains were individually cloned into the pET28a vector (Novagen) for Circular Dichroism (CD) measurements (Figs S3 and S4). Specifically, EC1–3, EC1–3G267D and EC1–3R139G domains from PCDH15 and EC1–2, EC1–2D101G, EC6–8 and EC6–8E737V domains from CDH23 were produced in order to test possible structural effects at different Ca2+ concentrations.
Polyprotein expression and purification
All the polyproteins (pFS + EC1–2, pFS + EC1–2G267D, pFS + EC1–3, pFS + EC1–3R139G and pFS + EC1–3G267D from PCDH15; pFS + EC1–2, pFS + EC1–2D101G, pFS + EC6–8 and pFS + EC6–8E737V from CDH23) as well as the EC domains (EC1–3, EC1–3G267D and EC1–3R139G from PCDH15 and EC1–2, EC1–2D101G, EC6–8 and EC6–8E737V from CDH23) were expressed in E. coli C41 (DE3)34 and BL21 (DE3) strains. Cultures were grown at 37 °C until an OD600 of 0.6–0.8 was reached, when the over-expression of the recombinant polyproteins was induced with 1.0 mM IPTG during 4 h at 37 °C, whereas 100 μM IPTG was used to induce the expression of the different EC domain constructs at room temperature for ~16 h. Bacterial pellets were subsequently lysed with 1.0 mg/ml lysozyme and 1% Triton X-100.
Recombinant polyproteins were purified by Ni2+ affinity chromatography in Histrap HP (GE Healthcare) columns, using 50 mM TrisHCl/500 mM NaCl/50 mM imidazole [pH 7.5] as binding buffer, adding 500 mM imidazole in the elution buffer. Once eluted, 1.0 mM CaCl2 was added to the protein solutions. Fractions with pure protein were concentrated using Amicon 10 K filters (Millipore). A further size-exclusion chromatography was applied in a Hiload 16/60 SD 200 column (GE Healthcare) using 50 mM TrisHCl/150 mM NaCl/1 mM DTT/1 mM CaCl2 [pH 7.5] buffer (final experimental buffer). Pure monomeric proteins were concentrated with Amicon 10 K filters (Millipore). Protein purity was monitored by SDS-PAGE and Western blotting as previously described25. Recombinant EC domain constructs were purified under denaturing conditions as previously described22.
NTA-Ni 2+ functionalization of coverslips for SMFS
We used NTA-Ni2+ functionalized coverslips as substrates for SMFS to bind full-length ectodomains from the tip-link cadherins, as well as pFS-1 polyproteins, via their C- and N-terminal Histags, respectively. Coverslips were first immersed into a 20 N KOH solution overnight, after which they were transferred to a 2% 3-(mercaptopropyl)triethoxysilane (Sigma-Aldrich), 0.02% acetic acid at 90 °C for 1 h. Afterwards, they were washed in a MilliQ water flow for 1 h and then cured for 15 min in an oven at 120 °C. Next, they were transferred to a 100 mM DTT solution for 15 min and washed under a MilliQ water flow for 1 h. Then, 50 μl of a solution of 20 mg/ml maleimide-C 3-NTA (Dojindo Laboratories) dissolved in 10 mM 3-(N-morpholino)propanesulfonic acid (pH 7) were added and the substrates were incubated for 1/2 h. After a final wash in MilliQ water, a drop of 10 mM NiCl2 was added (>50 μl) on the coverslips, incubated for 10 min, and then they were washed with MilliQ water before storage.
AFM: SMFS and imaging
Approximately 20 μl of the protein sample at ~0.3 mg/ml concentration was adsorbed onto the Atomic Force Microscopy (AFM) substrate (NTA-Ni2+-functionalized glass coverslips) via the terminal His-tags and then washed with 50 mM TrisHCl/150 mM NaCl/1 mM CaCl2 [pH 7.5]. The buffer composition during SMFS experiments was the same but changing the amount of CaCl2 (0.3 mM CaCl2, 1 mM EGTA) when required. 30 μM free Ca2+ buffer was prepared using diBr2-BAPTA as chelating agent35. The chemical equilibrium was simulated with the Maxchelator program and the amount of free Ca2+ on the experimental buffers was monitored using a Kwik-Tip Ca2+ electrode (World Precision Instruments). Recombinant proteins were extensively dialysed against 50 mM TrisHCl/150 mM NaCl [pH 7.5] before further extensive dialysis in the final experimental buffer 50 mM TrisHCl/150 mM NaCl/125 μM CaCl2/100 μM diBr2-BAPTA [pH 7.5].
All the Single Molecule Force Spectroscopy (SMFS) experiments were performed using the so-called length-clamp mode at 0.4 nm/ms constant pulling speed. In this mode, the feedback loop is switched off in the contact region, so that the piezo approaches to the cantilever tip and returns back. The contact portion of the trace was fixed to 0.05% of the total length of the trace. Since the extension in our experiments was performed at 400 nm/s, in general, the total contact time was in the range of 0.1 to 0.5 s, time during which the sample deposited in the substrate can be adsorbed onto the AFM tip via non-specific interactions.
The spring constant of each individual atomic force microscope cantilever (MLCT-AUHW, Veeco Probes, with a spring constant of ~40 pN/nm; or BL-RC, Olympus, Tokyo, Japan; with a spring constant of ~30 pN/nm) was calibrated using the equipartition theorem36. The length of the protein chain under tension was calculated by fitting the wormlike chain (WLC) model of polypeptide entropic elasticity37,38,
where F is the force, p is the persistence length, x is the end-to-end length, and Lc is the contour length of the stretched protein. The measured unfolding force of each peak (F) in the so-called saw-tooth pattern was pooled and averaged. We also measured the increase in the contour length (ΔLc) as a measure of the protein length that is force hidden. Values are reported with the standard deviation. At least 4 independent experiments were performed for each condition reported in this study.
In an optimal experiment we can amount to ~10000 trials, 6% of which contains attached protein. The final success rate of analyzable traces is about 2% (194 traces per analysis). In the case of CDH23 and PCDH15, this low efficiency could be due to the absence of a fusion protein that favors attachment to the AFM tip by affinity interactions. In addition, CDH23 and PCDH15 full ectodomains were not included inside a single-molecule marker. We did not use here the strategy of flanking domains to avoid compromising the Ca2+ coordination efficiency and levels of expression. Therefore, in this case, to be sure that single molecules are being pulled, AFM images and analysis of persistence length had to be performed (Figure S6). In addition, both ΔLC and the number of unfolded modules were used as single-molecule criteria. ΔLc is the criterion used for analysis of those proteins showing “de-canalized” (i.e., polymorphic) mechanical properties (Fig. S2b,c)17. Regarding the full tip-link ectodomains (Fig. 1), only those recordings showing a saw-tooth pattern with a ΔLc close to the expected value for an EC domain (~33–35 nm)17 without intercalated peaks, and showing less force peaks than EC domains contained in the protein (27 ECs for CDH23 and 11 for PCDH15) were selected as single-molecule force unfolding recordings. Besides, since these proteins have a high tendency to dimerize (as shown in Fig. S6 and in4), the unfolding recordings should additionally show similar persistence length, p, across the unfolding curve, indicating that a single polypeptide was being stretched (p~0.34 nm)23. Indeed, we found several traces showing different p values across the unfolding trace, indicating that we often stretched two cadherins in parallel (Fig. S6c,d). Initial CDH23 calcium rivets showed p values of 0.47 ± 0.09 nm, while for PCDH15 initial rivets showed 0.49 ± 0.05 nm of persistence length. Once ruptured, p values dropped to values ≈ 0.36 nm, corresponding to the unfolding of the polypeptide chain.
Despite the extension of the piezo was set to 1 μm in z direction, which allows for full unraveling of tip-link cadherin ectodomains, full-length CDH23 showed an average of 9 ± 1 EC unfolding events per force recording, while PCDH15 showed an average of 7 ± 1 unfolded EC domains per curve. This indicates that the attachment to the tip was not strong enough to maintain the protein attached for the longer unraveling events. Indeed, we observed total Lc values of 396.0 ± 95.2 nm for CDH23 and 303.3 ± 42.2 nm for PCDH15 (corresponding to 306 and 242 nm of average total ΔLc, respectively). Because the attachment to the tip is random and the protein could be detached at any time, information on preferential or hierarchical mechanical response across the entire ectodomains is not accessible.
AFM images (Fig. S6a,b) were acquired with the protein samples diluted in 50 mM TrisHCl/150 mM NaCl/1 mM CaCl2 [pH 7.5]. A drop of the protein solution was deposited on freshly cleaved mica and the tapping mode was employed to acquire images in liquid conditions as described in39.
Circular Dichroism spectroscopy
Far-UV CD spectroscopy was performed using a JASCO-J810 spectropolarimeter (JASCO Inc.) equipped with a Peltier temperature control unit and using quartz cuvettes of 1 mm cell-path length. EC domain samples were prepared at 2–5 μM in 15 mM TrisHCl/50 mM NaCl [pH 7.5] changing only the CaCl2 concentration (1.0 mM CaCl2, 0.3 mM CaCl2, 0.03 mM CaCl2 and 1.0 mM EGTA) for each specific sample. The corrected spectra were converted into molar ellipticity ([Θ]) using the average molecular masses per residue with Spectra Manager software (Jasco Inc.). The CDNN analysis program40 was used to monitor the secondary structure content for each construct in each Ca2+ concentration.
Free Ca2+ Concentration
Ca2+ concentrations in all the experiments shown in this study refer to measured free Ca2+ concentrations as described above.
Data Availability
Additional relevant data are available from the corresponding authors upon reasonable request.
References
Gillespie, P. G. & Walker, R. G. Molecular basis of mechanosensory transduction. Nature 413, 194–202 (2001).
Ahmed, Z. M. et al. The tip-link antigen, a protein associated with the transduction complex of sensory hair cells, is protocadherin-15. J Neurosci 26, 7022–7034 (2006).
Gillespie, P. G. & Muller, U. Mechanotransduction by hair cells: models, molecules, and mechanisms. Cell 139, 33–44 (2009).
Kazmierczak, P. et al. Cadherin 23 and protocadherin 15 interact to form tip-link filaments in sensory hair cells. Nature 449, 87–91 (2007).
Pickles, J. O., Comis, S. D. & Osborne, M. P. Cross-links between stereocilia in the guinea pig organ of Corti, and their possible relation to sensory transduction. Hear Res 15, 103–112 (1984).
Siemens, J. et al. Cadherin 23 is a component of the tip link in hair-cell stereocilia. Nature 428, 950–955 (2004).
Sotomayor, M., Weihofen, W. A., Gaudet, R. & Corey, D. P. Structure of a force-conveying cadherin bond essential for inner-ear mechanotransduction. Nature 492, 128–132 (2012).
Assad, J. A., Shepherd, G. M. & Corey, D. P. Tip-link integrity and mechanical transduction in vertebrate hair cells. Neuron 7, 985–994 (1991).
Beurg, M., Fettiplace, R., Nam, J. H. & Ricci, A. J. Localization of inner hair cell mechanotransducer channels using high-speed calcium imaging. Nat Neurosci 12, 553–558 (2009).
Denk, W., Holt, J. R., Shepherd, G. M. & Corey, D. P. Calcium imaging of single stereocilia in hair cells: localization of transduction channels at both ends of tip links. Neuron 15, 1311–1321 (1995).
Halbleib, J. M. & Nelson, W. J. Cadherins in development: cell adhesion, sorting, and tissue morphogenesis. Genes Dev 20, 3199–3214 (2006).
Vollrath, M. A., Kwan, K. Y. & Corey, D. P. The micromachinery of mechanotransduction in hair cells. Annu Rev Neurosci 30, 339–365 (2007).
Zhao, Y., Yamoah, E. N. & Gillespie, P. G. Regeneration of broken tip links and restoration of mechanical transduction in hair cells. Proc Natl Acad Sci USA 93, 15469–15474 (1996).
Ahmed, Z. M. et al. Gene structure and mutant alleles of PCDH15: nonsyndromic deafness DFNB23 and type 1 Usher syndrome. Hum Genet 124, 215–223 (2008).
Astuto, L. M. et al. CDH23 mutation and phenotype heterogeneity: a profile of 107 diverse families with Usher syndrome and nonsyndromic deafness. Am J Hum Genet 71, 262–275 (2002).
Sollner, C. et al. Mutations in cadherin 23 affect tip links in zebrafish sensory hair cells. Nature 428, 955–959 (2004).
Oroz, J. et al. Nanomechanics of the cadherin ectodomain: “canalization” by Ca2+ binding results in a new mechanical element. J Biol Chem 286, 9405–9418 (2011).
Bosher, S. K. & Warren, R. L. Very low calcium content of cochlear endolymph, an extracellular fluid. Nature 273, 377–378 (1978).
Salt, A. N., Inamura, N., Thalmann, R. & Vora, A. Calcium gradients in inner ear endolymph. Am J Otolaryngol 10, 371–375 (1989).
Furness, D. N., Katori, Y., Nirmal Kumar, B. & Hackney, C. M. The dimensions and structural attachments of tip links in mammalian cochlear hair cells and the effects of exposure to different levels of extracellular calcium. Neuroscience 154, 10–21 (2008).
Elledge, H. M. et al. Structure of the N terminus of cadherin 23 reveals a new adhesion mechanism for a subset of cadherin superfamily members. Proc Natl Acad Sci USA 107, 10708–10712 (2010).
Sotomayor, M., Weihofen, W. A., Gaudet, R. & Corey, D. P. Structural determinants of cadherin-23 function in hearing and deafness. Neuron 66, 85–100 (2010).
Galera-Prat, A., Gomez-Sicilia, A., Oberhauser, A. F., Cieplak, M. & Carrion-Vazquez, M. Understanding biology by stretching proteins: recent progress. Curr Opin Struct Biol 20, 63–69 (2010).
Powers, R. E., Gaudet, R. & Sotomayor, M. A Partial Calcium-Free Linker Confers Flexibility to Inner-Ear Protocadherin-15. Structure 25, 482–495 (2017).
Oroz, J., Hervas, R. & Carrion-Vazquez, M. Unequivocal single-molecule force spectroscopy of proteins by AFM using pFS vectors. Biophys J 102, 682–690 (2012).
Ahmed, Z. M. et al. PCDH15 is expressed in the neurosensory epithelium of the eye and ear and mutant alleles are responsible for both USH1F and DFNB23. Hum Mol Genet 12, 3215–3223 (2003).
Schwander, M. et al. A mouse model for nonsyndromic deafness (DFNB12) links hearing loss to defects in tip links of mechanosensory hair cells. Proc Natl Acad Sci USA 106, 5252–5257 (2009).
Koch, A. W., Pokutta, S., Lustig, A. & Engel, J. Calcium binding and homoassociation of E-cadherin domains. Biochemistry 36, 7697–7705 (1997).
Cheung, E. L. & Corey, D. P. Ca2+ changes the force sensitivity of the hair-cell transduction channel. Biophys J 90, 124–139 (2006).
Corey, D. P. & Hudspeth, A. J. Response latency of vertebrate hair cells. Biophys J 26, 499–506 (1979).
Corey, D. P. & Hudspeth, A. J. Kinetics of the receptor current in bullfrog saccular hair cells. J Neurosci 3, 962–976 (1983).
Rhode, W. S. & Geisler, C. D. Model of the displacement between opposing points on the tectorial membrane and reticular lamina. J Acoust Soc Am 42, 185–190 (1967).
Jin, X. et al. Crystal structures of Drosophila N-cadherin ectodomain regions reveal a widely used class of Ca(2)+-free interdomain linkers. Proc Natl Acad Sci USA 109, E127–134 (2012).
Miroux, B. & Walker, J. E. Over-production of proteins in Escherichia coli: mutant hosts that allow synthesis of some membrane proteins and globular proteins at high levels. J Mol Biol 260, 289–298 (1996).
Bers, D. M., Patton, C. W. & Nuccitelli, R. A practical guide to the preparation of Ca2+ buffers. Methods Cell Biol 40, 3–29 (1994).
Florin, E.-L. et al. Sensing specific molecular interactions with the atomic force microscope. Biosensors & Bioelectronics 10, 895–901 (1995).
Bustamante, C., Marko, J. F., Siggia, E. D. & Smith, S. Entropic elasticity of lambda-phage DNA. Science 265, 1599–1600 (1994).
Marko, J. F. & Siggia, E. D. Statistical mechanics of supercoiled DNA. Phys Rev E Stat Phys Plasmas Fluids Relat Interdiscip Topics 52, 2912–2938 (1995).
Valbuena, A. et al. Quasi-simultaneous imaging/pulling analysis of single polyprotein molecules by atomic force microscopy. Rev Sci Instrum 78, 113707 (2007).
Böhm, G., Muhr, R. & Jaenicke, R. Quantitative analysis of protein far UV circular dichroism spectra by neural networks. Protein Engineering 5, 191–195 (1992).
Humphrey, W., Dalke, A. & Schulten, K. VMD: Visual Molecular Dynamics. Journal Molecular Graphics 14, 33–38 (1996).
Acknowledgements
We thank Prof. Ulrich Müller (Johns Hopkins School of Medicine) for the mouse clones and protein samples of CDH23 and PCDH15 as well as Dr. E. Tejera (Instituto Cajal) for preparing the CDH23 and PCDH15 proteins, Dr. A. Gómez-Sicilia for assistance with AFM experiments, and Dr. Douglas Laurents for critically reading the manuscript. This work was supported by the BIO2010-22275 grant from the Spanish Ministry of Science and Innovation (MICINN) to M.C.-V.
Author information
Authors and Affiliations
Contributions
M.C.-V. conceived, designed and supervised the project; J.O. designed the project, conducted protein preparation and AFM data acquisition and analysis; A.G.-P. and D.F.B. performed AFM experiments; R.H. performed CD experiments; A.V. performed AFM experiments and acquired images by AFM; J.O. and M.C.-V. wrote the paper.
Corresponding authors
Ethics declarations
Competing Interests
The authors declare no competing interests.
Additional information
Publisher’s note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Oroz, J., Galera-Prat, A., Hervás, R. et al. Nanomechanics of tip-link cadherins. Sci Rep 9, 13306 (2019). https://doi.org/10.1038/s41598-019-49518-x
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-019-49518-x
This article is cited by
-
Emergence of slip-ideal-slip behavior in tip-links serve as force filters of sound in hearing
Nature Communications (2024)
-
Anisotropy in mechanical unfolding of protein upon partner-assisted pulling and handle-assisted pulling
Communications Biology (2021)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
