
Abstract
For 107 endemic mammal species in the Afro-Arabian region, Sahara-Sahel and Arabian Desert, we used ensemble species distribution models to: (1) identify the hotspot areas for conservation, (2) assess the potential impact of the projected climate change on the distribution of the focal species, and (3) assign IUCN threat categories for the focal species according to the predicted changes in their potential distribution range. We identified two main hotspot areas for endemic mammals: the Sinai and its surrounding coastal area in the East, and the Mediterranean Coast around Morocco in the West. Alarmingly, our results indicate that about 17% of the endemic mammals in the Afro-Arabian region under the current climate change scenarios could go extinct before 2050. Overall, a substantial number of the endemic species will change from the IUCN threat category “Least Concern” to “Critically Endangered” or “Extinct” in the coming decades. Accordingly, we call for implementing an urgent proactive conservation action for these endemic species, particularly those that face a high risk of extinction in the next few years. The results of our study provide conservation managers and practitioners with the required information for implementing an effective conservation plan to protect the biodiversity of the Afro-Arabian region.
Similar content being viewed by others
Introduction
The unprecedented loss of biodiversity, with an estimated extinction of a species every 20 minutes1, indicates that a “sixth mass extinction” is under way1,2,3,4. There is now overwhelming evidence that both climate change and habitat fragmentation are together responsible for this exceptional biodiversity loss5, with further accelerated extinction predicted during the next decades2,5. Unless we implement effective conservation measures immediately about two-thirds of the vertebrate populations could vanish by 20203. Accordingly, the Convention on Biological Diversity listed strategic goals to forestall the ongoing loss of biodiversity6. Assessing the current and future biodiversity status and future trends are imperative to achieve these goals7,8.
Previous empirical studies show that species’ ecological traits influence their response to climate change and that response varies across species9,10. Traits that may increase risk of extinction because of climate change include: (1) narrow distribution ranges; (2) limited dispersal ability; (3) high level of specialisation, and (4) low reproductive performance11. Narrow-ranged species, tend to possess most of these traits, and are therefore more vulnerable to climate change than other species11,12. Accordingly, assessing the potential impact of climate change on biodiversity patterns and species distribution is essential to determine the most vulnerable species and areas, which would allow for prioritising conservation efforts. Several assessments at global and continental scales12,13,14,15,16 have been carried out; however, effective conservation plans require information at the more local scale of biomes or ecoregions17,18.
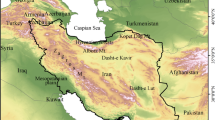
The warm desert biome has been neglected in conservation ecology2,8,19. Yet, the warm desert biome encompassing the Sahara and the Arabian Desert (Fig. 1; henceforth referred to as the “Afro-Arabian” region) harbours a unique mix of fauna and flora shaped by the harsh climatic conditions and the long-term geographical connectivity between Africa and Asia20. Therefore, many of the Afro-Arabian species are endemic and uniquely adapted to extreme environmental conditions2,8,19,21.

The geographic location of the Afro-Arabian region.
Climatic conditions limit mammal distribution by influencing physiological processes and vegetation conditions, which in turn determine resource availability22. The mammal fauna of the Afro-Arabian region, and in particular the endemic species, are among the most adapted to the extreme and harsh environmental conditions8,23. However, evidence suggests that the genetic adaptation in mammals can not keep pace with the current rate of climate change24 making living at the edge of the physiological limits particularly risky24,25. Mammals, as consumers, are at higher trophic levels, and have a unique functional role in the ecosystem2; therefore, focusing our attention on them could be a useful surrogate for greater biodiversity conservation and a suitable way to prioritise the conservation effort22.
The long history of conflict, political instability, and perception as a bare area with comparably low biomass, are responsible for the lack of information regarding the biodiversity patterns and species distributions in the Afro-Arabian region2,8,26. In addition, despite their important functional roles in the ecosystem, these species have also been understudied because of the difficulties associated with studying the often nocturnal, rare and secretive species1,2. Such lack of information represents the main challenge for any conservation action.
The main goal of this study, which also forms part of a published PhD thesis27, is to identify the conservation status and diversity patterns of the endemic mammal species in the Afro-Arabian region based on species distribution models (SDMs). Species distribution models allow for delineating species geographic and environmental niches through quantifying the relationships between their occurrences and associated biotic and abiotic variables, and allow for predicting species niches in unsampled regions28,29. SDMs are powerful tools and widely used in many fields (e.g., biogeography, ecology, conservation, and epidemiology) and can project species niche spatially and temporally30. Specifically, we aim to address the following questions:
-
Where are the biodiversity hotspot areas located?
-
What is the expected impact of climate change on biodiversity in the Afro-Arabian region?
-
What would the conservation status of focal species be under climate change?
We delineated the contemporary potential distribution ranges for the endemic mammal species of the Afro-Arabian region using SDMs. Then, based on the range maps derived from the SDMs, we defined the biodiversity hotspot areas in the study area. We then projected the current potential ranges temporally into 2050 and 2070 to predict the potential impact of climate change on the distribution of the endemic mammals in the future. According to the degree of predicted change in the species’ ranges, we then assigned a threat category to each species following the IUCN Red List criterion A3(c).
Results
We compiled data for 107 endemic mammal species belonging to 17 families (Supplementary Table S1). The most represented families were Muridae (41 species), Soricidae (18 species), and Bovidae (16 species). All ensemble SDMs had a good predictive performance (True Skill Statistic (TSS) > 0.49, mean = 0.80, SD = 0.09, and Area Under the Receiver Operator Curve (AUC) > 0.74, mean = 0.89, SD = 0.05) suggesting that the SDMs were well capable to identify suitable areas within the study region (AUC & TSS values for each species are listed in Supplementary Table S2). The spatial analysis revealed that the contemporary species richness reaches a maximum of 35 endemic mammal species per ~25 km2. The areas with the highest richness values were concentrated along the coast of the Mediterranean and Red sea, and around the mountainous regions. The areas with the lowest richness were found in central Algeria and Mauritania in the West and central Saudi Arabia in the East (Fig. 2). It is noteworthy that the species richness showed a latitudinal gradient for endemic mammal species increasing northward.

Species richness areas of the endemic mammal species in the Afro-Arabian region along three time slices. The “Current” refers to the contemporary status of the species richness, 2050 and 2070 refer to the predicted status in those years. The scale bar shows the score of the species richness, where zero means almost no species and 30 means the number of species in the pixel.
Our results also revealed that both species richness and turnover showed significant temporal trends, where the projected change in climate conditions resulted in markedly reduced mammalian diversity in the study area. We found, under the Unlimited Dispersal scenario (UD) assuming that the species will be able to disperse freely to the projected suitable habitats, about 17% reduction in the species richness by 2050, increasing to 20% by 2070. Similarly, species turnover—the change in species composition over time— was predicted to increase steadily during the 21st century (Fig. 3). The changes in species turnover were predicted to be higher particularly in the central Sahara and the western side of the Arabian Desert.

Temporal species turnover of the endemic mammal species in the Afro-Arabian region. The zero value indicates no change in species composition over time (2050 and 2070) and the value of one indicates a complete change in species composition.
The impact of the projected climate change scenarios on the potential species distribution was not consistent and varied among the species, where some were predicted to reduce the size of their distribution ranges and become threatened, while other species were predicted to expand their distribution ranges and thus benefit from climate change. According to the No Dispersal scenario (ND), assuming that the species will not be able to occupy the projected suitable habitats, 17 and 22 species were predicted to go extinct (losing 100% of their suitable range) by 2050 and 2070 respectively. Under the same scenario, 84 and 87 species were predicted to lose more than 30% of their suitable ranges by 2050 and 2070 respectively (Fig. 4 and Supplementary Table S3). In the unlimited dispersal scenario, the models predicted 15 and 22 species to go extinct by 2050 and 2070 respectively, while 15 species were predicted to expand their distribution ranges over that time (Fig. 4). The majority of the species at risk of going extinct were small mammals belonging to the family Soricidae (seven out of 21) and the family Muridae (ten out of 42). In general, about 80 endemic mammal species in the Afro-Arabian region will be likely to become severely threatened (losing 50–100% of their current potential suitable habitats) according to the projected climate change (Fig. 4).

The percentage of change in climatically-suitable habitat for the endemic mammal species in 2050 (left) and 2070 (right) inhabiting the Afro-Arabian region. The negative numbers on the x-axis indicate loss in suitable habitat. The numbers on the bars represent the number of species in each category.
Our results show a striking difference between the current IUCN conservation status and the predicted status according to the projected change in the distribution range (Fig. 5). About 50% of the species currently classified as “Least Concern” are expected to be classified in the near future anywhere between the categories “Vulnerable” to even “Extinct” (according to the Unlimited Dispersal scenario).

Changes in the threat category for the endemic mammal species in the Afro-Arabian region over time. The upper plot represents the unlimited dispersal scenario (UD), while the lower plot represents the no dispersal scenario (ND). The x-axis represents the temporal scale, where the current refers to the current species conservation status according to the IUCN, while 2050 and 2070 are the predicted conservation status according to the change in distribution range in those respective years.
Discussion
Based on the best available species distribution models, we examined how the projected climate change could influence the distribution and diversity patterns of endemic mammals in the Afro-Arabian region. Our results show that many of the endemic mammals are facing an extremely augmented risk of extinction in the immediate future (Fig. 4 and Supplementary Table S3). This result supports the findings of previous studies that have shown that a substantial number of vertebrate species globally are expected to become extinct in the next decade3,31. This high extinction risk could be amplified by the influence of additional drivers such as changing land-use patterns, disease outbreaks, human pressure, and introduction of novel parasites or invasive species32,33. In fact, the impact of these drivers is already obvious in the Afro-Arabian region, in particular catalysed by the absence of law enforcement32. Additionally, changing rainfall patterns due to climate change can increase a species’ susceptibility to disease outbreaks, exemplified by the population decline in the Kruger park as a result of an Anthrax outbreak after the 1990 drough22.
In contrast, species ecological traits could also dampen the impact of climate change. Some species may show enough phenotypic or behavioural plasticity to adapt to new climate condition34. For instance, studies showed that the bushy-tailed woodrat, Neotoma cinerea, increased in body mass to adapt to low temperatures in the Late Quaternary, and decreased it to adapt to the recent increases in temperature34,35,36. For some species, particularly small mammals, avoiding high temperatures can be achieved by finding shelter such as burrows22. Other species can avoid the effects of rising temperature by employing “adaptive heterothermy” a physiological mechanism to reduce heat gain and evaporative heat loss and thus conserving body water25,37. Accordingly, if species succeed to adapt to local conditions, the impact of climate change may not be as pronounced as projected22.
For those species predicted to expand their climatically suitable ranges (Fig. 4), it will be important to have a dispersal rate that conforms with the climatic velocity (i.e. the rate at which these species disperse should keep pace with the change in climate)38,39,40. In the Afro-Arabian region, which is dominated by high climatic velocity, the species dispersal ability is either naturally low or artificially limited by habitat fragmentation. Furthermore, biotic interactions and the existence of natural and anthropogenic barriers further limit the species ability to expand or shift their distribution ranges5,38,39. Assuming that species would be able to disperse to the new climatically-suitable habitats, the species still may not be able to colonise the new habitats if it is not meeting the requirements34. The ability of these species to migrate, especially herbivorous mammals, is limited by the mobility of the plant species required as food34. Whereas particularly the predators are intimately limited by prey availability rather than just dispersal capacities to the new generally climatically-suitable habitats22.
Our results show a high divergence between the current IUCN conservation status and the predicted status according to the projected change in the distribution range (Fig. 5). This disagreement is owed to applying different criteria to assess species extinction. For instance, our assessment is based on the A3(c) criterion that assign species to a threat category according to the degree of change in species’ ranges. Other criteria use other factors (e.g., the size of distribution range, population status, number of sub-population, and threats severity) to assign species to a threat category. As such, species can be considered threatened according to a specific criterion and not threatened according to another. We acknowledge that SDMs do not identify the proximate causes of species extinction12, however, the reduction in species distribution range is very likely to increase the risk of local extension41. For instance, species predicted to be or become range restricted to a few locations, could easily go extinct by any further local threat such as land transformation or disease outbreaks12. In any case, our findings, together with the results of previous studies, advocate for considering the impact of projected climate change in future assessments calling for immediate action12,22,34,42.
The latitudinal gradient in the current species richness corroborates the pattern for other taxa43,44,45,46. This pattern could be attributed to factors such as palaeogeography and/or historical land-use patterns which complemented each other to form a mosaic of interconnected habitats, resulting in high species richness in the northern parts of the study area47,48. Additionally, areas with high species richness are also geographic transition zones. For instance, the Sinai is a transitional habitat between Asia and Africa, while Morocco lies between Africa and Europe. These transition habitats are characterised by high levels of species diversity and endemism, and the occurrence of species with allopatric ranges49.
Although species richness was predicted to decrease and hotspot areas to become isolated during the next decades, the regions with high richness were predicted to remain around the coastal areas and in the Sinai region (Fig. 3). Previous studies also revealed the importance of these regions for many other taxa at global and local scales32,44,45,50,51,52,53,54. However, the political conflicts, particularly in the Sinai region, and the sprawling development along the coastal areas do not hold much promise for the survival of species in these regions. An accurate assessment of biodiversity and its threats is not available across the Afro-Arabian region because of its remoteness and political instability. However, the rate of hunting wildlife unsustainably substantially increased and several wildlife massacres have been reported particularly after the “Arab spring” in 201126,55,56. Recent analyses show that the increased conflict and anthropogenic activities resulted in a dramatic population decline for many species55,57,58. Additionally, most of the megafauna in the southern part of the study area have been almost extinguished because of armed conflicts55. The current conflicts and instability together with other disturbances (mining, grazing, agriculture, and urbanisation), leave endemic species with nowhere to go, amplifying the risk of extinction to be realistically much more dramatic than our projections2,55,58.
It is noteworthy that the regions with high species turnover corresponded to the regions with low species richness, except in the Sinai region (Fig. 3). This could be because the species assemblages in our study follow the hollow curve pattern, where most species have small distribution ranges with just a few large ones59,60,61. The change in species composition reflected by high species turnover, due to species range contractions and expansions, as well as influx of new species and/or departure of other species, could negatively influence the existing species interaction patterns and the trophic network structure22,34,42.
Our results should be interpreted cautiously, given some of the inherent uncertainties associated with SDMs being projected into the future. A primary source of uncertainty in our study is the quality and quantity of species occurrences. In this study, the distribution ranges for about 20% of the focal species were modelled with low quantities of five to 10 occurrences. Using such small numbers for modelling is challenging and likely to reduce prediction accuracy. However, we minimised model uncertainty by avoiding model complexity using fewer predictors (i.e. equal to or less than the number of occurrences) which should reduce the risk of model over-fitting62. Additionally, we used SDM algorithms that proved to have good performance with low numbers of occurrences63. Since our data was collected from different sources with different sampling approaches, it certainly contained sampling bias, which often results in environmental bias (i.e. over‐representation of certain environments) and, in turn, results in truncated response curves and ultimately yields less reliable predictions64,65. We used, however, a spatial filtering approach to address such potential bias in the species occurrence data. Another sources of uncertainty comes from model extrapolation, projecting models onto non-analogue environments that were not used for calibration62,66. To minimise model extrapolation bias and improve the prediction reliability, we projected the current distributions onto the future climate scenarios using the ‘clamping’ option, which allows some extrapolation onto non-analogue environments, but in a constrained manner. We acknowledge that there is also another more conservative approach, which completely avoids model extrapolation, however, such an approach can produce truncated response curves and even less reliable projections67. We are not claiming that our projections have no extrapolation bias (they certainly have), however, this extrapolation bias is restricted and reasonable from a practical stand point. A further source of uncertainty is variable selection and availability. Information, for example, on how vegetation variables (VCF) might change in the future is not available, accordingly we assumed it to remain (i.e. static variable) unchanged. It is therefore crucial to understand that our projections are only valid under the assumption we had to make. However, since most of the species with distribution ranges shaped by VCF were bats, which have the ability to disperse and track changes in VCF that could happen in the future, we do not expect that changing VCF in the future will significantly change the conclusions of our study.
Conservation Implication
For systematic conservation planning and setting conservation priorities it is imperative to know about species distributions and their relationship with biotic and abiotic factors68,69,70. However, despite their ecological roles in the ecosystem, particularity in the Sahara and the Arabian Desert, mammal species are widely neglected from conservation plans, and not even fully recognised by the local governments2,19,27. The ecological role that many mammal species play makes them ideal umbrella or keystone species. Prioritising the conservation efforts towards these species would provide collateral protection to co-occurring species22. For instance, the recruitment of Acacia species, a keystone species in the Sahara and the Arabian Desert, increased with the presence of herbivore species (e.g. Dorcas gazelle Gazella dorcas)71. Unlike other herbivorous species, Dorcas gazelles are known to be browsers and not grazers, which, in turn, allows the vegetation cover for the desert ecosystem to survive and persist, in addition they also act as plant seeds dispersers72,73,74. At the same time, the gazelles are the main prey for several predators (e.g. Cheetah Acinonyx jubatus and Wolf Canis lupus) that have suffered from decline in prey availability75.
Our results, together with other studies5,10,26 all show that biodiversity in the Afro-Arabian region is highly vulnerable to projected climate change. To save what is left, conservation measures should prioritise the hotspot areas, i.e. regions with a high number of endemic species. Vanishing species and populations will potentially affect the existing ecological network as some species may experience ecological release and others may be suppressed causing cascades of extinctions42,76. The Afro-Arabian species exhibit high physiological plasticity and distinct evolutionary histories that provide valuable insights into the genetic basis of species tolerance to environmental extremes2,4,5. Thus, losing these endemic species could lead to a loss in the pool of important genetic information that is vital for understanding species’ evolution and survival in extreme environments4. As such, conservation efforts need to maximise species dispersal within their suitable habitat; this would slow down the rate of extinction by maintaining the interaction between species and populations and preserving the genetic network77. We highly recommend applying adaptive conservation strategies inside and outside protected areas as a first critical step forward32,78. These strategies include three options: 1) the resistance option, reducing the threat impacts and conserving valued resources (e.g., reducing non-climatic stress—invasive species and habitats fragmentation— and promoting green energy), 2) the resilience option, restoring the ecosystem functions (e.g., improving protected area networks and habitats connectivity), and 3) the response option, allowing the ecosystem transition to new conditions (e.g., using transboundary protected areas)32,78. The existing protected area networks can be used to initiate the adaptive conservation strategies. Increasing redundancy in the existing protected area networks can provide species with more opportunities to move and disperse, which, ultimately, improves species resilience to climate change79. The existing protected area networks in the Afro-Arabian region are few, small in size, and spatially disconnected32. Therefore, expanding the protected area networks is likely to preserve various environmental conditions, improve habitat connectivity, and facilitate species dispersal along different environmental gradients77. Additionally, we recommend establishing new protected areas to cover the regions predicted to experience high turnover rates (Fig. 3), and managing the surrounding habitats in ways that allow for species dispersal and movement. The socio-economic aspect and community-based conservation approach need to be considered in the conservation management plan to achieve a long-term conservation success32.
Methods
Study area
This study focuses on the Afro-Arabian region which covers ~17,820,000 km2, and extends over 26 countries in Africa and Asia (Fig. 1). It covers two deserts, the Saharan and the Arabian Desert, that occupy the southern part of the western Palearctic and extend also into the Afro-tropical realm27. The region is bordered by the Mediterranean Sea in the north, Persian Gulf in the east, the Atlantic Ocean in the west, and the political borders of the Sahel countries in the south (Fig. 1). This region has relatively high topographic heterogeneity (altitude varies from ~−400 to 4000 meters a.s.l.) and habitat diversity (sand dunes, coastal plains, mountains). It encompasses mainly three distinct climatic regions: warm Mediterranean, warm semi-arid, and warm desert80. The latter climate region encompasses the Sahara biome, the largest warm desert in the world, with mean annual temperature of ~32 °C and mean annual precipitation of zero26, and spreading over ~80% of the study region. The study region experienced environmental and geological oscillations in the past that allowed for faunal interchange between Africa and Asia20.
Species data
Point occurrences for 107 mammal species (Supplementary Table S1), specifically 95 endemic and 12 near-endemic species (>70% of their distribution ranges within the study region), were assembled from different resources, including personal fieldwork, published and “gray” literature, online portals, and personal communications (Supplementary Table S4). We updated species taxonomy following the Mammal Species of the World portal81. We restricted the species records to those only collected from the wild between 1960 and 2005, and omitted the duplicated occurrences within one grid cell (~5 × 5 km). To correct for sampling bias, we applied a spatial filtering approach82,83. Specifically, we filtered records of each species to obtain the highest achievable occurrences that are 5, 10, and 20 km apart for small, volant, and large mammals respectively. We chose these distances to represent the mean dispersal ability of each group. This should lead to less spatially autocorrelated records with less environmental bias, and yield better prediction for species distribution ranges84. We acknowledge that there are several bias correction approaches such as calibrating the model with extra predictors representing site accessibility or sampling effort, or background manipulation85,86. However, in this study we favoured spatial filtering since it has been shown to outperform other bias correcting approaches87,88,89,90. As a consequence, it has been widely applied recently to correct the spatial and environmental bias in species data83,90,91,92,93,94. Spatial filtering outweighs the negative influence of the biased data and substantially minimises omission errors (i.e. suitable areas not predicted by models) and commission errors (i.e. areas mistakenly predicted to be suitable) particularly in areas with limited surveying effort87. (For assessing the environmental bias see Supplementary Methods “Environmental sampling bias assessment” & Supplementary Figure S1).
Environmental data
We initially selected 26 environmental predictors, 19 contemporary climatic predictors available from the WorldClim database and seven non-climatic predictors characterising the topography and vegetation, at a resolution of 2.5 arc-min (Supplementary Table S5), to model species distributions. This resolution allows for mitigating the uncertainty that might result from the positional uncertainty associated with the species occurrences, and also for avoiding the issue of not identifying local refugia, a common issue in coarse spatial resolution climate change projections found on correlative SDMs32,95. Model over-fitting is a problem in the presence of multi-collinearity between the predictors96,97. We therefore calculated the collinearity between the predictors based on the Variance Inflation Factor (VIF), and removed the predictors with VIF values greater than 1098, using the “vifstep” function in “usdm” R package99. This reduced the number of predictors to 13 (Supplementary Tables S5 & S6). To assess the potential impact of climate change on the distribution of the focal species, we retrieved projections of future climate variables from four general circulation models (GCMs), HadGEM2-AO, CCSM4, IPSL-CM5A-LR, and ACCESS1-0, downscaled to 2.5 arc-min resolution from the IPCC Fifth Assessment, available from the WorldClim database for the years 2050 and 2070100. We used the VIF to assess collinearity within future predictors (2050 and 2070) and to avoid making predictions based on collinearity structures in time101 (Supplementary Table S6).
We used the RCP 8.5 pessimistic emission scenario that assumes increasing in the global population and slow per capita economic growth, leading to high energy demand and high GHG emissions and the average temperature over time in absence of climate change policies102,103. The Afro-Arabian region, in many of its countries, is suffering from civil conflicts and wars, political unrest, increasing population size, low per capita income, and absence of climate change policies55,56. Therefore, we restricted our analysis to the RCP 8.5 scenario as most realistic scenario (or even the most pessimistic), for the Afro-Arabian region, that is probably representing the actual situation in the study area and also bears only the risk of our results being a conservative outcome. Finally, since there is no data on the ‘vegetation continuous field’ (VCF) variable104 and how it might change in the future, we assumed that VCF remains constant to be able to include it in the future predictions105,106. To avoid prediction unreliability owing to non-analogue environments in the future (2050 and 2070), we used ExDet tools107 to identify the presence of non-analogue environments and the extent of model extrapolation (see Supplementary Methods “non-analogue environments and extrapolation” & Supplementary Figures S2 and S3).
Species Distribution Models (SDMs)
We adopted ensemble SDMs with the weighted average consensus approach that accounts for model-based uncertainty108,109 to delineate the contemporary and future distribution ranges. The ensembles consisted of four different SDM algorithms: MaxEnt110, Random Forest (RF)111, Generalized Boosted Model (GBM)112, and Generalized Linear Model (GLM)113, available within the “biomod2” R package114. However, for each species, the selection of SDM algorithms was based on the number of species occurrences and species specialisation following the framework developed by Soultan and Safi27,63. To minimise the model complexity and avoid model over-fitting82,83, we calibrated an initial model for each species using the filtered species occurrences and the uncorrelated current environmental predictors. Subsequently, for each species, we calculated the variable importance of each predictor to model gain, retaining those with a relative contribution greater than 5% for the final model calibration (Supplementary Table S7)115,116,117. To avoid geographical over-prediction, we used a species-specific study area118. Specifically, we restricted the model calibration to the buffered minimum convex polygon (MCP) of the species occurrences, where the buffer distance was equal to the mean distance between the occurrence points of the focal species83,84,119. This was done to avoid model over-fitting when the environmental conditions in species occurrences were different from those in background locations120,121,122. We thus randomly sampled 10’000 background points, constrained to within the buffered area, to characterise the environmental background122.
Finally, we projected the model predictions onto the Afro-Arabian region to determine the contemporary potential distribution for each species. For species with a large MCP (>50% of the study area), we used the entire study area for model calibration. We used spatial cross-validation to avoid calibrating and evaluating based on the same data50,83. To this end, we segregated species localities geographically into four blocks, then, models were built in an iterative manner, using three blocks for calibration and the remaining fourth block for evaluation in each iteration, until all blocks have been used once for evaluation123,124. This allowed for: 1) maintaining spatial independence between training and test data, 2) avoiding AUC inflation due to spatial auto-correlation between training and test data and, 3) incorporating the uncertainty in the estimated probability123,124. We evaluated the predictive performance of the models by assessing their ability to predict relative likelihoods of species occurrence within the test data blocks, using the area under the curve (AUC) of the receiver operating characteristic (ROC) plot125, as well as the True Skill Statistic (TSS)126. We projected the contemporary species distribution ranges into the future (2050 and 2070) in order to assess the possible consequences of climate changes on the biodiversity in the Afro-Arabian region. There is large variability among general circulation models (GCMs), which can cause uncertainties in species distribution projections123,127,128,129. Accordingly, to reduce the bias in certain areas from one GCM, we calculated the median of the predictions from the four GCMs129,130,131,132. This approach showed to provide accurate and less biased predictions and provides better consideration for the uncertainties in future species distribution129,131,133.
Spatial analysis
We quantified the impact of global change on the spatial biodiversity patterns through two indices: species richness and species turnover. Species richness is important for understanding the ecological and evolutionary processes that shape the biodiversity in a given location and is one of essential measures for prioritising conservation efforts8,134. We estimated species richness for current and future, 2050 and 2070, by summing up the predicted relative likelihoods of species occurrence for all species for the corresponding time34,124. Species turnover is a measure of dissimilarity in the composition of species assemblages from site to site within a given time frame or over time within a given site135. We used the Bray-Curtis index to quantify the “temporal” species turnover, dissimilarity for each pixel between the current and future, using the “vegdist” function in the “vegan” R package136.
Additionally, we accounted for the ability of species to track the changes in climate using two dispersal scenarios: Unlimited Dispersal (UD), assuming that the species can disperse freely and adjust their distribution range according to climate, and No Dispersal (ND), where species are assumed to be unable to disperse and, as a consequence, will not be able to occupy the projected suitable habitats. According to the rate of change in species distribution ranges, we assigned each species to a threat category following the International Union for Conservation of Nature (IUCN) threat categories137. We applied the following criteria as described in IUCN Red List criterion A3(c): Extinct when a species is predicted to lose 100% of its current potential range; Critically Endangered when a projected range loss is >80%; Endangered projects a range loss of >50%; and Vulnerable when a projected range loss is >30%9,12,34. IUCN Red List criterion A3(c) estimate the rate of geographic range decline over a period of 10 year or three generations, whichever is the longer138. Since 10 years is a short period to measure the impact of climate change and also does not reflect time scales for human interventions138, we used the generation definition and considered each time period (2050 and 2070) as equal to three generations.
References
Levin, P. S. & Levin, D. A. Macroscope: The Real Biodiversity Crisis. Am. Sci. 90, 6–8 (2002).
Durant, S. M. et al. Fiddling in biodiversity hotspots while deserts burn? Collapse of the Sahara’s megafauna. Divers. Distrib. 20, 114–122 (2014).
WWF.. Living Planet Report 2016. Risk and resilience in a new era. (WWF International, Gland, Switzerland, 2016).
Ceballos, G., Ehrlich, P. R. & Dirzo, R. Biological annihilation via the ongoing sixth mass extinction signaled by vertebrate population losses and declines. Proc. Natl. Acad. Sci. USA 114, E6089–E6096 (2017).
Vale, C. G. & Brito, J. C. Desert-adapted species are vulnerable to climate change: Insights from the warmest region on Earth. Glob. Ecol. Conserv. 4, 369–379 (2015).
CBD. Strategic Plan for Biodiversity 2011–2020. Further information related to the technical rationale for the Aichi Biodiversity Targets, including potential indicators and milestones. UNEP/CBD/COP/10/INF/12/Rev.1. Convention on Biological Diversity (2010).
Visconti, P. et al. Projecting Global Biodiversity Indicators under Future Development Scenarios. Conserv. Lett. 9, 5–13 (2016).
Vale, C. G. et al. Biogeography and conservation of mammals from the West Sahara-Sahel: an application of ecological niche-based models and GIS. Hystrix, Ital. J. Mammal. 27, 1–10 (2016).
Thuiller, W. et al. Consequences of climate change on the tree of life in Europe. Nature 470, 531–534 (2011).
Sow, A. S., Martínez-Freiría, F., Dieng, H., Fahd, S. & Brito, J. C. Biogeographical analysis of the Atlantic Sahara reptiles: Environmental correlates of species distribution and vulnerability to climate change. J. Arid Environ. 109, 65–73 (2014).
Gibson, L., McNeill, A., Tores, Pde, Wayne, A. & Yates, C. Will future climate change threaten a range restricted endemic species, the quokka (Setonix brachyurus), in south west Australia? Biol. Conserv. 143, 2453–2461 (2010).
Thuiller, W., Lavorel, S., Araújo, M. B., Sykes, M. T. & Prentice, I. C. Climate change threats to plant diversity in Europe. Proc. Natl. Acad. Sci. USA 102, 8245–8250 (2005).
Rondinini, C. et al. Global habitat suitability models of terrestrial mammals. Philos. Trans. R. Soc. London B Biol. Sci. 366, 2633–2641 (2011).
Boitani, L. et al. Distribution of medium- to large-sized African mammals based on habitat suitability models. Biodivers. Conserv. 17, 605–621 (2008).
Record, S., Charney, N. D., Zakaria, R. M. & Ellison, A. M. Projecting global mangrove species and community distributions under climate change. Ecosphere 4, art34 (2013).
Ceballos, G. & Brown, J. H. Global Patterns of Mammalian Diversity, Endemism, and Endangerment. Conserv. Biol. 9, 559–568 (1995).
Christianini, A. V., Oliveira, P. S., Bruna, E. M. & Vasconcelos, H. L. Fauna in decline: Meek shall inherit. Science. 345, 1129–1129 (2014).
Moura, M. R., Argôlo, A. J. & Costa, H. C. Historical and contemporary correlates of snake biogeographical subregions in the Atlantic Forest hotspot. J. Biogeogr. 44, 640–650 (2017).
Durant, S. M. et al. Forgotten Biodiversity in Desert Ecosystems. Science. 336, 1379–1380 (2012).
Metallinou, M. et al. Conquering the Sahara and Arabian deserts: systematics and biogeography of Stenodactylus geckos (Reptilia: Gekkonidae). BMC Evol. Biol. 12, 258 (2012).
Brito, J. C. et al. Conservation Biogeography of the Sahara-Sahel: additional protected areas are needed to secure unique biodiversity. Divers. Distrib. 22, 371–384 (2016).
Thuiller, W. et al. Vulnerability of African mammals to anthropogenic climate change under conservative land transformation assumptions. Glob. Chang. Biol. 12, 424–440 (2006).
Ward, D. Biodiversity and Biogeography of Deserts. In The Biology of Deserts 192–216, https://doi.org/10.1093/acprof:oso/9780198732754.003.0009 (Oxford University Press, 2009).
Hetem, R. S., Fuller, A., Maloney, S. K. & Mitchell, D. Responses of large mammals to climate change. Temp. (Austin, Tex.) 1, 115–27 (2014).
Hetem, R. S. et al. Variation in the daily rhythm of body temperature of free-living Arabian oryx (Oryx leucoryx): does water limitation drive heterothermy? J. Comp. Physiol. B 180, 1111–1119 (2010).
Brito, J. C. et al. Unravelling biodiversity, evolution and threats to conservation in the Sahara-Sahel. Biol. Rev. Camb. Philos. Soc. 89, 215–231 (2014).
Soultan, A. Biogeography and conservation of the neglected biodiversity. (University of Konstanz, 2018).
Carvalho, S. B., Brito, J. C., Pressey, R. L., Crespo, E. & Possingham, H. P. Simulating the effects of using different types of species distribution data in reserve selection. Biol. Conserv. 143, 426–438 (2010).
Guisan, A. et al. Predicting species distributions for conservation decisions. Ecol. Lett. 16, 1424–35 (2013).
Elith, J. & Leathwick, J. R. Species Distribution Models: Ecological Explanation and Prediction Across Space and Time. Annu. Rev. Ecol. Evol. Syst. 40, 677–697 (2009).
Ripple, W. J. et al. Collapse of the world’s largest herbivores. Sci. Adv. 1, e1400103 (2015).
Maiorano, L. et al. The future of terrestrial mammals in the Mediterranean basin under climate change. Philos. Trans. R. Soc. B Biol. Sci. 366, 2681–2692 (2011).
Hoffmann, M. et al. The changing fates of the world’s mammals. Philos. Trans. R. Soc. B Biol. Sci. 366, 2598–2610 (2011).
Levinsky, I., Skov, F., Svenning, J.-C. & Rahbek, C. Potential impacts of climate change on the distributions and diversity patterns of European mammals. Biodivers. Conserv. 16, 3803–3816 (2007).
Smith, F. A., Browning, H. & Shepherd, U. L. The influence of climate change on the body mass of woodrats Neotoma in an arid region of New Mexico, USA. Ecography. 21, 140–148 (1998).
Smith, F. A. & Betancourt, J. L. Response of Bushy-Tailed Woodrats (Neotoma cinerea) to Late Quaternary Climatic Change in the Colorado Plateau. Quat. Res. 50, 1–11 (1998).
Schmidt-Nielsen, K., Schmidt-Nielsen, B., Jarnum, S. A. & Houpt, T. R. Body Temperature of the Camel and Its Relation to Water Economy. Am. J. Physiol. Content 188, 103–112 (1956).
Loarie, S. R. et al. The velocity of climate change. Nature 462, 1052–1055 (2009).
Carroll, C. et al. Biotic and Climatic Velocity Identify Contrasting Areas of Vulnerability to Climate Change. PLoS One 10, e0140486 (2015).
Menéndez, R., González-Megías, A., Lewis, O. T., Shaw, M. R. & Thomas, C. D. Escape from natural enemies during climate-driven range expansion: a case study. Ecol. Entomol. 33, 413–421 (2008).
Thomas, C. D. et al. Extinction risk from climate change. Nature 427, 145–148 (2004).
Velásquez-Tibatá, J., Salaman, P. & Graham, C. H. Effects of climate change on species distribution, community structure, and conservation of birds in protected areas in Colombia. Reg. Environ. Chang. 13, 235–248 (2013).
Patiny, S. & Michez, D. Biogeography of bees (Hymenoptera, Apoidea) in Sahara and the Arabian deserts. Insect Syst. Evol. 38, 19–34 (2007).
Patiny, S., Michez, D., Kuhlmann, M., Pauly, A. & Barbier, Y. Factors limiting the species richness of bees in Saharan Africa. Bull. Entomol. Res. 99, 337 (2009).
Bilgin, R., Keşişoğlu, A. & Rebelo, H. Distribution Patterns of Bats in the Eastern Mediterranean Region Through a Climate Change Perspective. Acta Chiropterologica 14, 425–437 (2012).
Myers, N., Mittermeier, R. A., Mittermeier, C. G., Da Fonseca, G. A. B. & Kent, J. Biodiversity hotspots for conservation priorities. Nature 403, 853–858 (2000).
Medail, F. & Quezel, P. Biodiversity Hotspots in the Mediterranean Basin: Setting Global Conservation Priorities. Conserv. Biol. 13, 1510–1513 (1999).
Covas, R. & Blondel, J. Biogeography and history of the Mediterranean bird fauna. Ibis (Lond. 1859). 140, 395–407 (2008).
Bilgin, R., Karataş, A., Çoraman, E., Disotell, T. & Morales, J. Regionally and climatically restricted patterns of distribution of genetic diversity in a migratory bat species, Miniopterus schreibersii (Chiroptera: Vespertilionidae). BMC Evol. Biol. 8, 209 (2008).
Tisseuil, C., Leprieur, F., Grenouillet, G., Vrac, M. & Lek, S. Projected impacts of climate change on spatio-temporal patterns of freshwater fish beta diversity: a deconstructing approach. Glob. Ecol. Biogeogr. 21, 1213–1222 (2012).
García Molinos, J. et al. Climate velocity and the future global redistribution of marine biodiversity. Nat. Clim. Chang. 6, 83–88 (2015).
Ficetola, G. F., Bonardi, A., Sindaco, R. & Padoa-Schioppa, E. Estimating patterns of reptile biodiversity in remote regions. J. Biogeogr. 40, 1202–1211 (2013).
Jones, M. C. & Cheung, W. W. L. Multi-model ensemble projections of climate change effects on global marine biodiversity. ICES J. Mar. Sci. 72, 741–752 (2015).
Leach, K., Zalat, S. & Gilbert, F. Egypt’s Protected Area network under future climate change. Biol. Conserv. 159, 490–500 (2013).
Brito, J. C. et al. Armed conflicts and wildlife decline: Challenges and recommendations for effective conservation policy in the Sahara-Sahel. Conserv. Lett. e12446, https://doi.org/10.1111/conl.12446 (2018).
Daskin, J. H. & Pringle, R. M. Warfare and wildlife declines in Africa’s protected areas. Nature, https://doi.org/10.1038/nature25194 (2018).
Duncan, C., Kretz, D., Wegmann, M., Rabeil, T. & Pettorelli, N. Oil in the Sahara: mapping anthropogenic threats to Saharan biodiversity from space. Philos. Trans. R. Soc. Lond. B. Biol. Sci. 369, 20130191 (2014).
Newby, J., Wacher, T., Durant, S. M., Pettorelli, N. & Gilbert, T. Desert Antelopes on the Brink: How Resilient is the Sahelo-Saharan Ecosystem? In Antelope Conservation: From Diagnosis to Action 253–279, https://doi.org/10.1002/9781118409572.ch13 (John Wiley & Sons, Ltd, 2016).
Ochoa-Ochoa, L. M., Rodríguez, P., Mora, F., Flores-Villela, O. & Whittaker, R. J. Climate change and amphibian diversity patterns in Mexico. Biol. Conserv. 150, 94–102 (2012).
McGill, B. J. et al. Species abundance distributions: moving beyond single prediction theories to integration within an ecological framework. Ecol. Lett. 10, 995–1015 (2007).
Krebs, C. The Ecological World View. (CSIRO Publishing, 2008).
Merow, C. et al. What do we gain from simplicity versus complexity in species distribution models? Ecography. 37, 1267–1281 (2014).
Soultan, A. & Safi, K. The interplay of various sources of noise on reliability of species distribution models hinges on ecological specialisation. PLoS One 12, e0187906 (2017).
Bystriakova, N., Peregrym, M., Erkens, R. H. J., Bezsmertna, O. & Schneider, H. Sampling bias in geographic and environmental space and its effect on the predictive power of species distribution models. Syst. Biodivers. 10, 305–315 (2012).
Hortal, J., Jiménez-Valverde, A., Gómez, J. F., Lobo, J. M. & Baselga, A. Historical bias in biodiversity inventories affects the observed environmental niche of the species. Oikos 117, 847–858 (2008).
Elith, J., Kearney, M. & Phillips, S. The art of modelling range-shifting species. Methods Ecol. Evol. 1, 330–342 (2010).
Stohlgren, T. J., Jarnevich, C. S., Esaias, W. E. & Morisette, J. T. Bounding species distribution models. Curr. Zool. 57, 642–647 (2011).
Mokany, K. & Ferrier, S. Predicting impacts of climate change on biodiversity: a role for semi-mechanistic community-level modelling. Divers. Distrib. 17, 374–380 (2011).
Ficetola, G. F., Pennati, R. & Manenti, R. Spatial segregation among age classes in cave salamanders: habitat selection or social interactions? Popul. Ecol. 55, 217–226 (2013).
Carvalho, J. C. et al. Biogeographic patterns of spiders in coastal dunes along a gradient of mediterraneity. Biodivers. Conserv. 20, 873–894 (2011).
Ward, D. & Saltz, D. Forging at Different Spatial Scales: Dorcas Gazelles Foraging for Lilies in the Negev Desert. Ecology 75, 48–58 (1994).
Alqamy, H. El & Din, S. B. El Contemporary status and distribution of gazelle species (Gazella dorcas and Gazella leptoceros) in Egypt. Zool. Middle East 39, 5–16 (2006).
Alqamy, H. Developing and Assessing a Population Monitoring Program for Dorcas Gazelle (Gazella dorcas) Using Distance Sampling in Southern Sinai, Egypt. (University of ST. Andrews, 2002).
Osborn, D. J. & Helmy, I. The contemporary land mammals of Egypt (including Sinai), https://doi.org/10.5962/bhl.title.2801 (Fieldeiana Zoology, New Series, 1980).
Durant, S. M. et al. The global decline of cheetah Acinonyx jubatus and what it means for conservation. Proc. Natl. Acad. Sci. USA 114, 528–533 (2017).
Stralberg, D. et al. Re-Shuffling of Species with Climate Disruption: A No-Analog Future for California Birds? PLoS One 4, e6825 (2009).
Rouget, M., Cowling, R. M., Pressey, R. L. & Richardson, D. M. Identifying spatial components of ecological and evolutionary processes for regional conservation planning in the Cape Floristic Region, South Africa. Divers. Distrib. 9, 191–210 (2003).
Millar, C. I., Stephenson, N. L. & Stephens, S. L. Climate change and forests of the future: managing in the face of uncertainty. Ecol. Appl. 17, 2145–2151 (2007).
Lawler, J. J. Climate Change Adaptation Strategies for Resource Management and Conservation Planning. Ann. N. Y. Acad. Sci. 1162, 79–98 (2009).
Peel, M. C., Finlayson, B. L. & McMahon, T. A. Updated world map of the Köppen-Geiger climate classification. Hydrol. Earth Syst. Sci. 11, 1633–1644 (2007).
Wilson, D. E. & Reeder, D. M. (editors). Mammal Species of the World. A Taxonomic and Geographic Reference. (3rd ed. Johns Hopkins University Press, 2, 142pp. Available at http://www.press.jhu.edu, 2005).
Shcheglovitova, M. & Anderson, R. P. Estimating optimal complexity for ecological niche models: A jackknife approach for species with small sample sizes. Ecol. Modell. 269, 9–17 (2013).
Radosavljevic, A. & Anderson, R. P. Making better M axent models of species distributions: complexity, overfitting and evaluation. J. Biogeogr. 41, 629–643 (2014).
Anderson, R. P. & Raza, A. The effect of the extent of the study region on GIS models of species geographic distributions and estimates of niche evolution: preliminary tests with montane rodents (genus Nephelomys) in Venezuela. J. Biogeogr. 37, 1378–1393 (2010).
Phillips, S. J. et al. Sample selection bias and presence-only distribution models: implications for background and pseudo-absence data. Ecol. Appl. 19, 181–197 (2009).
El-Gabbas, A. & Dormann, C. F. Wrong, but useful: regional species distribution models may not be improved by range-wide data under biased sampling. Ecol. Evol. 8, 2196–2206 (2018).
Kramer-Schadt, S. et al. The importance of correcting for sampling bias in MaxEnt species distribution models. Divers. Distrib. 19, 1366–1379 (2013).
Fourcade, Y., Engler, J. O., Rödder, D. & Secondi, J. Mapping species distributions with MAXENT using a geographically biased sample of presence data: a performance assessment of methods for correcting sampling bias. PLoS One 9, e97122 (2014).
Boria, R. A., Olson, L. E., Goodman, S. M. & Anderson, R. P. Spatial filtering to reduce sampling bias can improve the performance of ecological niche models. Ecol. Modell. 275, 73–77 (2014).
Beck, J., Böller, M., Erhardt, A. & Schwanghart, W. Spatial bias in the GBIF database and its effect on modeling species’ geographic distributions. Ecol. Inform. 19, 10–15 (2014).
Fourcade, Y., Engler, J. O., Besnard, A. G., Rödder, D. & Secondi, J. Confronting expert-based and modelled distributions for species with uncertain conservation status: A case study from the corncrake (Crex crex). Biol. Conserv. 167, 161–171 (2013).
Anderson, M. J. et al. Navigating the multiple meanings of β diversity: a roadmap for the practicing ecologist. Ecol. Lett. 14, 19–28 (2011).
Bradley, B. A. Predicting abundance with presence-only models. Landsc. Ecol. 31, 19–30 (2016).
Veloz, S. D. Spatially autocorrelated sampling falsely inflates measures of accuracy for presence-only niche models. J. Biogeogr. 36, 2290–2299 (2009).
Randin, C. F. et al. Climate change and plant distribution: local models predict high-elevation persistence. Glob. Chang. Biol. 15, 1557–1569 (2009).
Graham, M. H. Confronting multicollinearity in ecological multiple regression. Ecology 84, 2809–2815 (2003).
Townsend Peterson, A., Papeş, M. & Eaton, M. Transferability and model evaluation in ecological niche modeling: a comparison of GARP and Maxent. Ecography. 30, 550–560 (2007).
Quinn, G. P. & Keough, M. J. Experimental design and data analysis for biologists. (Cambridge University Press, 2002).
Naimi, B. usdm: Uncertainty Analysis for Species Distribution Models. R Packag. version 1.1-15, https://cran.r-project.org/package=usdm (2015).
Hijmans, R. J., Cameron, S. E., Parra, J. L., Jones, P. G. & Jarvis, A. Very high resolution interpolated climate surfaces for global land areas. Int. J. Climatol. 25, 1965–1978 (2005).
Dormann, C. F. et al. Collinearity: a review of methods to deal with it and a simulation study evaluating their performance. Ecography. 36, 27–46 (2013).
IPCC. Climate change 2014: synthesis report. Contribution of working groups I, II and III to the fifth assessment report of the intergovernmental panel on climate change (2014).
Riahi, K. et al. RCP 8.5—A scenario of comparatively high greenhouse gas emissions. Clim. Change 109, 33–57 (2011).
DiMiceli, C. M. et al. Annual Global Automated MODIS Vegetation Continuous Fields (MOD44B) at 250 m Spatial Resolution for Data Years Beginning Day 65, 2000–2010, Collection 5 Percent Tree Cover. (2011).
Martin, Y., Van Dyck, H., Dendoncker, N. & Titeux, N. Testing instead of assuming the importance of land use change scenarios to model species distributions under climate change. Glob. Ecol. Biogeogr. 22, 1204–1216 (2013).
Stanton, J. C., Pearson, R. G., Horning, N., Ersts, P. & Reşit Akçakaya, H. Combining static and dynamic variables in species distribution models under climate change. Methods Ecol. Evol. 3, 349–357 (2012).
Mesgaran, M. B., Cousens, R. D. & Webber, B. L. Here be dragons: a tool for quantifying novelty due to covariate range and correlation change when projecting species distribution models. Divers. Distrib. 20, 1147–1159 (2014).
Araújo, M. B. & New, M. Ensemble forecasting of species distributions. Trends Ecol. Evol. 22, 42–47 (2007).
Marmion, M., Parviainen, M., Luoto, M., Heikkinen, R. K. & Thuiller, W. Evaluation of consensus methods in predictive species distribution modelling. Divers. Distrib. 15, 59–69 (2009).
Phillips, S. J., Anderson, R. P. & Schapire, R. E. Maximum entropy modeling of species geographic distributions. Ecol. Modell. 190, 231–259 (2006).
Breiman, L. Random Forests. Mach. Learn. 45, 5–32 (2001).
Elith, J., Leathwick, J. R. & Hastie, T. A working guide to boosted regression trees. J. Anim. Ecol. 77, 802–813 (2008).
McCullagh, Peter & Nelder, J. Generalized Linear Models. (Chapman and Hall, 1989).
Thuiller, W., Georges, D. & Engler, R. biomod2: Ensemble platform for species distribution modeling. R Packag. version 3.3-13/r726, https://r-forge.r-project.org/projects/biomod/ (2016).
Belkhiria, J. et al. Application of Species Distribution Modeling for Avian Influenza surveillance in the United States considering the North America Migratory Flyways. Sci. Rep. 6, 33161 (2016).
Hung, C.-M. et al. Drastic population fluctuations explain the rapid extinction of the passenger pigeon. Proc. Natl. Acad. Sci. USA 111, 10636–41 (2014).
Warren, D. L., Wright, A. N., Seifert, S. N. & Shaffer, H. B. Incorporating model complexity and spatial sampling bias into ecological niche models of climate change risks faced by 90 California vertebrate species of concern. Divers. Distrib. 20, 334–343 (2014).
Anderson, R. P. & Gonzalez, I. Species-specific tuning increases robustness to sampling bias in models of species distributions: An implementation with Maxent. Ecol. Modell. 222, 2796–2811 (2011).
Brown, J. L., Cameron, A., Yoder, A. D., Vences, M. & Jarvis, A. A necessarily complex model to explain the biogeography of the amphibians and reptiles of Madagascar. Nat. Commun. 5, 5046 (2014).
Phillips, S. J. & Dudík, M. Modeling of species distributions with Maxent: new extensions and a comprehensive evaluation. Ecography. 31, 161–175 (2008).
Phillips, S. J. Transferability, sample selection bias and background data in presence-only modelling: a response to Peterson et al. (2007). Ecography. 31, 272–278 (2008).
Bagchi, R. et al. Forecasting potential routes for movement of endemic birds among important sites for biodiversity in the Albertine Rift under projected climate change. Ecography. 41, 401–413 (2018).
Baker, D. J. et al. Assessing climate change impacts for vertebrate fauna across the West African protected area network using regionally appropriate climate projections. Divers. Distrib. 21, 991–1003 (2015).
Bagchi, R. et al. Evaluating the effectiveness of conservation site networks under climate change: accounting for uncertainty. Glob. Chang. Biol. 19, 1236–1248 (2013).
Fielding, A. H. & Bell, J. F. A review of methods for the assessment of prediction errors in conservation presence/absence models. Environ. Conserv. 24, 38–49 (1997).
Allouche, O., Tsoar, A. & Kadmon, R. Assessing the accuracy of species distribution models: prevalence, kappa and the true skill statistic (TSS). J. Appl. Ecol. 43, 1223–1232 (2006).
Baker, D. J., Hartley, A. J., Butchart, S. H. M. & Willis, S. G. Choice of baseline climate data impacts projected species’ responses to climate change. Glob. Chang. Biol. 22, 2392–2404 (2016).
Buisson, L., Thuiller, W., Casajus, N., Lek, S. & Grenouillet, G. Uncertainty in ensemble forecasting of species distribution. Glob. Chang. Biol. 16, 1145–1157 (2010).
Goberville, E., Beaugrand, G., Hautekèete, N.-C., Piquot, Y. & Luczak, C. Uncertainties in the projection of species distributions related to general circulation models. Ecol. Evol. 5, 1100–1116 (2015).
Cianfrani, C., Broennimann, O., Loy, A. & Guisan, A. More than range exposure: Global otter vulnerability to climate change. Biol. Conserv. 221, 103–113 (2018).
Beale, C. M. et al. Incorporating uncertainty in predictive species distribution modelling. Philos. Trans. R. Soc. Lond. B. Biol. Sci. 367, 247–58 (2012).
Barbet-Massin, M., Thuiller, W. & Jiguet, F. How much do we overestimate future local extinction rates when restricting the range of occurrence data in climate suitability models? Ecography. 33, 878–886 (2010).
Jiang, H. et al. Predicting the Potential Distribution of Polygala tenuifolia Willd. under Climate Change in China. PLoS One 11, e0163718 (2016).
Pineda, E. & Lobo, J. M. Assessing the accuracy of species distribution models to predict amphibian species richness patterns. J. Anim. Ecol. 78, 182–190 (2009).
González-Orozco, C. E. et al. Quantifying Phytogeographical Regions of Australia Using Geospatial Turnover in Species Composition. PLoS One 9, e92558 (2014).
Oksanen, J. et al. vegan: Community Ecology Package. R Packag. version 2.4-4, https://CRAN.R-project.org/package=vegan (2017).
IUCN. IUCN Red List Categories and Criteria: Version 3.1 (IUCN, 2012).
IUCN Standards and Petitions Subcommittee. Guidelines for Using the IUCN Red List Categories and Criteria. Version 11 (2014).
Acknowledgements
This manuscript is part of a PhD thesis, which has been published according to the requirements of the University of Konstanz awarding the qualification to Alaaeldin Soultan in 2018. AS was financially supported by the German Academic Exchange Service (DAAD) and International Max Planck Research School (IMPRS). We appreciate the valuable comments and suggestions from Anne Scharf, Omar Attum, Ahmed El-Gabbas and Jessica Schnell on this manuscript.
Author information
Authors and Affiliations
Contributions
A.S., M.W. and K.S. designed the research. A.S. collected the data and conducted the analyses. All authors contributed to writing the main manuscript text. All authors reviewed the manuscript.
Corresponding author
Ethics declarations
Competing Interests
The authors declare no competing interests.
Additional information
Publisher’s note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Soultan, A., Wikelski, M. & Safi, K. Risk of biodiversity collapse under climate change in the Afro-Arabian region. Sci Rep 9, 955 (2019). https://doi.org/10.1038/s41598-018-37851-6
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-018-37851-6
This article is cited by
-
Uncovering the dynamics in global carbon dioxide utilization research: a bibliometric analysis (1995–2019)
Environmental Science and Pollution Research (2021)
-
Using species distribution models to gauge the completeness of the bat checklist of Eswatini
European Journal of Wildlife Research (2021)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
