
Abstract
Here we present a visual reverse transcription loop-mediated isothermal amplification (RT-LAMP) assay for detecting the gene encoding the σB major outer-capsid protein of novel duck reovirus (NDRV). A set of primers, composed of two outer primers, two inner primers and two loop primers, was designed based on the gene of interest. The LAMP reaction was conducted in a traditional laboratory water bath at 65 °C for 50 min. We compared the performance of calcein/Mn2+ and SYBR Green I dyes, as well as electrophoresis on agarose gel stained with GoldView nucleic acid dye to detect the RT-LAMP-amplified products and all assays could be employed to discriminate between positive and negative specimens in visible or UV light. Our data showed that there is no cross-reaction with other viruses and the RT-LAMP technique displayed high sensitivity for detecting NDRV with a minimal detection limit of 200 fg RNA input. This assay was more sensitive than conventional PCR in detecting NDRV both in natural and experimental infection. In conclusion, the RT-LAMP technique was remarkably sensitive, specific, rapid, simple and profitable for the identification of NDRV.
Similar content being viewed by others

Introduction
A novel duck reovirus (NDRV) disease, called “spleen necrosis disease,” “new liver disease in Muscovy ducks” or “duck hemorrhagic-necrotic hepatitis,” was recently found among several duckling species, including shelducks, Pekins, wild mallards and Muscovy in China1. Similarly, avian reovirus (ARV) infection was recorded in Muscovy ducks (Cairina moschata) in south western Poland during the summer 20122. NDRV is a member of the genus Orthoreovirus in the family Reoviridae3.
The disease can be distinguished from Muscovy duck reovirus (MDRV) infections by clinical presentation of hemorrhagic and necrotic lesions in the liver and spleen and 5–50% mortality rates. Furthermore, the existing commercial vaccines against ARVs or MDRVs fail to prevent NDRV infection and transmission. A reverse transcription loop mediated isothermal amplification (RT-LAMP) provides a rapid and precise detection assay for viral pathogens with high sensitivity and specificity4. This technique is profitable and convenient, only requires a constant temperature water bath and averts some deficiencies, including high necessity for equipment, tremendous cost, lengthened examination period and low sensitivity5. In comparison with the equipment required for traditional polymerase chain reaction (PCR) and quantitative polymerase chain reaction (qPCR) assays, RT-LAMP is easy to conduct in resource-limited laboratory settings in underdeveloped countries6. RT-LAMP has been widely exploited in clinical diagnosis of various viral pathogens including porcine epidemic diarrhea virus (PEDV), transmissible gastroenteritis virus (TGEV), classic swine fever virus (CSFV), H10N8 subtype of influenza A virus, porcine deltacoronavirus (PDCoV), Zika virus (ZIKV), chikungunya virus (CHIKV), Rift Valley fever virus (RVFV), St. Louis encephalitis virus (SLEV), yellow fever virus (YFV), dengue virus serotypes 1–4, Japanese encephalitis virus (DENV1–4) and West Nile virus (WNV)7,8,9,10. The ARV structural and molecular compositions are generally similar to those of mammalian reovirus (MRV). However, some of the ARV biological properties differ from mammalian reovirus, which have been shown elsewhere11,12. The S3 segment encoding the σB protein of duck and MRVs is structurally similar to ARV σB gene13. Reverse transcription PCR (RT-PCR) on both sigma C (σC) and sigma B (σB)-encoding genes followed by restriction fragment length polymorphism (RFLP) analyses were employed to characterize Tunisian ARV isolates14. Furthermore, the highly variable sequences of the S3 and M3 have been reported to differentiate between novel duck reovirus (NDR), MDRV and avian reovirus15. Thus, a simple, rapid and sensitive diagnostic technique for detection of NDRV is required. The potential application of RT-LAMP assay using the S3 gene of NDRV-NPO3 strain for specific diagnosis of NDRV infection with limited sensitivity and without its utilization in the detection of NDRV in naturally- and experimentally-infected ducks have been reported16. In this study, we developed and evaluated a RT-LAMP assay targeting the gene encoding the σB major outer-capsid protein to detect NDRV in naturally suspected NDRV-infected ducks and experimentally infected ducklings with NDRV.
Materials and Methods
Ethics statement
This study was approved by the Animal Ethics Committee of the College of life science and engineering, Foshan University, Guangdong, China. The College did not issue a number or ID to this animal study, because the studied ducks are not an endangered or protected species. Specimen collection was carried out based upon the protocol issued by the Animal Ethics Committee of the College of life science and engineering. Furthermore, all methods were performed in accordance with the relevant guidelines and regulations.
Virus isolates for specificity evaluation
Novel duck reovirus disease SH12 (NDRV- SH12) (preserved in the laboratory of Preventive Veterinary Medicine), was isolated from the infected ducks in Guandong province, China, Muscovy Duck Reovirus (MDRV-S12), Avian Reovirus (ARV-S1133), Duck Disease Virus (DPV), Duck Hepatitis Virus (DHAV), Duck Newcastle Disease Virus (NDV), H9 subtype avian influenza virus (AIV), H5 subtype AIV and duck Tanzuru virus (DTMUV) were stocked at −80 °C in our laboratory at College of life science and engineering, Foshan University, China. A total of 15 ducks were collected from the Guangdong region suspected to be infected with NDRV. The specimens were sent to the College of life science and engineering, Foshan University, China and examined upon arrival.
Experimental infection of ducklings
Ten 1-day-old Muscovy ducklings were randomly divided into 2 groups (5 birds per group). Group 1 ducklings were intraperitoneally injected with 0.2 mL (104.00ELD50) of the NDRV allantoic fluid and ducklings in group 2 were intraperitoneally injected with physiological saline and served as the control group. All ducklings were observed hourly for 72 h post-infection (hpi).
RNA extraction
Total RNA was extracted employing an EasyPure Viral DNA/RNA kit (Transgen, Beijing, China) according to the manufacturer’s protocol and was stored immediately at −80 °C until use.
Reverse transcriptase PCR (RT-PCR)
RT-PCR was performed employing a PrimeScript One Step RT-PCR Kit (TaKaRa, Japan). Primers S-F (5′-GCTTTTTGAGTCCTCAGCGTG-3′) and S-R (5′-GATGAATAGGCGAGTCCCGC-3′) were used to amplify the corresponding S3 segment encoding sigma B gene. For RT-PCR, 2.5 µL of extracted viral RNA was mixed with a reaction mixture containing 1 µL primer (10 pmol), 1 µL Prime Script One Step Enzyme mix, 8 µL RNase-free H2O and 12.5 µL 2 × One Step Buffer. Reverse transcription was performed at 50 °C for 30 min. The cycling conditions for the PCR were 94 °C for 2 min, followed by 32 cycles of denaturation (94 °C for 10 s), annealing (58 °C for 30 s) and extension (72 °C for 1 min), followed by a final extension at 72 °C for 7 min.
Cloning and sequencing
The amplified PCR products were subjected to agarose gel electrophoresis, excised from the gel and purified using an Agarose Gel DNA Purification Kit (TaKaRa, Japan). The PCR products were cloned into the pMD19-T vector according to the manufacturer’s instructions (TaKaRa, Japan). After the recombinant plasmid was transformed into DH5α competent cells, the plasmid DNA purified using the E.Z.N.A.® Plasmid Mini Kit I (Omega Bio-Tek, USA) and quantified by spectrophotometric analysis. Three clones were sent to Shanghai Sangon Bioengineering Ltd. to be sequenced using the aforementioned primer pair and the sequence was analyzed.
Design of primers for LAMP
Based on the gene encoding the σB major outer-capsid protein (GenBank accession number JQ866923.1; 1104 bp) of NDRV (DRB-QY strain), a set of six primers consisted of two outer primers (F3 and B3), two inner primers (FIP and BIP) and two loop primer (Loop primer Fc and Loop primer B) was generated employing the Primer Explorer V4 software (http://primerexplorer.jp/e/). The forward inner primer (FIP) composed of the complementary sequence of F1c and F2. The backward inner primer (BIP) consisted of B1c and the complementary sequence of B2c (B2). The outer primers consisted of the forward outer primer F3 and the backward outer primer B3 (the complementary sequence of B3c). In addition, there are two loop primers (Loop primer Fc and Loop primer B) to accelerate the amplification reaction as previously described17. The positions of the LAMP primers used in this study are shown in Fig. 1 and Table 1. Primers were synthesized by Shanghai Sangon Bioengineering Ltd, China.

Location of the NDRV RT-LAMP primer in the gene encoding the σB major outer-capsid protein.
RT-LAMP assay optimization
The RT-LAMP reaction was carried out in a final volume of 25 μL. To optimize the RT-LAMP assay, the parameters, including the amplification temperature (59–66 °C) and the amplification time (10–60 min) were determined. The parameters of the reaction mixtures, such as the ratio of inner and outer primers (2:1–12:1), the ratio of loop and outer primers (0:0.2–6:1) and the ratio of calcein and manganese ion (1:8–1:128) was described independently. In addition, the concentrations of betaine (0–1.5 mM; Sigma, USA), dNTPs (0.2–0.7 mM; Promega, USA), MgSO4 (0–6 mM; Sigma, USA), AMV reverse transcriptase (0.06–0.16 U/µL; Takara, Japan), Bst DNA polymerase (0.8–0.48 U/µL; New England Biolabs, USA) and 10 × ThermoPol buffer (0–4 µL) were sequentially optimized.
Detection of RT-LAMP products
The RT-LAMP reactions for NDRV were carried out at 65 °C for 50 min, followed by denaturation for 5 min at 85 °C to cease the reaction. The RT-LAMP-amplified products were visualized on 2% agarose gel stained with GoldViewTM under UV transillumination. The calcein visualization RT-LAMP assay can be inspected under UV light, which emitted strong green fluorescence. Moreover, products were visualized directly by addition of 1.0 µL of 1:10 diluted SYBR Green I (Invitrogen, USA). To determine the sequence specificity, the cleavage site of RT-LAMP amplified fragment was analyzed by SeqBuilder software and the single Hpa II restriction site was screened. Two microliters of RT-LAMP amplified products were digested with restriction enzyme HhaI (New England Biolabs, USA) at 37 °C for 2 h according to the manufacturer’s protocol and the restriction enzyme digestion products were analyzed on 2% agarose gel electrophoresis. Each reaction was repeated at least three times.
Specificity of RT- LAMP identification
To determine the specificity of RT-LAMP assay, RT-LAMP was conducted with the different nucleic acid templates extracted from Muscovy duck reovirus (MDRV-S12), avian reovirus (ARV-S1133), duck plague virus (DPV-YF15), duck hepatitis virus (DHAV-3), duck Newcastle disease virus (NDV-SS14), H9 subtype avian influenza virus (NH/2013), H5 subtype avian influenza virus (GM/2014) and duck virus (DTMUV-GD13) under the aforementioned conditions. Each virus was investigated at least three times.
Sensitivity of detection amongst RT-LAMP visualized by calcein, conventional RT-LAMP and RT-LAMP visualized by SYBR Green I
Detection sensitivities between RT-LAMP visualized by calcein, RT-LAMP analyzed by agarose gel and RT-LAMP visualized by SYBR Green I were compared using 10-fold serial dilutions of the virus-positive total RNA extracts (NDRV) and recombinant plasmid. The resulting concentrations were as follows: 2 ng, 200 pg, 20 pg, 2 pg, 200 fg, 20 fg and 2 fg. Detection limits were determined by the lowest input nucleic acid concentration at which a positive result was apparent.
Results
Optimization of RT-LAMP assay for NDRV detection
All of the possible variables were investigated to enhance the amplification efficiency. Three replications were carried out for each trial. RT-LAMP was performed under different reaction temperatures from 59 °C to 65 °C. The results showed that RT-LAMP at 65 °C produced very clear and bright bands. Therefore, 65 °C was selected as the optimal temperature of NDRV RT-LAMP reaction. The reaction time indicated that 50 min was adequate for reaction completion. The ratio of calcein and manganese ion (1:8–1:128) was analyzed and the color was obvious at the ratio of 1:12. Consequently, the ratio of 1:12 was selected as the optimal ratio to use in proceeding experiments. The effect of Mg2+ ion concentration was analyzed. Magnesium sulfate (Mg2+) was added at a concentrations ranging from 0 to 6 mM reaction mixtures containing positive sample. Upon gel analysis, bands were obvious at 3 mM Mg2+. When the concentration was greater than 3.0 mM (lane 4~7), the reaction efficiency decreased but tended to be stable. Therefore, 3.0 mM was chosen as the best reaction concentration for NDRV RT-LAMP. Moderately various yields were detected with the different amounts of betaine employed and a concentration of 0.5 mM was found to be proper for a more distinct pattern. The size of the ladder increased as the concentration of dNTPs increased and a concentration greater 0.3 mM was found to be sufficient. AMV reverse transcriptase concentration of 0.14 U/µL was found to be appropriate for the amplification efficiency of NDRV RT-LAMP reaction. The reaction efficiency decreased as the concentration of the AMV enzyme concentration increased. However, a more intense band was observed with 2.5 µL of 10 × Buffer, a 10:1 ratio of inner vs. outer primers (FIP + BIP 2 M, F3 + B3 0.2 µM) and a ratio of 4:1 ring and outer primers (LoopF + LoopB 0.8 M, F3 + B3 0.2 µM) and 0.32 U/L of Bst DNA polymerase (Figs S1; 2–3).

Procedures optimization of the NDRV RT-LAMP reaction. (A) The effect of temperature: M, 2000 bp DNA Marker; lanes 1–8 (66 °C, 65 °C, 64 °C, 63 °C, 62 °C, 61 °C, 60 °C and 59 °C, respectively) and lane 9: ddH2O (negative control; NC). (B) The effect of time: lanes 1–6 (10, 20, 30, 40, 50 and 60 min, respectively); lane 7 NC.

Primer optimization of the NDRV RT-LAMP reaction. (A) The effect of the ratio of inner and outer primers: M, 2000 bp DNA Marker, lanes 1–6 (2:1, 4:1, 6:1, 8:1, 10:1 and 12:1, respectively) and lane 7 NC. (B) The effect of the ratio of loop (Loop primer Fc and Loop primer B) and outer primers (F3 and B3): M, 2000 bp DNA Marker, lanes 1–7 (0:0.2, 1:1, 2:1, 3:1, 4:1, 5:1 and 6:1, respectively); lane 8 NC.
Taken together, the optimized parameters of the RT-LAMP reaction system were as follows: 3 mM MgSO4, 0.5 mM betaine, 0.3 mM dNTPs, 0.14 U/μL AMV reverse transcriptase, 0.32 U/μL Bst DNA polymerase large fragment, 2.5 μL10 × ThermoPol buffer, the ratio of inner (2.0 μM; FIP and BIP) and outer (0.2 μM; F3 and B3) primers and the ratio of loop (0.8 μM; Loop primer Fc and Loop primer B) and outer primers (0.2 μM; F3 and B3) with an incubation condition at 65 °C for 50 min in a water bath. The RT-LAMP-amplified products were resolved on 2.0% agarose gel and checked by visual investigation employing calcein under UV or the production of insoluble manganese/ magnesium phosphate and by electrophoresis on GoldViewTM nucleic acid stained agarose gels under UV light. Moreover, the RT-LAMP products were also visually detected by adding SYBR Green I dye.
Analysis of RT-LAMP digestion products
Electrophoresis of the RT-LAMP products showed that all the positive LAMP reactions produced characteristic ladder-like pattern bands on agarose gel. HhaI restriction enzyme digestion and electrophoresis gave a strong band at approximately 100 bp corresponding to the predicted sizes (a mixture of 127 bp and 114 bp) (Fig. 4).

Agarose gel electrophoresis results for NDRV RT-LAMP reaction products and their HhaI restriction enzyme digestion results. M, 2000 bp DNA Marker; lane 1(HhaI restriction enzyme digestion results); lane 2 ladder-like pattern bands; lane 3 NC.
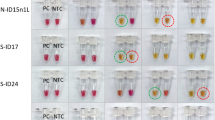
Specificity of RT-LAMP identification
As illustrated in Fig. 5, only when the recombinant plasmid and RNA from NDRV were present. The RT-LAMP reaction was positive and the RT-LAMP-amplified products were detected, showing a typical ladder-like pattern on gel electrophoresis, which indicated that stem-loop DNA with inverted repeats was generated, whereas RT-LAMP reaction was negative for double distilled water (negative control; NC) and the other viruses tested, indicating that the LAMP reaction was highly specific to NDRV.

Identification specificity of the NDRV RT-LAMP assay. Samples were resolved on 2.0% agarose gels. LAMP was carried out with the different sources of nucleic acids. Lane M, 2000 bp DNA ladder; Lane 1, recombinant plasmid; Lane 2 NDRV sample; Lanes 3–11, MDRV, ARV, DPV, DHAV, NDV, H9 AIV, H5 AIV, DTMUV and NC.
RT-LAMP sensitivity comparison
The sensitivity of RT-LAMP assay visualized by calcein was first judged by the dilution of RNA input and recombinant plasmid into 2 ng, 200 pg, 20 pg, 2 pg, 200 fg, 20 fg and 2 fg, respectively and the result of this assay was compared with those of RT-LAMP visualized by SYBR Green I and RT-PCR analyzed by agarose gel (Fig. S2A–F). The comparison study revealed that the detection limit of RT-LAMP visualized by calcein was 200 fg. This result is in accord with that of RT-LAMP visualized by SYBR Green I and RT-LAMP analyzed by agarose gel, but it is more sensitive than of conventional RT-PCR (20 pg detection limit).
RT-LAMP assay evaluation with field samples (suspected NDRV infection) from Guangdong province
A total of 15 clinical samples were used with their tissue mixed together for each individual duck and 12 clinical sample organs (including heart, liver, spleen, lung, thymus and bursa) were used separately from two NDRV-infected ducks in this study. The detailed results of detection (specimen by specimen) are listed in Tables 2 and 3. The comparative evaluation of RT-LAMP and conventional RT-PCR revealed that RT-LAMP and RT-PCR were consistent when the commonly affected organs were mixed together (Table 2). The viral RNAs of NDRV were detected in 20% (3/15) of the 15 duck studied. However, the positive rates of conventional RT-PCR and RT-LAMP for separately affected organs NDRV with were 83% (10/12) and 92% (11/12), respectively as indicated in Table 3.
RT-LAMP assay evaluation with experimentally infected ducklings
A total of ten 1-day-old ducklings were randomly allocated into two groups (five ducklings each). Group 1 animals were received intraperitoneal inoculation of NDRV allantoic fluid (Table 4). Ducklings from group 2 were intraperitoneally inoculated with physiological saline and served as the control group. Ducklings were followed hourly for 72 hours with their corresponding uninfected-control ones. All ducklings in the infected group died with 72 hours post-infection (hpi). The gross anatomical lesions of the ducklings showed an enlarged liver (hepatomegaly) and pleural exudates with yellow discoloration 48 hpi and hepatomegaly with brittle texture, plaque bleeding and necrosis, red darken splenomegaly, patchy hemorrhagic necrosis, bursal necrosis, renal bleeding, enlarged heart and inflated intestine 72 hpi (Fig. 6).

Macroscopic lesions of NDRV affected 1-day-old ducklings. (A) liver and spleen in physiological saline-injected ducklings; (B) hepatomegaly and pleural exudates with yellow discoloration 48 hpi; (C) hepatomegaly with brittle texture and patchy hemorrhagic necrosis 72 hpi; (D) red darken splenomegaly, enlarged heart and inflated intestine 72 hpi.
Twenty eight samples were collected at 24, 48 and 72 h, respectively from different affected organs, including heart, liver, spleen, lung and brain as well as anal swab (sticky stool) and serum were collected. Among the 21 NDRV LAMP-positive specimens, 18 were positive RT-PCR (Table 4). The RT-LAMP rapid detection assay was 10.7% (3/28) higher than that of the traditional RT-PCR method. Thus, the conventional RT-PCR method had low sensitivity and is inappropriate for use in diagnosis of NDRV.
Discussion
NDRV was isolated and identified from various duck species, for example, Muscovy, Mule, Peking and Sheldrake ducks18. The infected ducks revealed the fundamental clinical symptoms with hemorrhage and necrosis in the liver results in high mortality rates in ducks. In China, NDRV causes tremendous economic losses to the duck industry and most of the affected ducks, particularly ducklings died within 72 hpi. Therefore, it is crucially required to develop an easy, rapid, highly sensitive and specific diagnostic method for NDRV detection. In this study, we employed RT-LAMP to identify NDVR as a serious pathogen for duck industry in Guangdong province, China. Specificity of the RT-LAMP primers was investigated exploiting Muscovy Duck Reovirus (MDRV), Avian Reovirus (ARV), Duck Disease Virus (DPV), Duck Hepatitis Virus (DHAV), Duck Newcastle Disease Virus (NDV), H9 subtype avian influenza virus (AIV), H5 subtype AIV and duck Tanzuru virus (DTMUV). The utilization of the six specific primers that recognize distinct regions on the gene encoding the σB major outer-capsid protein of NDRV assured high specificity of the template nucleic acid amplification. Our data showed that there was no cross-reaction between NDVR and other examined viruses. The RT-LAMP specificity detection of NDRV was in accord with other studies about specificity of RT-LAMP reaction in viral detection7,8,9,10,19. The RT-LAMP assay in the detection of NDRV displayed sufficient sensitivity in addition to its higher specificity.
When the nucleic acid and plasmid quantities were checked by the RT-LAMP visualized by calcein assay, the detection limit was found to be 200 fg RNA, which was similar to that of RT-LAMP analyzed by agarose gel and RT-LAMP visualized by SYBR Green I. However, the detection limit of the conventional PCR (20 pg RNA). These results are in line with those of previous studies20,21,22,23,24. Interestingly, the current assay is more sensitive than the previously reported one16. Furthermore, the sensitivity of RT- LAMP in NDRV-infected ducklings (known) and field samples of ducks (suspected NDVR infection) was found to be higher than that of conventional PCR assay. Although the current study is based on a small number of experimental animals, RT-LAMP is a simple and powerful amplification method for the rapid diagnosis and early detection of experimentally NDVR- infected ducklings within 48 hpi.
The greater sensitivity is due to the high amplification efficiency of the RT- LAMP assay. Moreover, there is no time loss for thermal alteration under isothermal conditions in RT-LAMP. Others have reported that the enzymes used in LAMP are more resistant to inhibitory components in clinical samples25,26,27,28,29. Furthermore, it has been reported that PCR assay is more prone to PCR inhibitors in samples, which interfere with the amplification and sensitivity of PCR in comparison with LAMP assay30,31. We compared the performance of both calcein and SYBR Green I and our results indicated that both assays could be utilized to differentiate between positive and negative samples in visible or UV light.
The RT-LAMP-amplified products were detected by visual inspection employing calcein and SYBR Green I dyes, as well as by electrophoresis on agarose gels. As a result of the high amplification efficiency of LAMP31,32 and binding affinity of SYBR Green I to DNA33, the sensitivity of LAMP detection using calcein (the fluorescent emissions from calcein and the production of insoluble manganese/magnesium phosphate)34 and SYBR Green I was very high. The visual inspection result with calcein and SYBR Green I dyes was found to match with gel electrophoresis. Thereby, the visual inspection of LAMP amplified products by employing calcein and SYBR Green I dyes instead of gel electrophoresis made the RT-LAMP assay more rapid and simple.
This one-step RT-LAMP established in this study will provide an effective assay for the rapid diagnosis, surveillance and the examination of molecular epidemiology of NDRV for both developed and underdeveloped countries.
References
Bi, Z. et al. Induction of a robust immunity response against novel duck reovirus in ducklings using a subunit vaccine of sigma C protein. Scientific reports 6, 39092, https://doi.org/10.1038/srep39092 (2016).
Wozniakowski, G., Samorek-Salamonowicz, E. & Gawel, A. Occurrence of reovirus infection in Muscovy ducks (Cairina moschata) in south western Poland. Polish journal of veterinary sciences 17, 299–305 (2014).
Chen, Z., Zhu, Y., Li, C. & Liu, G. Outbreak-associated Novel Duck Reovirus, China, 2011. Emerging Infectious Diseases 18, 1209–1211, https://doi.org/10.3201/eid1807.120190 (2012).
Ge, Y. et al. Rapid and sensitive detection of novel avian-origin influenza A (H7N9) virus by reverse transcription loop-mediated isothermal amplification combined with a lateral-flow device. PloS one 8, e69941, https://doi.org/10.1371/journal.pone.0069941 (2013).
Su, C. W. et al. A novel application of real-time RT-LAMP for body fluid identification: using HBB detection as the model. Forensic science, medicine and pathology 11, 208–215, https://doi.org/10.1007/s12024-015-9668-6 (2015).
Lee, D. et al. Clinical evaluation of a loop-mediated isothermal amplification (LAMP) assay for rapid detection of Neisseria meningitidis in cerebrospinal fluid. PloS one 10, e0122922, https://doi.org/10.1371/journal.pone.0122922 (2015).
Chen, Q., Li, J., Fang, X. E. & Xiong, W. Detection of swine transmissible gastroenteritis coronavirus using loop-mediated isothermal amplification. Virology journal 7, 206, https://doi.org/10.1186/1743-422x-7-206 (2010).
Zhang, F. et al. A simple and rapid identification method for newly emerged porcine Deltacoronavirus with loop-mediated isothermal amplification. Biological research 50, 30, https://doi.org/10.1186/s40659-017-0135-6 (2017).
Ren, X. & Li, P. Development of reverse transcription loop-mediated isothermal amplification for rapid detection of porcine epidemic diarrhea virus. Virus genes 42, 229–235, https://doi.org/10.1007/s11262-011-0570-3 (2011).
Calvert, A. E., Biggerstaff, B. J., Tanner, N. A., Lauterbach, M. & Lanciotti, R. S. Rapid colorimetric detection of Zika virus from serum and urine specimens by reverse transcription loop-mediated isothermal amplification (RT-LAMP). PloS one 12, e0185340, https://doi.org/10.1371/journal.pone.0185340 (2017).
Xu, W. & Coombs, K. M. Avian reovirus L2 genome segment sequences and predicted structure/function of the encoded RNA-dependent RNA polymerase protein. Virology Journal 5, 153–153, https://doi.org/10.1186/1743-422X-5-153 (2008).
Borodavka, A., Ault, J., Stockley, P. G. & Tuma, R. Evidence that avian reovirus σNS is an RNA chaperone: implications for genome segment assortment. Nucleic Acids Research 43, 7044–7057, https://doi.org/10.1093/nar/gkv639 (2015).
Zhang, Y. et al. Characterization of the sigmaB-encoding genes of muscovy duck reovirus: sigmaC-sigmaB-ELISA for antibodies against duck reovirus in ducks. Veterinary microbiology 121, 231–241, https://doi.org/10.1016/j.vetmic.2006.12.008 (2007).
Kort, Y. H., Bourogaa, H., Gribaa, L., Hassen, J. & Ghram, A. Genotyping and Classification of Tunisian Strains of Avian Reovirus using RT-PCR and RFLP Analysis. Avian Dis 59, 14–19 (2015).
Park, S. I. et al. Immune response induced by ppGpp-defective Salmonella enterica serovar Gallinarum in chickens. Journal of microbiology (Seoul, Korea) 48, 674–681, https://doi.org/10.1007/s12275-010-0179-6 (2010).
Li, Z. L. et al. Development and evaluation of a reverse transcription-loop-mediated isothermal amplification method for rapid detection of novel duck reovirus. J. Anim. Vet. Adv. Journal of Animal and Veterinary Advances 11, 2756–2761 (2012).
Gou, H. et al. Rapid and sensitive detection of type II porcine reproductive and respiratory syndrome virus by reverse transcription loop-mediated isothermal amplification combined with a vertical flow visualization strip. J Virol Methods 209, 86–94, https://doi.org/10.1016/j.jviromet.2014.09.011 (2014).
Li, N. et al. The pathogenicity of novel duck reovirus in Cherry Valley ducks. Veterinary microbiology 192, 181–185, https://doi.org/10.1016/j.vetmic.2016.07.015 (2016).
Li, Q. et al. Rapid detection of porcine reproductive and respiratory syndrome virus by reverse transcription loop-mediated isothermal amplification assay. J Virol Methods 155, 55–60, https://doi.org/10.1016/j.jviromet.2008.09.012 (2009).
Budziszewska, M., Wieczorek, P. & Obrępalska-Stęplowska, A. One-step reverse transcription loop-mediated isothermal amplification (RT-LAMP) for detection of tomato torrado virus. Archives of Virology 161, 1359–1364, https://doi.org/10.1007/s00705-016-2774-2 (2016).
Hu, S. F. et al. Development of reverse-transcription loop-mediated isothermal amplification assay for rapid detection and differentiation of dengue virus serotypes 1-4. BMC microbiology 15, 265, https://doi.org/10.1186/s12866-015-0595-1 (2015).
Li, S. et al. Simultaneous detection and differentiation of dengue virus serotypes1-4, Japanese encephalitis virus and West Nile virus by a combined reverse-transcription loop-mediated isothermal amplification assay. Virology journal 8, 360, https://doi.org/10.1186/1743-422x-8-360 (2011).
Wheeler, S. S. et al. Surveillance for Western Equine Encephalitis, St. Louis Encephalitis and West Nile Viruses Using Reverse Transcription Loop-Mediated Isothermal Amplification. PloS one 11, e0147962, https://doi.org/10.1371/journal.pone.0147962 (2016).
Wang, X. et al. Rapid and sensitive detection of Zika virus by reverse transcription loop-mediated isothermal amplification. J Virol Methods 238, 86–93, https://doi.org/10.1016/j.jviromet.2016.10.010 (2016).
Priye, A. et al. A smartphone-based diagnostic platform for rapid detection of Zika, chikungunya and dengue viruses. Scientific reports 7, 44778, https://doi.org/10.1038/srep44778 (2017).
Elvira-Gonzalez, L. et al. Fast detection of Southern tomato virus by one-step transcription loop-mediated isothermal amplification (RT-LAMP). J Virol Methods 241, 11–14, https://doi.org/10.1016/j.jviromet.2016.12.004 (2017).
Francois, P. et al. Robustness of a loop-mediated isothermal amplification reaction for diagnostic applications. FEMS immunology and medical microbiology 62, 41–48, https://doi.org/10.1111/j.1574-695X.2011.00785.x (2011).
Hu, Y., Xu, P., Luo, J., He, H. & Du, W. Absolute Quantification of H5-Subtype Avian Influenza Viruses Using Droplet Digital Loop-Mediated Isothermal Amplification. 89, 745–750, https://doi.org/10.1021/acs.analchem.6b03328 (2017).
Kaneko, H., Kawana, T., Fukushima, E. & Suzutani, T. Tolerance of loop-mediated isothermal amplification to a culture medium and biological substances. Journal of biochemical and biophysical methods 70, 499–501, https://doi.org/10.1016/j.jbbm.2006.08.008 (2007).
Wilson, I. G. Inhibition and facilitation of nucleic acid amplification. Applied and environmental microbiology 63, 3741–3751 (1997).
Mori, Y., Nagamine, K., Tomita, N. & Notomi, T. Detection of loop-mediated isothermal amplification reaction by turbidity derived from magnesium pyrophosphate formation. Biochemical and biophysical research communications 289, 150–154, https://doi.org/10.1006/bbrc.2001.5921 (2001).
Notomi, T. et al. Loop-mediated isothermal amplification of DNA. Nucleic acids research 28, E63 (2000).
Karlsen, F., Steen, H. B. & Nesland, J. M. SYBR green I DNA staining increases the detection sensitivity of viruses by polymerase chain reaction. J Virol Methods 55, 153–156 (1995).
Tomita, N., Mori, Y., Kanda, H. & Notomi, T. Loop-mediated isothermal amplification (LAMP) of gene sequences and simple visual detection of products. Nature protocols 3, 877–882, https://doi.org/10.1038/nprot.2008.57 (2008).
Acknowledgements
This work was supported by Guangdong University Students, Science and Technology Innovation and Cultivate Special Funds (pdjh2017b0534) and the Start-up Research Grant Program provided by Foshan University, Foshan city, Guangdong province for distinguished researchers.
Author information
Authors and Affiliations
Contributions
Z.L., Y.C. and S.H. designed this project Z.L., Y.C., S.H., G.L., S.E., M.M., W.H., X. L., W.L. and C.H. performed the experiments and revised the manuscript.
Corresponding authors
Ethics declarations
Competing Interests
The authors declare no competing interests.
Additional information
Publisher's note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Li, Z., Cai, Y., Liang, G. et al. Detection of Novel duck reovirus (NDRV) using visual reverse transcription loop-mediated isothermal amplification (RT-LAMP). Sci Rep 8, 14039 (2018). https://doi.org/10.1038/s41598-018-32473-4
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-018-32473-4
Keywords
This article is cited by
-
Development and application of a multiplex qPCR assay for the detection of duck circovirus, duck Tembusu virus, Muscovy duck reovirus, and new duck reovirus
Virus Genes (2023)
-
DDX1 from Cherry valley duck mediates signaling pathways and anti-NDRV activity
Veterinary Research (2021)
-
Novel duck reovirus exhibits pathogenicity to specific pathogen-free chickens by the subcutaneous route
Scientific Reports (2021)
-
Using loop-mediated isothermal amplification for visual detection of porcine parvovirus 7
3 Biotech (2021)
-
High-mobility group box 1 protein (HMGB1) from Cherry Valley duck mediates signaling pathways and antiviral activity
Veterinary Research (2020)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
