
Abstract
Neuropeptide Y (NPY), a powerful neurotransmitter of the central nervous system, is a key regulator of angiogenesis and biology of adipose depots. Intriguingly, its peripheral vascular and angiogenic powerful activity is strictly associated to platelets, which are source of clinical hemoderivates, such as platelet lysate (PL), routinely employed in several clinical applications as wound healing, and to preserve ex vivo the progenitor properties of the adipose stromal cells pool. So far, the presence of NPY in PL and its biological effects on the adipose stromal cell fraction (ASCs) have never been investigated. Here, we aimed to identify endogenous sources of NPY such as PL-based preparations and to investigate which biological properties PL-derived NPY is able to exert on ASCs. The results show that PL contains a high amount of NPY, which is in part also excreted by ASCs when stimulated with PL. The protein levels of the three main NPY subtype receptors (Y1, Y2, Y5) are unaltered by stimulation of ASCs with PL, but their inhibition through selective pharmacological antagonists, considerably enhances migration, and a parallel reduction of angiogenic features of ASCs including decrease in VEGF mRNA and intracellular calcium levels, both downstream targets of NPY. The expression of VEGF and NPY is enhanced within the sites of neovascularisation of difficult wounds in patients after treatment with leuco-platelet concentrates. Our data highlight the presence of NPY in PL preparations and its peripheral effects on adipose progenitors.
Similar content being viewed by others
Introduction
Neuropeptide Y (NPY) is a 36-amino acid peptide produced by the central nervous system and implicated in physiological neuron-mediated activities such as food intake or circadian rhythm1,2. In the autonomous nervous system NPY is stored in macrovesicles close to the varicosities present along the end terminals of sympathetic nerve fibres, where it is located in association with the adrenergic mediators. NPY is the most abundant neuropeptide in the heart and brain and it is released during nerve activity as well as during ischemia, leading to vasoconstriction and smooth muscle cell proliferation3. Importantly, NPY receptors are heterogeneously expressed depending on the tissue of origin and cell type4,5. Specifically, Y1, Y2 and Y5 receptors are able to exert differentiated roles in stromal/mesenchymal progenitors compared to endothelial or neural cells, where they are reported to prevent neural death upon cytotoxic insults5,6. The Y5 receptor is mainly associated with proliferative and anti-aging properties in bone marrow-derived mesenchymal stem cells (BMMSCs)7, whereas Y1 and Y2 are known to influence the differentiation of stromal cells into the osteoblast-adipocyte lineage in vertebrates8,9,10, as well as in capillary angiogenesis11. Despite this heterogeneity, Y1, Y2 and Y5 receptors knockout mice similarly exhibit higher body weight, increased white adipose tissue and alteration of the cardiometabolic traits, suggesting that a multi- and combined receptor action exists12. Additionally, NPY exerts an anti-fibrotic potential in the ischemic heart1,13, thus contributing to tissue remodelling in the injured cardiac tissue. More importantly, NPY mediates the angiogenic process by enhancing endothelial cell proliferation in a calcium-dependent manner14. Intriguingly, it has been recently reported that platelets produce NPY, which is critical to mediate neo-angiogenesis upon in vivo ischemic insult15, exhibiting vascular activity comparable to that exerted by acknowledged angiogenic soluble mediators such as basic FGF or VEGF16,17,18. To date, regenerative medicine is focused on the identification of factors involved in the recruitment of stem cells to the site of injury and on molecules promoting maturation processes of undifferentiated progenitors.
In view of that, platelet lysate (PL), defined as an hemoderivate of platelets origin, is enriched with plethora of growth factors and biologically active mitogens19,20 and it is currently and safely employed in several clinical applications, in in vivo studies and as in vitro substitute to foetal bovine serum for the ex vivo expansion of the adipose stromal fraction21. Platelet lysate is known to enhance angiogenesis, a key feature for tissue injuries such as dermal wounds and ulcers, where the employment of PL or autologous leuco-platelet concentrate has been successfully consolidated14,15,22. Importantly, the restorative activity of PL both in vivo and in vitro consists in the enhancement of defined biological properties such as proliferation, migration and preservation of the clonogenic ability combined to changes of the cellular and phenotypic plasticity14,20,21,22,23. PL is also described to influence the biological behaviour of multiple adult cell populations19,20,22,24. The adipose stromal fraction represents the most reactive tissue source to PL. A large number of reports have clearly demonstrated the biological advantages and the efficacy of PL on the adipose tissue-derived stromal cells (ASCs), including modifications of the ASC-derived secretome, angiogenesis and differentiation capacity14,20,21,22,25.
Similarly to PL, NPY is either involved in the promotion of adipogenesis and inflammation and in the peripheral vasoconstriction/angiogenesis8,11,23,26, including wound healing10,27. More importantly, the mesenchymal fraction is a target of NPY, as demonstrated in the bone marrow niche, whose homeostasis is strictly controlled by NPY28.
The presence of NPY in PL-based preparations can be only indirectly assumed, as it has never been proven. Consequently, the biological and molecular mechanisms mediating PL-derived NPY action are still to be fully unravelled. Notably, Tilan et al. have demonstrated that during the late stages of ischemia in a murine model, systemic levels of NPY are enhanced in platelets by circulating megakaryocytes, therefore boosting capillary angiogenesis and vessel maturation at the sites of neovascularization11. This study has significantly highlighted the close dependence between NPY originated from platelets and peripheral vascular/angiogenic effects.
Accordingly, we investigated if PL may be identified as endogenous source of NPY with bio-activating properties, and in particular if NPY may contribute as endogenous mediator to the intrinsic biological properties of PL on the human adipose stromal fraction, routinely expanded ex vivo using this hemoderivate.
Our findings highlight that PL contains a high amount of endogenous NPY, which is able to significantly modulate ASCs angiogenesis and chemotaxis. Moreover, the blocking of NPY action through the selective Y1, Y2 and Y5 antagonist receptors inhibition, significantly downregulates intracellular calcium levels in ASCs stimulated with PL, without impacting on ERK signalling pathway and nitric oxide (NO) levels.
Overall, our data demonstrate the contribution of PL-derived NPY in mediating both angiogenesis and migration of human ASCs, identifying a potential calcium-dependent mechanism of action and elucidating the somatotropic and biological effects of NPY.
Materials and Methods
Isolation, expansion and in vitro stimulation of adipose stromal cells (ASCs)
Human adipose stromal cells (ASCs) were isolated from subcutaneous depots as we previously reported20,21,22. Briefly, biopsies were washed with PBS without Ca2+/Mg2+ and erythrocytes were lysed (Qiagen, Hilden, Germany). Samples were then mechanically chopped and enzymatically digested in Trypsin/EDTA and 1 mg/ml Collagenase I (Gibco, Invitrogen) for 45 minutes at 37 °C. Samples were then filtered and serially spun at 1200 rpm for 5 minutes. ASC cultures were seeded at 4000 cells/cm2 in DMEM-Low glucose in 10% Fetal Bovine Serum (FBS)/200mM L-glutammine/100 mM sodium pyruvate/non-essential aminoacids/penicillin/Amphotericin. After 3 days, the whole media was removed and the cultures expanded until reaching subconfluence. Viability was performed by Trypan Blue exclusion assay. At passage 3, cells were starved with 0.2%PL (patented GMP-compliant Platelet Lysate, Mesengen, Italy)14,19,20,21,22 overnight, then stimulated with DMEM-Low glucose/10%PL ± NPY subtype receptors antagonists (0.01 M Y1, BIBP3226 Cat. N. 2707; 0.025 M Y2, BIIE0246 Cat. N. 1700; 0.1 M Y5, L-152,804 Cat. N. 1382, all Tocris) for 48 hours. Procedures performed in this study involving human participants were in accordance with the ethical standards of the institutional Ethical Committee Policlinico Umberto I (Prot. 688/11 Rif. 2229/21.07.2011) and with the 1964 Helsinki declaration and its later amendments or comparable ethical standards. Informed consent was obtained from all individual participants included in the study.
Clonogenic and adipogenic differentiation assays
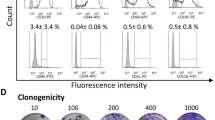
Clonogenic potential has been performed as already described20,22. Briefly, secondary colonies were obtained by seeding at low density (10 cells/cm2) in complete medium with 10%PL and ± NPY subtype receptor antagonists. Cultures were incubated for 14 days at 37 °C. Colonies produced were fixed with 4% paraformaldehyde and then stained with Giemsa (Sigma, Milan, Italy) for 1 hour and counted by optical microscope. A cluster of >50 cells was considered as a colony. The Colony forming efficiency (%) was calculated as ratio: N. secondary colonies/N. plated cells x 100. For adipogenic differentiation tests ASCs (1 × 104 cells/cm2 were cultured for 3 weeks using StemPro® Adipogenesis Differentiation kit (GIBCO, Grand Island, NY, USA, Cat. N. A10070-01)20,22. Accumulation of lipid droplets was evaluated by Oil Red Oil staining (Sigma-Aldrich, St. Louis, MO, USA, Cat. N. 01391-500 ml). Adipogenesis was quantified in 10 randomly selected fields observed under an optical microscope. The number of positive adipocytes was counted. The percentage of adipogenic differentiation was calculated as following: (N. adipogenic cells/N. Total cells) x 100.
Migration assay
After starvation human ASCs were resuspended in DMEM serum free ± Y1, Y2 and Y5 receptor antagonists and seeded at 5 × 104/well in the upper side of 96 well Boyden chamber (8 µm pores, BD Biosciences). The lower chamber contained DMEM or 10% PL ± Y1, Y2 and Y5 receptor antagonists. Cells were incubated at 37 °C for 16 hours. Afterwards, the inner side of the insert was wiped with a wet swab, in order to remove the cells and the outer side of the insert gently rinsed with PBS, fixed with methanol for 20 minutes and stained with 0.25% Giemsa (Sigma, St. Louis, USA) for 30 minutes, rinsed again, and dried. Migrated cells were counted (ten fields per chamber) under an optical microscope. Experiments were repeated three times in duplicate.
Proliferation assay
Cell proliferation was evaluated by MTS assay as already described19 by employing the CellTiter 96 AQueus One Solution Cell Proliferation Assay (Promega, Milan, Italy) and according to manufacturer’s instructions. After starvation (0.2% PL), ASCs were seeded in 96 well plates (cell density of 6 × 103 cells/well) in quintuplicate and conditioned for 48 hours with 10% PL ± NPY subtype receptor antagonists. The stimulation with recombinant NPY (10−9 M, Tocris Cat. N. 1153) was considered as biological control. The optical density signal measured at 492 nm was normalized to that of day 0 after cell seeding.
In vitro angiogenesis
Adipose stromal cells-derived angiogenesis was evaluated by Matrigel assay. Fifty microliters of Matrigel matrix growth factor reduced (BD, San Jose, CA, USA, Cat. N. 356231) were layered in 96-well plates and incubated for 1 h at 37 °C. After treatment with DMEM (unstimulated control) or 10% PL ± NPY subtype receptor antagonists, ASCs were then trypsinized and seeded on the top of the solidified Matrigel (1 × 104 cells/well). The cultures were incubated at 37 °C and after 2 hours, cells were checked under an optical microscope then photographed. Parameters including number of loops and network length were quantified by ImageJ Angiogenesis Analyser Software (National Institutes of Health, USA).
ELISA assay
Soluble levels of NPY in undiluted PL preparations, 10% PL fractions, DMEM (used as negative control) and ASC-derived conditioned media have been detected by ELISA assay (Elabscience, Cat. No. E-EL-H1893) according to manufacturer’s instruction.
Western Blotting
Cells were washed and lysed for 30 minutes at +4 °C in equal volumes of ice-cold RIPA lysis buffer (10 mM Tris pH 8, 1%Triton, 0,1% SDS, 0,1% Deoxycholate, 140 mM NaCl, 1 mM EDTA) and a cocktail of protease inhibitors (1 mM DTT, 1 mM PMSF, 1 Protease inhibitor tablet/10 mL Sigma, St. Louis, USA). Homogenates were incubated in ice for 20 minutes, then clarified by centrifugation at 14460 g for 15 minutes. Supernatants were boiled for 5 minutes in Laemmli sample buffer. Protein concentration was determined by Bradford and then separated by 15% SDS-PAGE electrophoresis and electrotransferred to polyvinylidene difluoride membranes. Incubation was performed overnight with the primary antibodies: Rabbit anti-Y1 and Rabbit anti-Y2 (1:500, Bioss Cat. N. bs-1070R and Cat. N. bs-0937R, respectively), Rabbit anti-Y5 (1:500, Abcam Cat. N. ab43824), Rabbit Anti-NPY (1:1000), phospho-ERK1/2 (1:200) and ERK (1:200; all Cell Signaling Technology, Cat. N. 11976 S, 1:1000, Cat. N. #4695, Cat. N. #9106, respectively) and a horseradish peroxidase-conjugated rabbit secondary antibody (Amersham mouse Cat. N. NA93IVS, Amersham rabbit NA934VS). The membranes were developed by enhanced chemiluminescence kit (ECL, GE Healthcare Bio-Science, Piscataway, NJ, USA). Acquisition and densitometry analysis was performed by ChemiDoc and Image Lab 5.2.1 (Biorad).
Immunofluorescence
In order to detect the expression of Y1, Y2 and Y5 on treated ASCs, cells were fixed with 4% Paraformaldehyde for 15–20 minutes at room temperature then washed 3X with PBS. Cells were then washed again with PBS, then resuspended in blocking buffer (0.25%gelatin/4%FBS) for 1 hour at room temperature. Primary antibodies (Anti-Y1 Bioss bs-1070R; Anti-Y2 Bioss bs-0937R; Anti-Y5 Abcam Cat. N. ab43824. All diluted 1:100) were incubated overnight in 0.25% gelatin at 4 °C. Goat Anti-Rabbit IgG (H + L) (Highly Cross-Adsorbed Secondary antibodies, Alexa Fluor-488, Thermo Fisher, Cat. No. #A-11034) for 45 minutes at room temperature in the dark were added after washing in PBS. Nuclei were stained with DAPI (4′−6′-Diamidino-2-phenylindole, powder ≥98%; Sigma, Milan, Italy Cat. N. D9542). Images were visualized and images acquired by Nikon Eclipse.
Real Time PCR
Total RNA was isolated (RNeasy kit, Qiagen) and c-DNA obtained and amplified by the SensiMix SYBR Hi-ROX kit (Bioline, London, UK). Templates were amplified by the 7900HT Fast Real Time PCR System (Applied Biosystems, Cheshire, UK) for 40 cycles according to the following protocol: 95 °C for 15 seconds, 60 °C for 10 seconds, 72 °C for 30 seconds. Primer sequences used are: NPY forward GCGACACTACATCAACCTCATC; NPY reverse TGTGCTTTCTCTCATCAAGAGG; VEGF forward AAAAACGAAAGCGCAAGAAA; VEGF reverse TTTCTCCGCTCTGAACAAGG; GAPDH forward ACAGTCAGCCGCATCTTC; GAPDH reverse GCCCAATACGACCAAATCC. GAPDH was considered as housekeeping. The reaction products were analysed by SDS 2.1.1 Software (Applied Biosystems, Cheshire, UK).
Nitric oxide (NO) levels
NO production in ASC-derived supernatants was evaluated by measurement of nitrite, a stable end product of NO. Nitrite was determined by a colorimetric assay with Griess reagent. One-hundred microliter of supernatant reacted with an equal volume of Griess reagent (one part of 1% sulfanilamide dissolved in 0.5 M HCl and one part of 0.1% N-(1-napthyl)-ethylenediamine dihydrocloride dissolved in distilled water) in 96-well plate under reduced light at room temperature during 10 minutes. The absorbance was measured with a microplate reader at 545 nm using a calibration curve of sodium nitrite standards (0–100 μM).
Calcium detection
After starvation, ASCs were trypsinized and washed in PBS and incubated with the treatments and the staining solution (3 μM Fluo-4-AM/2.5 mM Probenicide/3% DMSO, all Thermo Fisher) for 20 minutes at 37 °C. After incubation, cells were washed 2X with PBS and then resuspended with cold FACS buffer (PBS/2% FBS). Cells stimulated with Ionomycin (1 μM), EDTA (20 mM) or recombinant NPY (10−9 M) were used as positive, negative and biological internal control, respectively. Cytometry was performed using FACSAria II (B&D, San Jose, USA) and data were acquired and analysed by Software DiVa (v6.1.1, B&D, San Jose, USA).
Immunoistochemistry
Immunohistochemistry analysis was performed on paraffin-embedded consecutive sections (3 μm thickness) from difficult wounds. After deparaffinization with graded concentrations of xylene and alcohol, rehydration and blocking of endogenous peroxidases, samples were firstly incubated in 0.1% Trypsin/PBS and then with serum for 30 and 10 minutes, respectively. Primary antibodies Rabbit Anti-NPY (Cell Signaling Technology, Cat. N. 11976 S 1:100), human VEGF (Abcam, Cat. N. Ab46154, 1:200) or CD31 (Novocastra Leica, Cat N. NCL-CD31-1A10, 1:100) were incubated at 4 °C overnight. The staining was revealed by the avidin-biotin immunoperoxidase complex (Vectastain ABC kit, Vector Laboratories, Peterborough, UK) for single staining. For colocalization studies (NPY/CD31) secondary antibodies were the following: Ultratek HRP (Scytek Laboratories, Cat. N. UHM125 and AFK600) and HighDef Blue IHC chromogen (Enzo, Cat. N. ADI-950-151-0030) and AEC substrate kit (Scytek Laboratories, Cat. N. ACG500). Negative controls consisted of the secondary antibody alone and the use of the Immunoglobulin from the same animal species of the primary antibodies. Mayer’s haematoxylin was used as nuclear counterstain. The Images of the samples were analyzed under a light microscope (Nikon Eclipse). At least 4 random fields were acquired and quantified by identifying the foci of angiogenesis. Results have been expressed as the following ratio = Number of positive foci of angiogenesis/Number total foci of angiogenesis and normalized vs time 0.
Statistical Analysis
Statistical Analysis was performed by using software GraphPad Prism 5 software (San Diego, USA). Comparison between two or more groups was performed by t test or ANOVA (Bonferroni correction or Fisher LSD post-test), respectively. A p value < 0.05 has been considered statistically significant. Data are presented as mean ± standard error unless specified.
Results
We have investigated whether PL preparations contain a source of endogenous NPY, and to this aim we have quantified soluble NPY levels in five different batches of PL by ELISA. Results have shown that undiluted PL-based preparations contain the highest amount of NPY (Fig. 1a, p < 0.0001vs both DMEM and 10% PL), which is still quantifiable in the corresponding 10% PL preparations (the percentage of PL provided as supplement to ASCs cultures in the following experiments). The levels of NPY in the sole DMEM were undetectable (Fig. 1a).

(a–c) ELISA assay for the detection of soluble human NPY levels in PL-based preparations and expression of Y1, Y2 and Y5 receptors on ASC cultures. (a) The graph shows the amount of NPY contained in undiluted platelet lysate preparations as well as in 10% PL. DMEM has been used as negative control. N = 5). PL, platelet lysate. #p < 0.0001. (b) Western blot analysis shows that the stimulation with 10% PL for 48 hours does not alter the protein levels of each of the three main receptors for NPY. GAPDH was used as loading control. N = 3. Cropped images are from the same samples split on three different gel runs (Y1, Y2, Y5). Full-length blots are shown in Supplementary Fig. 1d. (c) Immunofluorescence for Y1, Y2 and Y5 on ASCs cultures confirms the expression of the three main receptors for NPY. Magnification 20×. PL, platelet lysate. DAPI (blue, nuclei); Y1, Y2, Y5 receptors (green).
In light of this observation, we have explored the contribution of NPY in PL-mediated effects in ASCs expansion, routinely performed ex vivo using PL in the culture media20,21,22. Thus, after isolation of human ASCs, cultures have been stimulated with 10% PL for 48 hours. To explore whether treatment with PL was able to influence the levels of the three main functional active NPY receptors (Y1, Y2 and Y5)29,30, we have performed a Western blot analysis. Results have displayed that the stimulation with PL was unable to modulate the protein levels of any of three main NPY receptors (Fig. 1b, Y1 p = 0.45, Y2 p = 0.62, Y5 p = 0.25). The presence of the three main receptors for NPY was confirmed by immunofluorescence. The cellular distribution of NPY receptors on the ASCs cultures showed a distinct and punctuate localization of the Y5 receptor respect to Y1 and Y2, both displaying a more diffuse pattern (Fig. 1c).
In order to understand the potential contribution of ASCs in sourcing NPY beyond PL and considering that the 10% PL media provided a considerable amount of soluble NPY (Fig. 1a), we stimulated ASC cultures with 10% PL for 48 hours and evaluated possible alterations of soluble levels of NPY in ASC-derived conditioned media. Results have shown a significant increased extracellular release of the neuropeptide compared to starved cultures (Fig. 2a, p = 0.036). This result was coherent with the enhancement of NPY protein levels in ASC cultures (Fig. 2b, p = 0.012), therefore strengthening the hypothesis regarding the contribution of ASCs in sourcing NPY in the microenvironment beyond the endogenous amount in PL-based preparations.

(a,b) NPY amount from ASCs and ASC-conditioned media (a) ELISA assay for soluble human NPY. The levels of excreted NPY in ASC-conditioned media after 48 hours of stimulation with 10% PL are significantly higher compared to starvation. PL, platelet lysate; starv, starvation. *p < 0.05. N = 5. (b) Western blot analysis for NPY in ASCs. After 48 hours of stimulation with 10% PL, ASC cultures exhibit enhanced protein levels of NPY. Tubulin is the loading control. Cropped images are from samples ran on the same gel. Full-length blots are shown in Supplementary Fig. 2c. N = 4. PL, platelet lysate; starv, starvation. *p < 0.05.
Next, in order to prevent the response of ASCs to the PL-derived NPY, we have specifically inhibited the exogenous NPY action by blocking the three main receptors on cells surface through a combined pre-treatment with selective Y1, Y2 and Y5 competitive antagonists4,31,32. The targeted inhibition of the three receptors did not significantly influence the clonogenic efficiency and the differentiation capacity of ASCs towards the adipogenic lineage after treatment with 10% PL (Fig. 3a,b), suggesting that NPY is not the determining factor for these processes.

(a–c) ASCs clonogenic activity, adipogenic differentiation and chemotactic activity. (a) Bar graph shows that the clonogenic capacity of ASC cultures is not influenced by the stimulation with 10% PL and the selective blocking for 48 hours of NPY receptors antagonists. %CFE (Colony forming efficiency) has been expressed as N. secondary colonies/N. plated cells x 100. N = 5. (b) Similarly, the rate of adipogenic differentiation of ASCs remains unaltered. The rate (%) of adipogenic differentiation has been calculated as: (N. adipogenic cells/N. Total cells) x 100. N = 5. (c) The chemotactic ability of ASCs is significantly enhanced by selective blocking of Y1, Y2 and Y5 receptors after stimulation with 10% PL. *p < 0.05, **p < 0.01, #p < 0.0001. N = 4. Representative images of migrated ASCs stained with Giemsa after stimulation with DMEM, 10% PL alone or in presence of NPY receptor antagonists. Magnification 10×. PL, platelet lysate. NPYRAnts, NPY receptor antagonists.
Interestingly, selectively blocking NPY receptors significantly enhanced the chemotaxis capacity of ASCs both compared to the stimulation with 10% PL and to non-stimulated controls (DMEM. Figure 3c, p < 0.05 and p < 0.001, respectively). The treatment with 10% PL was able to significantly increase chemotactic efficiency of ASCs compared to DMEM as expected (Fig. 3c, p < 0.01). The acquisition of the enhanced migratory activity of ASCs upon the combination of 10% PL and receptors antagonists for NPY was not ascribable to an altered cell proliferation (Supplementary Fig. 3d).
More importantly, PL-induced angiogenesis in vitro (Fig. 4a, p < 0.01 vs DMEM, the unstimulated control) was reduced by blocking NPY receptors, as demonstrated by decreased number of both loops and network length generated by ASCs seeded onto Matrigel (Fig. 4a, p < 0.01). Notably, the degree of the reduced angiogenesis in presence of NPY receptors antagonists was comparable to unstimulated controls (Fig. 4a, p > 0.05). When cultures were supplemented with increasing concentrations of PL (15 and 20%) in presence of NPY receptor antagonists, the angiogenesis reduction effect was preserved only up to 15% PL (Supplementary Fig. 4d), indicating a partial concentration dependent effect. The stimulation with 10% PL was able per se to significantly increase the expression of VEGF mRNA compared to non-stimulated controls (Fig. 4b, p = 0.029), and more importantly the selective blocking of NPY receptors reduced VEGF mRNA levels in ASCs respect to 10% PL (Fig. 4b, p = 0.015), showing comparable levels to unstimulated controls. VEGF is a major angiogenic growth factor and is activated downstream of NPY signalling pathways1,8,20,22. The reduction in angiogenesis did not appear to be due to changes of NO levels released in ASC-derived conditioned media (Fig. 4c), whose production is reported to be upregulated by NPY1. However, in our experimental conditions the supplementation with the sole recombinant NPY did not alter NO levels, which were similar to unstimulated controls, although both significantly lower than those released after stimulation with 10% PL (Fig. 4c, both p < 0.001).

(a–c) Angiogenesis assay, VEGF expression and detection of nitric oxide soluble NO levels (a) Bar graph shows the decrease in vitro number of loops and network length found in Matrigel assays of ASCs cultured in 10% PL combined with or without NPY receptor antagonists, **p < 0.01. N = 5–6. Representative optical images of in vitro angiogenesis of ASCs. Magnification 4×. (b) Bar graph shows mRNA levels of VEGF detected by qPCR after stimulation with 10% PL and parallel blocking of NPY receptor in ASC cultures, compared to unstimulated control. N = 3. *p < 0.05. (c) Bar graph shows no significant alterations in the release of NO after blocking of NPY receptors with selective antagonists. However, 10% PL is able to significantly enhance the levels of NO respect to both unstimulated controls (DMEM) and recombinant NPY (10−9 M). N = 6. **p < 0.01. (rec NPY, recombinant NPY; PL, platelet lysate; NPYRAnts, NPY receptor antagonists.
As both chemotaxis and angiogenesis are regulated by calcium signal and NPY has been reported to increase intracellular calcium levels33,34, we have investigated whether the PL-dependent NPY inhibitory and regulatory effect on angiogenesis and chemotaxis, might involve alterations of intracellular calcium levels. We have found a significant decrease in the calcium sensitive probe Fluo-4AM staining of ASCs when NPY is antagonized by selective blocking of its receptors compared to PL alone (Fig. 5a, p = 0.028), indicating the ability of PL-derived NPY to regulate intracellular calcium stores upon stimulation with PL.

(a,b) Flow cytometry Analysis of ASC cultures stained with Fluo4AM and Western Blot Analysis of ERK signalling. (a) Boxplot displays the mean green fluorescence intensity of ASCs stimulated for 48 hours with 10% PL in presence of the selective antagonists for NPY receptors is significantly decreased compared to the treatment with 10% PL. The stimulation with recombinant NPY and Ionomycin have been used as biological and positive control, respectively. N = 3 biological replicates. *p < 0.05 (two-way ANOVA and Fisher LSD post-test). MFI, mean fluorescent intensity; PL, Platelet lysate; NPYRAnts, NPY receptor antagonists; rec NPY, recombinant NPY. (b) Bar graph shows that no difference in ERK phosphorylation is found in ASCs cultures after 48 hour of treatment with 10% PL combined to NPY receptor antagonists compared to the 10% PL. Nevertheless, the stimulation with recombinant NPY (10−9 M) is able to induce a significant increase of phospho ERK with respect to both treatments. N = 6. #p < 0.001. Densitometry is presented as the ratio (pERK 1/2/Total ERK) normalized to control (10% PL). Cropped images are from samples ran on the same gel. Full-length blots are shown in Supplementary Fig. 5c. PL, platelet lysate; NPYRAnts, NPY receptor antagonists.
Moreover, given that ERK phosphorylation is reported to be induced by NPY35,36,37,38 especially during chemotaxis39,40 and that the intracellular signalling cascade of NPY12 converging on ERK is also able to amplify growth-promoting effects34, we asked if ERK phosphorylation was altered after blocking of NPY. Our data have shown that the blocking of NPY receptors did not alter ERK phosphorylation in ASCs respect to 10% PL (Fig. 5b, p > 0.05), suggesting a negligible contribution of NPY to the ERK signalling pathway. However, the stimulation with recombinant NPY did significantly increase ERK phosphorylation either respect to 10% PL and when in combination with NPY receptor antagonists (Fig. 5b, both p < 0.0001).
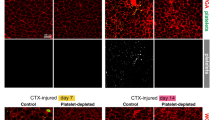
Finally, in order to confirm that the employment of PL-based preparations in vivo could foster an increase of NPY, we have focused on chronic wounds of patients that exhibit impaired healing and where PL is successfully employed as regenerative tool to enhance skin repair and local angiogenesis14,23,41,42. Specifically, we have selected human biopsy samples obtained from the edge of lesion of difficult wounds (defined as hard-to-heal or chronic wounds), which previously have been clinically treated with the autologous leuco-platelet concentrate, a PL-like preparation. Immunohistochemistry analysis on these biopsies has shown that after 48 hours of treatment with PL-like concentrates, the expression of NPY is increased in the ulcer and mainly localized at the sites of angiogenesis within newly formed capillaries, which are characterized by a cubic and reactive endothelium (Fig. 6a,b, p = 0.029). The colocalized expression of NPY and CD31, a marker of mature endothelial cells, was also observed (Fig. 6c). Similarly, the increase of VEGF after 48 hours of treatment occurs in parallel (Fig. 6a,b, p = 0.016), therefore indicating an enhanced local angiogenesis.

(a–c) Representative light microscopic appearance and staining for NPY, VEGF and CD31 of difficult wound biopsies from patients treated with leuco-platelet concentrates. (a) Bar graph showing the quantification of the expression of NPY and VEGF, which is significantly and concurrently enhanced after 48 hours of treatment. *p < 0.05. Results are expressed as the following ratio = No. Positive foci of angiogenesis/No. total foci of angiogenesis and normalized vs time 0. N = 3. (b) Staining for VEGF and NPY in patients difficult wound microscopic preparations. NPY is mainly distributed at the sites of angiogenesis including areas where it colocalizes with CD31 (c) a marker of mature endothelial cells. A similar localization of VEGF was also observed. Cell nuclei are counterstained with haematoxylin/eosin. In (c) CD31 stains blue and NPY red. Magnification 10X and 40X in b and c, respectively. The insert shows a higher magnification (20X and 60X in b and c, respectively).
Discussion
In this study for the first time we show that PL is a non-neural and endogenous source of NPY, which acts as active bioavailable vehicle on ASCs. Accordingly, in our experimental settings, NPY also originates from ASCs cultures, which respond to the stimulus (PL) either by increasing intracellular protein level and excreting NPY in the microenvironment. Thus, it is conceivable that a self-reinforcing loop of NPY upon stimulation with PL is produced by ASCs. Although, the ability to source NPY is not specific to stromal cells as already demonstrated26, however the stroma (the structural component of several stem cells niches and tissues) is a preferential target of NPY when released in the microenvironment28. Interestingly, the stimulation of ASCs with PL does not entail a specific upregulation of the three main receptors for NPY. This result indicates a potential synergism between receptors in response to the stimulus, leading us to consider that PL could influence more the activity or the function of Y1, Y2 and Y5 rather than the sole protein levels, as also reported in literature43. In fact, Y1, Y2 and Y5 expressed on ASCs are functional, as their simultaneous and selective blocking impacts both chemotaxis and angiogenesis but adipocyte differentiation and clonogenic ability. Platelet lysate is enriched with additional growth factors and mitogens, which equally preserve adipocytic differentiation of stromal cells, thus likely masking the effects of shutting down NPY by selective blocking of its receptors. Thus, the potential synergism between NPY and other soluble molecules contained in PL cannot be fully ruled out. Despite this, angiogenesis is significantly reduced by inhibiting NPY receptors in ASCs, even in presence of the additional growth factors in PL preparations, therefore strengthening the specificity of the paracrine and angiogenic action of PL-derived NPY on ASCs. In particular, our data indicate that the reduction of the extent of angiogenesis is restored to the unstimulated control level. This result suggests that NPY could represent the major player promoting PL-mediated angiogenesis. Accordingly, it has been demonstrated that the specific angiogenic contribution of NPY upon ischemia mainly originates from platelets, therefore clearly validating NPY as critical factor for revascularization upon damage conditions11. The angiogenic property of NPY has been well described in endothelial and vascular smooth muscle cells8 and in several tissues3,44. In this study, we implement the knowledge regarding the biological action of NPY on mesenchymal stem cells and specifically on the adipose progenitors instead of the mature adipose tissue or BMMSCs17,18. Notably, the origin of ASCs has been hypothesized from the stromal vascular fraction and specifically from the pericytes which are angiogenic populations closely associated to adipose progenitors20,22,45. In addition, the development of the adipose stromal population strictly depends on angiogenesis and ASCs are able to secrete growth factors22. The PL-derived NPY and that originated from ASCs in response might contribute to preserve this function in adipose progenitors. Interestingly, it has been reported that the pre-treatment of the mesenchymal cellular fraction with NPY prior to in vivo transplantation in murine models of myocardial infarction, increases cardiac differentiation of stromal cells combined with enhanced angiogenesis and amelioration of cardiac function16. However, to the best of our knowledge, no studies are currently investigating the angiogenic effects mediated by PL-derived NPY on ASCs. Notably, Tilan et al. have recently shown in a murine model of ischemia that the contribution of NPY during the late stages of recovery are dependent on the release of NPY from megakaryocyte/platelets activation, resulting in increased soluble levels of NPY in the systemic circulation and vascularization at the sites of angiogenesis11. Although it has been only demonstrated in endothelial and vascular smooth muscle cells, in line with our data this study strengthens the notion that NPY originating from platelets is able to increase cells angiogenic potential. In addition, adipose tissue-derived angiogenesis has been directly linked to the activation of NPY subtype receptors46, confirming that the angiogenic action of NPY is not limited to the neural, endothelial and vascular system47,48. Moreover, in our study we have highlighted the strict dependence of VEGF from NPY. Neuropeptide Y is upstream of VEGF signalling47, as well as either PL-based preparations and platelet activation have been implicated into the release of stored factors driving angiogenesis, including VEGF19,20,21,22,49,50. Upon ischemic damage, the activation of NPY subtype receptors triggers angiogenesis in endothelial cells through NO and release of VEGF8,48. Thus, the parallel decrease of angiogenesis and VEGF in ASCs after preventing the cross talk between cells and PL-derived NPY, recapitulates these effects. However, difference in NO soluble levels was not observed, likely due to the lack of a vascular insult, but also by the fact that in in vivo models of angiogenesis ascribable to NPY, the increase of NO does not always parallel VEGF8. Additionally, our results also highlight that inhibition of NPY receptors increases chemotaxis. The NPY is reported to possess bimodal features. Accordingly, its biological action has been described not only in a concentration-dependent fashion51 but also associated to the relative affinity with its receptors in the microenvironment17,34. Higher concentrations of NPY have been reported to neither increase cell proliferation, nor adipocytes differentiation and migration in wound scratch in vitro assays52,53. Thus, the observation that ASCs chemotaxis is increased by preventing the cross talk between NPY and ASCs is in line with a more regulatory role of NPY subjected to modulation in an activity-dependent manner. Interestingly, in our dataset, NPY shut down through receptors inhibition, decreases the intracellular calcium in ASCs, indicating that PL is able to primarily perturb calcium stores in ASCs through NPY. We cannot exclude that additional intracellular proteins are able to similarly mediate this function in the stromal fraction. Nevertheless, we found that the blocking of NPY function in ASCs alters both angiogenesis and chemotaxis, two main processes strictly restrained by calcium levels33. The NPY is reported to raise intracellular calcium levels in vascular smooth muscle and endothelial cells1,5,34,54 and to modulate cardiac contractility55,56. Notably, in some cell systems such as the retinal ganglion cells, NPY attenuates the calcium increase30. Although our results show a non-involvement of the ERK pathway, the stimulation with recombinant NPY instead of PL-derived NPY, strictly induces ERK phosphorylation. This suggests that the modality by which NPY is conveyed (in the form of single agent or as part of a plethora of soluble mediators contained in PL), may determine the activation of different biological and molecular signalling pathways. In this regard, the same effect has been observed for the release of NO in ASC-derived conditioned media.
So far, the majority of studies including those on BMMSCs17,18 have investigated the biological role of NPY by exogenous treatment with the recombinant protein. In this study, we have taken advantage of the NPY endogenously contained in PL, therefore reproducing a more physiological scenario. Accordingly, our data highlight that similarly to PL, leuco-platelet preparations employed in vivo may contain NPY. The tissue is able to respond by increasing the foci of angiogenesis where the positivity to NPY and VEGF is maximized. Coherent with our in vitro data, this result strengthens the significant the tropic contribution of platelets-derived NPY in periphery to induce angiogenesis and certainly migration (two main repairing processes amplified and enhanced in vivo by the employment of PL on difficult wounds). The role of NPY in wound healing has been demonstrated in mouse models and associated to its pro- and anti-inflammatory features, fostering chemotaxis and angiogenesis57 and involving macrophage-derived NPY within the adipose tissue58, a powerful modulator of inflammation22. Nevertheless, the precise molecular mechanisms and the modality by which PL-derived NPY could act in vivo on adipose tissue and cross talk with the immune system through the mesenchymal fraction, requires further investigation.
Our study has some limitations. First, we have hampered the in vitro response of ASCs to NPY rather than blocking NPY in PL by employing a neutralising antibody. This latter approach could have helped us to deeper understand if NPY is a discriminating factor in mediating the effects of PL. Similarly, we couldn’t inhibit the PL-derived NPY in patients for ethical reasons, therefore we are not able to fully confirm that the in vivo enhancement of PL-mediated angiogenesis is ascribable to the sole NPY. Finally, we have highlighted the in vitro effects of PL-derived NPY on the stromal lineage. Consequently it would be important to verify the involvement of the stromal fraction also in vivo.
In conclusion, this study has identified a non-neural source of NPY, present in the haemoderivate PL, and confirmed the trophic and peripheral angiogenic action of the peptide59. This feature could be exploited in wound healing as well as in other pathological cardiovascular disorders, where the enhancement of angiogenesis is desirable even indirectly by acting on the stromal progenitors. The combination of NPY and other growth repairing factors contained in PL, could also lead to new developed formulations of PL-based preparations, where the level of NPY could be modulated according to clinical needs, tissue-specificity and cell type.
References
Saraf, R., Mahmood, F., Amir, R. & Matyal, R. Neuropeptide Y is an angiogenic factor in cardiovascular regeneration. European journal of pharmacology 776, 64–70, https://doi.org/10.1016/j.ejphar.2016.02.033 (2016).
Reed, E. et al. Paclitaxel, cisplatin, and cyclophosphamide in human ovarian cancer: molecular rationale and early clinical results. Seminars in oncology 22, 90–96 (1995).
Zukowska-Grojec, Z. et al. Neuropeptide Y: a novel angiogenic factor from the sympathetic nerves and endothelium. Circulation research 83, 187–195 (1998).
Doods, H. N. et al. BIBP 3226, the first selective neuropeptide Y1 receptor antagonist: a review of its pharmacological properties. Regulatory peptides 65, 71–77 (1996).
Silva, A. P. et al. Neuropeptide Y expression, localization and cellular transducing effects in HUVEC. Biology of the cell 97, 457–467, https://doi.org/10.1042/BC20040102 (2005).
Xapelli, S., Silva, A. P., Ferreira, R. & Malva, J. O. Neuropeptide Y can rescue neurons from cell death following the application of an excitotoxic insult with kainate in rat organotypic hippocampal slice cultures. Peptides 28, 288–294, https://doi.org/10.1016/j.peptides.2006.09.031 (2007).
Igura, K., Haider, H., Ahmed, R. P., Sheriff, S. & Ashraf, M. Neuropeptide y and neuropeptide y y5 receptor interaction restores impaired growth potential of aging bone marrow stromal cells. Rejuvenation research 14, 393–403, https://doi.org/10.1089/rej.2010.1129 (2011).
Lee, N. J. et al. Critical role for Y1 receptors in mesenchymal progenitor cell differentiation and osteoblast activity. Journal of bone and mineral research: the official journal of the American Society for Bone and Mineral Research 25, 1736–1747, https://doi.org/10.1002/jbmr.61 (2010).
Lundberg, P. et al. Greater bone formation of Y2 knockout mice is associated with increased osteoprogenitor numbers and altered Y1 receptor expression. The Journal of biological chemistry 282, 19082–19091, https://doi.org/10.1074/jbc.M609629200 (2007).
Zhang, W., Bai, S., Liu, D., Cline, M. A. & Gilbert, E. R. Neuropeptide Y promotes adipogenesis in chicken adipose cells in vitro. Comparative biochemistry and physiology. Part A, Molecular & integrative physiology 181, 62–70, https://doi.org/10.1016/j.cbpa.2014.11.012 (2015).
Tilan, J. U. et al. Platelet neuropeptide Y is critical for ischemic revascularization in mice. FASEB journal: official publication of the Federation of American Societies for Experimental Biology 27, 2244–2255, https://doi.org/10.1096/fj.12-213546 (2013).
Brothers, S. P. & Wahlestedt, C. Therapeutic potential of neuropeptide Y (NPY) receptor ligands. EMBO molecular medicine 2, 429–439, https://doi.org/10.1002/emmm.201000100 (2010).
Matyal, R. et al. Local infiltration of neuropeptide Y as a potential therapeutic agent against apoptosis and fibrosis in a swine model of hypercholesterolemia and chronic myocardial ischemia. European journal of pharmacology 718, 261–270, https://doi.org/10.1016/j.ejphar.2013.08.023 (2013).
Businaro, R. et al. Multidisciplinary approaches to stimulate wound healing. Annals of the New York Academy of Sciences 1378, 137–142, https://doi.org/10.1111/nyas.13158 (2016).
Capoano, R. et al. Wounds Difficult to Heal: An Effective Treatment Strategy. Current vascular pharmacology 15, 582–588, https://doi.org/10.2174/1570161115666170301122216 (2017).
Wang, W. X. et al. Nerve growth factor induces cord formation of mesenchymal stem cell by promoting proliferation and activating the PI3K/Akt signaling pathway. Acta pharmacologica Sinica 32, 1483–1490, https://doi.org/10.1038/aps.2011.141 (2011).
Liu, S. et al. Neuropeptide Y stimulates osteoblastic differentiation and VEGF expression of bone marrow mesenchymal stem cells related to canonical Wnt signaling activating in vitro. Neuropeptides 56, 105–113, https://doi.org/10.1016/j.npep.2015.12.008 (2016).
Movafagh, S., Hobson, J. P., Spiegel, S., Kleinman, H. K. & Zukowska, Z. Neuropeptide Y induces migration, proliferation, and tube formation of endothelial cells bimodally via Y1, Y2, and Y5 receptors. FASEB journal: official publication of the Federation of American Societies for Experimental Biology 20, 1924–1926, https://doi.org/10.1096/fj.05-4770fje (2006).
Carducci, A. et al. GMP-grade platelet lysate enhances proliferation and migration of tenon fibroblasts. Frontiers in bioscience 8, 84–99 (2016).
Siciliano, C. et al. Optimization of the isolation and expansion method of human mediastinal-adipose tissue derived mesenchymal stem cells with virally inactivated GMP-grade platelet lysate. Cytotechnology 67, 165–174, https://doi.org/10.1007/s10616-013-9667-y (2015).
Siciliano, C. et al. The potential of GMP-compliant platelet lysate to induce a permissive state for cardiovascular transdifferentiation in human mediastinal adipose tissue-derived mesenchymal stem cells. BioMed research international 2015, 162439, https://doi.org/10.1155/2015/162439 (2015).
Siciliano, C. et al. The adipose tissue of origin influences the biological potential of human adipose stromal cells isolated from mediastinal and subcutaneous fat depots. Stem cell research 17, 342–351, https://doi.org/10.1016/j.scr.2016.07.010 (2016).
Sergeeva, N. S. et al. Analysis of Reparative Activity of Platelet Lysate: Effect on Cell Monolayer Recovery In Vitro and Skin Wound Healing In Vivo. Bulletin of experimental biology and medicine 162, 138–145, https://doi.org/10.1007/s10517-016-3563-3 (2016).
Robinson, S. T. et al. A novel platelet lysate hydrogel for endothelial cell and mesenchymal stem cell-directed neovascularization. Acta biomaterialia 36, 86–98, https://doi.org/10.1016/j.actbio.2016.03.002 (2016).
Dessels, C., Potgieter, M. & Pepper, M. S. Making the Switch: Alternatives to Fetal Bovine Serum for Adipose-Derived Stromal Cell Expansion. Frontiers in cell and developmental biology 4, 115, https://doi.org/10.3389/fcell.2016.00115 (2016).
Geloso, M. C., Corvino, V., Di Maria, V., Marchese, E. & Michetti, F. Cellular targets for neuropeptide Y-mediated control of adult neurogenesis. Frontiers in cellular neuroscience 9, 85, https://doi.org/10.3389/fncel.2015.00085 (2015).
Pradhan, L., Nabzdyk, C., Andersen, N. D., LoGerfo, F. W. & Veves, A. Inflammation and neuropeptides: the connection in diabetic wound healing. Expert reviews in molecular medicine 11, e2, https://doi.org/10.1017/S1462399409000945 (2009).
Park, M. H., Min, W. K., Jin, H. K. & Bae, J. S. Role of neuropeptide Y in the bone marrow hematopoietic stem cell microenvironment. BMB reports 48, 645–646 (2015).
Babilon, S., Morl, K. & Beck-Sickinger, A. G. Towards improved receptor targeting: anterograde transport, internalization and postendocytic trafficking of neuropeptide Y receptors. Biological chemistry 394, 921–936, https://doi.org/10.1515/hsz-2013-0123 (2013).
Martins, J. et al. Activation of Neuropeptide Y Receptors Modulates Retinal Ganglion Cell Physiology and Exerts Neuroprotective Actions In Vitro. ASN neuro 7, https://doi.org/10.1177/1759091415598292 (2015).
Mashiko, S. et al. A pair-feeding study reveals that a Y5 antagonist causes weight loss in diet-induced obese mice by modulating food intake and energy expenditure. Molecular pharmacology 71, 602–608, https://doi.org/10.1124/mol.106.029991 (2007).
Dumont, Y. et al. BIIE0246, a potent and highly selective non-peptide neuropeptide Y Y(2) receptor antagonist. British journal of pharmacology 129, 1075–1088, https://doi.org/10.1038/sj.bjp.0703162 (2000).
Kohn, E. C., Alessandro, R., Spoonster, J., Wersto, R. P. & Liotta, L. A. Angiogenesis: role of calcium-mediated signal transduction. Proceedings of the National Academy of Sciences of the United States of America 92, 1307–1311 (1995).
Pons, J. et al. Interactions of multiple signaling pathways in neuropeptide Y-mediated bimodal vascular smooth muscle cell growth. Canadian journal of physiology and pharmacology 86, 438–448, https://doi.org/10.1139/y08-054 (2008).
Shimada, K. et al. Neuropeptide Y activates phosphorylation of ERK and STAT3 in stromal vascular cells from brown adipose tissue, but fails to affect thermogenic function of brown adipocytes. Peptides 34, 336–342, https://doi.org/10.1016/j.peptides.2012.02.012 (2012).
Howell, O. W. et al. Neuropeptide Y stimulates neuronal precursor proliferation in the post-natal and adult dentate gyrus. Journal of neurochemistry 93, 560–570, https://doi.org/10.1111/j.1471-4159.2005.03057.x (2005).
Ruscica, M., Dozio, E., Motta, M. & Magni, P. Role of neuropeptide Y and its receptors in the progression of endocrine-related cancer. Peptides 28, 426–434, https://doi.org/10.1016/j.peptides.2006.08.045 (2007).
Hansel, D. E., Eipper, B. A. & Ronnett, G. V. Neuropeptide Y functions as a neuroproliferative factor. Nature 410, 940–944, https://doi.org/10.1038/35073601 (2001).
Zhang, P. et al. Neuropeptide Y Stimulates Proliferation and Migration of Vascular Smooth Muscle Cells from Pregnancy Hypertensive Rats via Y1 and Y5 Receptors. PloS one 10, e0131124, https://doi.org/10.1371/journal.pone.0131124 (2015).
Medeiros, P. J. et al. Neuropeptide Y stimulates proliferation and migration in the 4T1 breast cancer cell line. International journal of cancer 131, 276–286, https://doi.org/10.1002/ijc.26350 (2012).
Bonferoni, M. C. et al. Association of Alpha Tocopherol and Ag Sulfadiazine Chitosan Oleate Nanocarriers in Bioactive Dressings Supporting Platelet Lysate Application to Skin Wounds. Marine drugs 16, https://doi.org/10.3390/md16020056 (2018).
Caramella, C. M. et al. New therapeutic platforms for the treatment of epithelial and cutaneous lesions. Current drug delivery 10, 18–31 (2013).
Yi, M. et al. A Promising Therapeutic Target for Metabolic Diseases: Neuropeptide Y Receptors in Humans. Cellular physiology and biochemistry: international journal of experimental cellular physiology, biochemistry, and pharmacology 45, 88–107, https://doi.org/10.1159/000486225 (2018).
Ekstrand, A. J. et al. Deletion of neuropeptide Y (NPY) 2 receptor in mice results in blockage of NPY-induced angiogenesis and delayed wound healing. Proceedings of the National Academy of Sciences of the United States of America 100, 6033–6038, https://doi.org/10.1073/pnas.1135965100 (2003).
Brakenhielm, E. & Cao, Y. Angiogenesis in adipose tissue. Methods in molecular biology 456, 65–81, https://doi.org/10.1007/978-1-59745-245-8_5 (2008).
Kuo, L. E. et al. Neuropeptide Y acts directly in the periphery on fat tissue and mediates stress-induced obesity and metabolic syndrome. Nature medicine 13, 803–811, https://doi.org/10.1038/nm1611 (2007).
Lee, E. W. et al. Neuropeptide Y induces ischemic angiogenesis and restores function of ischemic skeletal muscles. The Journal of clinical investigation 111, 1853–1862, https://doi.org/10.1172/JCI16929 (2003).
Chen, S. H., Fung, P. C. & Cheung, R. T. Neuropeptide Y-Y1 receptor modulates nitric oxide level during stroke in the rat. Free radical biology & medicine 32, 776–784 (2002).
Hershey, J. C. et al. Revascularization in the rabbit hindlimb: dissociation between capillary sprouting and arteriogenesis. Cardiovascular research 49, 618–625 (2001).
Kondoh, K. et al. Conduction performance of collateral vessels induced by vascular endothelial growth factor or basic fibroblast growth factor. Cardiovascular research 61, 132–142 (2004).
Liu, M., Liu, H., Liang, F., Song, X. Q. & Hu, P. A. Neuropeptide Y promotes adipogenic differentiation in primary cultured human adipose-derived stem cells. Endocrine journal, https://doi.org/10.1507/endocrj.EJ17-0017 (2017).
Jain, M., LoGerfo, F. W., Guthrie, P. & Pradhan, L. Effect of hyperglycemia and neuropeptides on interleukin-8 expression and angiogenesis in dermal microvascular endothelial cells. Journal of vascular surgery 53, 1654–1660 e1652, https://doi.org/10.1016/j.jvs.2011.02.019 (2011).
Buttari, B. et al. Neuropeptide Y induces potent migration of human immature dendritic cells and promotes a Th2 polarization. FASEB journal: official publication of the Federation of American Societies for Experimental Biology 28, 3038–3049, https://doi.org/10.1096/fj.13-243485 (2014).
(ed Abba Kastin) Ch. 154, 1160-1165 (Academic Press, 2013).
Masliuko, P. M. et al. NPY1 Receptors Participate in the Regulation of Myocardial Contractility in Rats. Bulletin of experimental biology and medicine 162, 418–420, https://doi.org/10.1007/s10517-017-3629-x (2017).
Protas, L. & Robinson, R. B. Neuropeptide Y contributes to innervation-dependent increase in I(Ca, L) via ventricular Y2 receptors. The American journal of physiology 277, H940–946 (1999).
Zofia Zukowska, G. Z. F. The NPY Family of Peptides in Immune Disorders, Inflammation, Angiogenesis, and Cancer. (Springer Science & Business Media, 2006).
Singer, K. et al. Neuropeptide Y is produced by adipose tissue macrophages and regulates obesity-induced inflammation. PloS one 8, e57929, https://doi.org/10.1371/journal.pone.0057929 (2013).
Pedrazzini, T., Pralong, F. & Grouzmann, E. Neuropeptide Y: the universal soldier. Cellular and molecular life sciences: CMLS 60, 350–377 (2003).
Acknowledgements
We thank Futura Relife for providing Mesengen™ and Fondazione Roma for the continuous support. This study has been financed by Sapienza University, granted to Elena De Falco (Prot. No. C26A1578EJ) and Rita Businaro (Grant No. RM116154CCE6BAE7).
Author information
Authors and Affiliations
Contributions
E.S., A.B., F.P., M.C., C.S., V.P. performed all experiments, P.T. contributed to data collection and analysis, R.C., E.P., B.S. performed surgery, P.T. and MTV provided technical and experimental support, R.B., G.F. wrote the paper, R.B., E.D.F. designed the study and wrote the paper.
Corresponding author
Ethics declarations
Competing Interests
Prof. G. Frati holds a patent concerning platelet lysate in regenerative medicine (Platelet lysate, uses and method for the preparation thereof. Pub. number WO/2013/042095, International Application number PCT/IB2012/055062).
Additional information
Publisher's note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Businaro, R., Scaccia, E., Bordin, A. et al. Platelet Lysate-Derived Neuropeptide y Influences Migration and Angiogenesis of Human Adipose Tissue-Derived Stromal Cells. Sci Rep 8, 14365 (2018). https://doi.org/10.1038/s41598-018-32623-8
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-018-32623-8
Keywords
This article is cited by
-
Neuropeptide Y Promotes the Treatment of Adipose Stem Cells on Type 2 Diabetic Wounds
Tissue Engineering and Regenerative Medicine (2023)
-
Role of neurotransmitters in the regulation of cutaneous wound healing
Experimental Brain Research (2022)
-
Targeted genetic analysis unveils novel associations between ACE I/D and APO T158C polymorphisms with D-dimer levels in severe COVID-19 patients with pulmonary embolism
Journal of Thrombosis and Thrombolysis (2022)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
