
Abstract
Interspecific competition for resources leading to niche partitioning is considered as one of the major drivers of community assembly. Competitive niche partitioning is diagnosed from species co-occurrence, species abundance distributions (SADs), and body size distributions of species. For several decades, studies have explored these patterns for the relative significance of interspecific competition in shaping communities. We explored these patterns in a finite assemblage of insectivorous lizards in the Andaman & Nicobar Islands, both at the level of archipelago and individual islands. Negative geographic co-occurrences occurred only between species pairs in islands separated by deep ocean channels. Ecologically similar species did not show positive co-occurrence in guild co-occurrence analyses, indicating that the negative geographical co-occurrences between species in islands were due to historical allopatry. Species abundance distribution was best explained by a Pareto distribution in both metacommunity and local communities. There was no predictable spacing of body sizes among co-existing species in local communities. The empirical data on insular lizard community on species co-occurrence, SADs, and body size ratios does not lend support to assortment of species in islands caused by niche subdivision. Such niche-dissociated assembly of species in islands might be an important factor in formation of biological communities, regardless of geographic scale.
Similar content being viewed by others


Introduction
A long-standing paradigm in ecology is the assembly of biological communities through niche partitioning among species1,2. For the last 50 years, ecologists have largely used the Hutchinsonian niche concept1,3,4,5, and to a lesser extent, models based on stochastic processes, to explain the assembly of communities3,4,5,6,7. It has become increasingly evident that the complexity observed in biological communities can be the result of multiple processes acting at various temporal and spatial scales. This led to a unified theory of biodiversity that applies across such temporal and spatial scales3,4,8. This theory assumed per capita equivalence of individuals in a community (i.e. assuming no competition) and demonstrated that biological communities could be assembled based on random ecological drift, births, and deaths8,9,10. Despite this, competitive niche division remains the most dominant paradigm used to explain the coexistence of species (or the lack of it) in ecological communities11,12,13,14,15.
Niche division resulting from competition among species is expected to produce several emergent patterns in communities. Prominent among these are: negative co-occurrence patterns, species-abundance distributions, and constant size ratios of co-existing species. At regional scales, competition theory predicts that species with similar ecological requirements (or niches) may exclude each other, and co-occur less than expected by chance. The empirical evidence for this well-studied pattern is inconclusive and inferences ambiguous16,17,18,19,20,21,22,23,24,25,26,27. At the local community scale, patterns in species abundance distributions (hereafter abbreviated as SAD) inform about the nature of assembly and structure of communities1,28,29,30,31,32. Though many statistical and neutral models have attempted to explain the ‘hollow curve’ of SADs, mechanistic models based on niche partitioning have dominated the discussions9,29,32,33. Another expectation of interspecific competition is the partitioning of body sizes among species in a community. Strong competition among similarly sized species would cause character displacement to reduce size overlap (or reduced variability in size ratios), or hamper immigration of a species having similar body size into the community1,34,35,36. Therefore, niche partitioning should produce communities in which species are less similar to each other in body size than expected by chance.
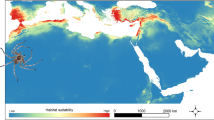
We tested competitive niche partitioning in a finite, indigenous, insectivorous lizard community in the Andaman & Nicobar Islands (Fig. 1), by examining patterns of species co-occurrence, SAD patterns, and body size ratios. We also examined the role of competition in structuring the lizard community at two geographic scales: (i) the archipelago or regional metacommunity and (ii) an island or local community. We used datasets on (i) species occurrences at the archipelago scale, (ii) species-abundance distributions for metacommunity and local communities, and (iii) body size ratios of co-existing species in local communities, to investigate the role of competitive niche partitioning in the assembly of this community. We show that from individual islands to archipelago, stochastic and historical factors were more important in the assembly of this insular lizard community. These findings advance our understanding of assembly of island communities, by proposing niche-dissociated processes as an important mechanism. It provides impetus for evidence-based conservation planning in the islands.

The Andaman & Nicobar Islands (ANI). The Andaman Islands (AND) are to the north of Ten Degree Channel and the Nicobar Islands (NIC) are to the south of the channel.
Results
We recorded 29 out of the 31 species of indigenous insectivorous lizards known from the ANI, the remaining two species are known only from museum records. Eighteen species were endemic to ANI, with eight species endemic to AND island group, and eight to NIC island group. In bounded quadrats, we recorded 850 individuals and 14 species. We collected body mass data from 875 lizards belonging to 29 species. Similar positive species-area relationships were observed in ANI (slope z = 0.19, R2 = 0.66, F = 51.74, df = 27), AND (slope z = 0.20, R2 = 0.74, F = 33.33, df = 12), and NIC (slope z = 0.18, R2 = 0.51, F = 13.75, df = 13).
In ANI, nearly half of all species pairs showed non-random (either positive or negative) species co-occurrence (Fig. 2a) (Table 1). However, more species pairs showed random co-occurrence than either positive or negative interactions (Fig. 2a) (Table 1). Either the Ten Degree Channel or the smaller channels that separated the island groups in the Nicobar Islands delineated all species pairs showing negative species co-occurrence. Both AND and NIC communities had low proportions of species pairs showing non-random co-occurrence (Fig. 2b,c) (Table 1). There were no species pairs in AND that showed negative species co-occurrence (Fig. 2b) (Table 1). Only 12 pairwise interactions in AND were non-random, all of which were positive species co-occurrences (Table 1). In NIC, species pairs showed both positive and negative species co-occurrence (Fig. 2c) (Table 1). All significant negative co-occurrence occurred between pairs of species that inhabited distinct island groups within NIC (Fig. 2c). In addition, all positive co-occurrences were between species that inhabited the same island group (Fig. 2c). In both AND and NIC, the majority of interactions between species pairs were random or unclassifiable (Fig. 2b,c) (Table 1). The negative co-occurrences in ANI was due to turnover of species between AND and NIC lizard communities across the Ten Degree Channel. The probabilities of co-occurrence of individual species pairs and the standardized effect sizes in ANI, AND, and NIC are summarized in Supplementary File 5(a,b and c). Sphenomorphus maculatus (SPM) was the only species that occurred in both AND and NIC and showed significant co-occurrence with six other species in ANI (Fig. 2a) (Supplementary File 5a). Among these six significant co-occurrences, only two were negative – with Bronchocela danieli (BRD) & Gekko smithii (GES). Both these species occur in the southern group of Nicobar Islands, which are separated from the rest of the Nicobar Islands by the Sombrero Channel (depth > 150 m). All other species pairs that showed negative co-occurrence (shaded grey in Supplementary File 5) occurred on island groups separated by deep ocean channels, i.e., Ten Degree Channel (depth > 1000 m) and Sombrero Channel (depth > 150 m). In addition, all positive co-occurrences were between species that occurred within groups of islands separated by shallow water, such as the islands within AND separated by sea less than 50 m deep and the islands in the central group in NIC (Fig. 2). In within guild pairwise interactions, no significant pairwise interactions among species in ANI, AND, and NIC communities were observed (Table 1). Since all classified interactions in the guild co-occurrence analysis were random (Table 1), comparing the two co-occurrence analyses using a contingency table approach revealed that significant negative co-occurrences in the islands are the result of historical allopatry37.

Species co-occurrence matrix for lizards in: (a) Andaman & Nicobar Islands (ANI) (b) Andaman Islands (AND) (c) Nicobar Islands (NIC). Species that showed only random or unclassifiable interactions are excluded from this matrix.
One species occurring at very high abundances and absence of rare species was characteristic of all observed SADs (Fig. 3). We examined the fit of fourteen species-abundance and relative abundance models to one metacommunity (AND) and seven local communities. Table 2 shows the top three models for AND, and two local communities (LAND and GNI) (Details of all models and communities are in Supplementary File 6). Pareto distribution (\(f(x)=\frac{b{a}^{b}}{{x}^{(b+1)}}\) for all x >= scale, and f(x) = 0 otherwise, where a = scale and b = shape) fit all the island communities well, though broken-stick model (p(x) = \(\frac{S-1}{N}{(1-\frac{x}{N})}^{S-2}\), where p(x) = probability density for a given abundance ‘x’, S = species richness, and N = number of individuals) also fit observed SADs for ANI, SA, RUT, NEIL, and CAM (Table 2, Supplementary File 6). Both the metacommunity and local communities exhibited similar SAD patterns (Table 2, Supplementary File 6). Examination of predicted abundance against observed values showed that pareto model best fit the observed abundance distributions. Broken-stick model fit mid-portions of the distributions (Fig. 3). Therefore, both the statistical pareto distribution and broken-stick model explained SAD patterns at different ecological scales in lizards of the Andaman & Nicobar Islands.

Species abundance distributions in: (a) Andaman Islands (AND) (b) Little Andaman Island (LAND) (c) Great Nicobar Island (GNI). Only the top three models from a ranking of models based on AIC values are presented here.
Insectivorous lizards ranged in body mass from 0.5 g (Cnemaspis sp.) to 84 g (Eutropis rudis). Species richness in individual islands ranged from four to 14 species (Table 3). Observed V-ratios (variance in size ratios of co-existing lizards) varied in the 23 islands sampled from 0.15 to 13.23. There was a significant negative correlation between species richness in a community and observed V-ratios (Pearson’s product-moment correlation, R = −0.43, t = −2.17, df = 21, P = 0.04). When Chester Island, which had a large observed V-ratio (13.23) was removed from the analysis, the correlation became insignificant (R = −0.28, t = −1.33, df = 20, P = 0.20). Most islands did not show significant difference between observed and simulated V-ratios (Table 3, Supplementary File 7). In several islands, (e.g., South Andaman Island, Great Nicobar Island, Tarmugli Island, & Car Nicobar Island, see Supplementary File 7) the V-ratios expected under the null model of community assembly were similar to observed V-ratios. Since body sizes of co-existing lizards did not show constant ratios, it suggests a random assortment of species based on body sizes in the Islands.
Discussion
If interspecific competition influenced the probability of a species being present or absent in an island, then species pairs that showed negative geographical co-occurrence should show significant positive guild co-occurrence. We observed the highest number of pairwise negative species co-occurrences in ANI. None of these species pairs showed a significant positive guild co-occurrence. In NIC, species pairs that showed negative geographic co-occurrence did now show any positive guild co-occurrence. Therefore, we infer that negative species co-occurrences detected in ANI and NIC are the result of ‘historical allopatry’37. These species or their progenitors independently invaded these island groups and never came in to contact with each other. Two factors that contribute to allopatry in this case could be limited dispersal abilities of species and geographic barriers. In the case of ANI, 95% of negative co-occurrences were between species that occur on either side of the Ten Degree Channel. It is more than 1000 m deep and approximately 140 km wide and serves as a barrier maintaining historical allopatry. In AND, where almost all islands are separated by shallow sea, no significant negative co-occurrence was shown by any species pair (Figs 1 and 2b)38,39,40. Consequently, lowering of sea levels would have in the past created land connections between these islands. A few species pairs showed negative co-occurrence in NIC, where groups of islands are separated by channels shallower and narrower than the Ten Degree Channel, yet deep enough for them to be separated even during the lowest Pleistocene sea levels38,41. In NIC, negative co-occurrences were between species pairs occurring in islands separated by the Sombrero Channel that is more than 200 m deep (Nancowry group and the southern group of islands)38,41. This would have kept these island groups separated from each other even during maximum lowering of sea levels during the Pleistocene38,41. Therefore, we hypothesize that distribution patterns of lizards in these islands are determined by barriers to dispersal rather than interspecific competition. The positive co-occurrences opened the question of whether these are true biological associations or a result of ‘common history’37. On a regional scale, trait overdispersion and habitat filtering should cause species with differing ecologies to show more positive co-occurrence42,43. In this case, species pairs that showed positive co-occurrence in geographical matrices should have shown corresponding negative co-occurrence in the guild matrices. However, guild based co-occurrence analysis showed no significant negative interactions regardless of the geographic scale (ANI, AND, or NIC). This suggests that ecological interactions between species did not cause species to co-occur in islands. Since all the classified ecological interactions turned out to be random, the positive co-occurrences in geographical matrices can only be the result of common history37. In AND where most of the positive co-occurrences occurred, current species compositions in individual islands are the result of fragmentation of a larger land mass (which seemingly obtained its fauna from Southeast Asia) at the end of Pleistocene39,40,44.
Many recent studies have failed to detect any evidence of interspecific competition determining species co-occurrence22,37,45,46,47. A meta-analysis of several presence-absence matrices showed that in general, birds, mammals, ants, and plants showed non-random species co-occurrence while fishes, herpetofauna, and several invertebrates did not48. Even congeneric species – which are often thought to have similar ecological requirements and likely to exhibit negative species co-occurrence – did not exhibit non-random patterns of species co-occurrence37,49. Interspecific competition in insular herpetofaunal communities might not be strong enough to competitively exclude species from islands creating negative co-occurrence patterns22. With several macroecological datasets, including ours, now available for drawing broad generalizations, negative co-occurrence resulting in competitive niche division between closely related species is probably an exception, rather than a rule22.
Species abundance distributions from continents often have long negatively skewed tails composed of rare species9. This is missing from the Andaman & Nicobar lizard communities. In continental communities, many species are considered rare primarily due to their low detection probabilities50. The idea of ‘veil of rarity’ was based on the observation that several species in communities are rare and not sampled easily51,52,53. However, island communities typically have low number of species, and it is possible to sample the entire community. For terrestrial herpetofauna, bounded quadrats used for sampling reptiles provided counts of all individuals in the sampled area, thus eliminating the possibility of major differences in detection probabilities of species, which may distort patterns in relative species abundance54,55,56,57. Therefore, the absence of a long tail of rare species in the relative abundance distribution in this case, is not a result of insufficient sampling or imperfect detection probabilities of species in the islands. Rather, it is due to enhanced threshold probability extinction of rare species in the islands.
Explaining patterns in the abundance of species gained momentum with mechanistic models that accounted for resource partitioning by species. In this study, pareto distribution was the best SAD model consistently across all communities, though the broken-stick model also fit the metacommunity (ANI) and some of the local communities. The consistency of the fit of pareto distribution to both local communities and metacommunity showed that SAD did not change from local communities to metacommunity, exhibiting a fractal nature of islands in this archipelago. The pareto distribution is a continuous power-law density distribution that was originally used in modelling distribution of personal incomes in countries but was introduced in community ecology in an early attempt to describe SAD58. The similarity between distributions in non-biological systems and SAD has been pointed out earlier, but rarely been discussed in ecology58,59. SAD like patterns are not a unique property of ecological communities, but are common to many complex systems60. If ecological communities are like other non-biological complex systems, then it might be necessary to use other approaches to further explain the most commonly observed SADs60,61. Nekola & Brown have suggested that perhaps ecological SADs may only be explained in a post-hoc manner (e.g. examining the variation in abundances in relation to local environmental factors or other resources), and that only qualitative predictions may be possible about their nature60.
If body size is important in determining the niches of species, then the division of body size in communities should predict a minimum size difference between ecologically similar co-existing species1. Though there is not much empirical evidence for a threshold size difference62,63,64, some studies have suggested that the spacing of body sizes between co-existing species is non-random, as documented in desert rodents and local communities of bog ants65,66. The insectivorous lizards in the Andaman & Nicobar Islands varied in size greatly (0.5 g to 84 g). However, there was no predictable spacing of body sizes between co-existing lizard species. Our results also show that body size difference between pairs of co-existing species need not be constant either within, or between communities, of varying sizes. Stochastic species extinctions also have undoubtedly contributed to the random variation in body size differences among these lizards. These outcomes are possible when the community of lizards was assembled in the islands by passive random sampling from a metacommunity without any niche apportionment.
Ecologically similar co-existing species competing for resources and their niches differing from each other to reduce competition is a dominant paradigm in community ecology. However, we found no evidence of niche partitioning affecting the emergent properties of insular lizard communities, whether one looked at regional (species co-occurrence) or local community (SAD and size ratios) structure. Our results point at the role of historical and stochastic events in the assembly of insectivorous indigenous lizards in the Andaman & Nicobar Islands. These results, and those of several other recent studies on other taxa, downsize the role of competitive interactions among species in the assembly of communities5,67,68,69. Studies on community assembly may benefit from borrowing the methods of complexity sciences and emphasizing the recognition of potentially common underlying factors60.
Global lizard distribution patterns do not completely overlap with that of other vertebrates70. Therefore, conservation actions should take into consideration the distinctive ecological and evolutionary processes that have shaped the distribution of lizards70. The community of lizards that occupy the Andaman & Nicobar Islands is the outcome of processes of natural selection during their evolutionary history. Now, their persistence in these islands is fraught with human-induced habitat loss and biological invasions71 that will hasten their extinction. With the significant turnover of species in the archipelago (between the Andaman Islands and the Nicobar Islands; within the Nicobar Islands), any attempt to conserve species through protected areas will have to devote equal effort in all the island groups. The existing protected areas in the Andaman Islands are extensive, with over 80% of terrestrial habitats under protection. However, it is not evenly represented in all island groups. The Nancowry group, in particular, has several endemic reptiles. However, it has only one protected area in one small (~17 km2) island. Since geographical barriers to dispersal are the primary constraints on species distribution in the islands, conservation efforts will have to be more widespread on multiple islands and island groups.
Methods and Materials
Study area
The Andaman & Nicobar Islands consists of 556 islands, islets, and rocks, covering 8249 km2, located in the eastern part of the Bay of Bengal44 (Fig. 1). These islands form a continuous chain of mountains sprawling in a great arc between Cape Negrais of Myanmar and Achin Head of Sumatra, about 155 km south-east of Great Nicobar Island. It is a part of the Great Alpine-Himalayan System41. Paleo plate reconstructions of Southeast Asia indicate that the emergence of these islands above sea level happened only during the late Miocene (10 million years before present)40. While the Nicobar Islands appear to be truly oceanic in nature, surrounded by deep channels, the possibility that the Andaman Islands at their northern tip might have been connected to mainland Asia cannot be ruled out72. The mean annual rainfall in these islands exceeds 3000 mm, supporting a predominantly tropical evergreen vegetation73. The Andaman Islands are a part of the Indo-Burma biodiversity hotspot and the Nicobar Islands are part of the Sundaland biodiversity hotspot74. Barren and Narcondam islands are volcanic, outlying islands towards the east of the main island archipelago, and not included in this study. Many previous authors have described these islands and their fauna in detail38,41,44,72.
The lizard fauna of Andaman & Nicobar Islands as currently known, consists of 31 species of indigenous insectivorous lizards, one species of insectivorous/carnivorous lizard (Varanus salvator), and three introduced species (Hemidactylus frenatus, Hemidactylus cf. brookii, and Calotes versicolor). Only the former 31 species are considered here for further analysis as V. salvator occupies a different trophic level and the latter three introduced species are found only in association with human habitation. Biogeographically, the reptile fauna of the Andaman Islands has Indochinese affinities, while that of the Nicobar Islands has Malayan-Sundaland affinities44,75.
Sampling lizards
We carried out intensive surveys in the Andaman & Nicobar Islands from March 2010 to January 2014, avoiding the heavy monsoon during the months of June-September. Species presence-absence was determined in 29 islands using visual encounter surveys, opportunistic records, museum records, and past publications records44,54,76,77,78,79,80,81. For estimating abundance, we sampled bounded quadrats of dimensions 10 m × 10 m, obtaining total counts of all individuals in the sampled quadrats54,55,82. We sampled 49 bounded quadrats in 14 islands the Andaman Islands (AND), with 10 of these in Little Andaman Island (LAND)54. In the Nicobar Islands (NIC), we sampled ten bounded quadrats in Great Nicobar Island (GNI) and four in Camorta Island (CAM). Since quadrats did not sample canopy-living species efficiently, we removed occasional records of such species from the data prior to analysis (five records out of 855 observations of lizards).
Datasets
We recorded 29 of the 31 indigenous species of lizards occurring in the Andaman & Nicobar Islands (ANI). The remaining two species (Scincella macrotis and Lipinia macrotympanum) are known only from historical records, but with accurate locality records78,83. Using these 31 species, we created three ‘geographic co-occurrence’ matrices: all islands sampled in the Andaman and Nicobar Archipelago (ANI, 29 islands), the Andaman Islands (AND, 14 islands), and the Nicobar Islands (NIC, 15 islands), with species in rows and islands in columns (Supplementary File 1). AND & NIC are nested within the larger ANI community. Abbreviations for species and island names are given in Supplementary File 1. Using natural history observations and literature, we classified the lizards into four guilds based on habitat preference (Arboreal or Terrestrial) and diel activity pattern (Diurnal or Nocturnal), creating a second ‘guild co-occurrence’ matrix with species in rows and guild names in columns (Supplementary File 2).
Quadrats sampled 14 species of forest floor and understorey species from the ANI (850 individuals). Since all islands sampled in the Andaman Islands had similar lizard communities with a high degree of nestedness in species composition, we pooled this data as Andaman Islands community (AND) to estimate average abundance of a species (number of individuals/100 m2). For the seven individual island communities analysed – Little Andaman Island (LAND), South Andaman Island (SAND), Rutland Island (RUT), North Andaman Island (NAND), Neil Island (NEIL), Camorta Island (CAM), and Great Nicobar Island (GNI) – we estimated local abundance of lizards using only samples from these islands. Thus, we created eight data sets with species ranked in descending order of abundance (Supplementary File 3).
We collected body mass data for 29 species from 875 adult individuals (Supplementary File 4). Scincella macrotis, (known from specimens collected in the 19th century from a single location in Great Nicobar Island) and Hemidactylus garnotii were not used in analysis of body size, as we could not collect reliable size and mass information on these species. For every species, we recorded body weight in grams (W) for multiple individuals (875 individuals) using a Pesola™ Spring Balance with 0.1 g accuracy. We confirm that all methods were carried out in accordance with relevant guidelines and regulations of the Department of Environment & Forests, Andaman & Nicobar Islands Wildlife. The study was approved by the Science and Engineering Research Board, Department of Science and Technology, India. All methods were approved by the Training Research Advisory Committee of the Wildlife Institute of India.
Data analysis
Since the islands were of varying sizes, we needed to assess potential intrinsic differences between these islands in their habitat suitability, ease of colonization, historical factors etc84. For this, we compared the slopes (z) of the well-known species-area relationships (the positive relationship between species richness and island areas) in ANI, AND, & NIC, using a linear least squares regression of log island area against log species richness. Since there was a high degree of overlap between communities of different islands, we assumed that any major variation in ‘z’ would be a result of intrinsic differences between islands
To explore patterns in the co-occurrence of lizards, we used a probabilistic analysis proposed by Veech85, by calculating the probability that two species co-occur less than or greater than the observed frequency of co-occurrence85,86,87. In a geographic co-occurrence matrix (species × islands), we counted any pair of species occurring together in the same island as a positive co-occurrence, and occurrence of only one of a pair of species in any given island as a negative co-occurrence. To examine whether dispersal limitation across biogeographical barriers would explain species co-occurrence patterns, we conducted three analyses on three presence-absence datasets: Andaman & Nicobar Islands Archipelago (ANI), the Andaman Islands (AND), and the Nicobar Islands (NIC). For ANI, we expected that the Ten Degree Channel, a deep-sea barrier between the Andaman Islands and the Nicobar Islands, would produce a significant number of negative co-occurrences purely due to limited dispersal between these groups. In AND, dispersal limitation would be a poor explanation for negative co-occurrence between species pairs as there are no major geographic barriers between the islands sampled, and interspecific interactions may be of importance38,41. In NIC, there are geographical barriers (channels deeper than 150 m below current sea level) between island groups (northern group, central group, and southern group)38,41. If negative co-occurrence patterns occur between pairs of species inhabiting different island groups in the NIC, it could be the result of dispersal limitation, while any negative co-occurrence between species inhabiting the same group could due to interspecific interactions. If competition is a causal factor, for species pairs that show significant negative geographic species co-occurrence, one would expect positive guild co-occurrence in a species × guild matrix, as competing species are expected to be ecologically similar to each other37.
For these analyses, we chose a minimum threshold of one for calculating expected species co-occurrence i.e., the two species may co-occur in at least one island. Species pairs that had expected co-occurrence less than one were removed from the analysis85. Significant positive co-occurrences are those where pairs of species co-exist in more sites than expected, while significant negative co-occurrences are those where they co-exist in fewer sites than expected (keeping α = 0.05)85. Random associations are those in which the observed number of co-occurrences did not deviate from their expected values by more than 0.1 times the total number of sites85. Standardized effect sizes were calculated as observed – expected value divided by total number of sites (range −1 to 1)85. We first analysed pairwise geographic species co-occurrence (species × island). To ascertain the causal factors behind the observed patterns, we conducted a second set of analyses on guild species co-occurrence for ANI, AND, & NIC (species × guild)37. The results of geographical and guild co-occurrence analyses were compared using an interaction matrix proposed by Sfenthourakis37. All analyses were conducted in programme R using the package ‘cooccur’87,88. Of the 465 species pair combinations in ANI, we removed 227 pairs (48.82%) from the analysis because their expected co-occurrence was <1. Similarly, we removed 27 (22.5%) out of 120 pairs in AND and 109 (57.37%) out of 190 pairs in NIC from the analysis. Thus, we analyzed 238 species pairs in ANI, 93 in AND, and81 in NIC.
We examined species abundance distributions (SAD) by fitting fourteen well-known models of species-abundance/rank-abundance to observed data using a Maximum Likelihood Estimation based fitting procedure88,89. The models examined here are: pareto distribution90, broken-stick model30, geometric series91, log-series92, lognormal51, Weibull distribution93, Power-discreet distribution90, Zipf distribution90, Zipf-Mandelbrot distribution94, zero-sum multinomial distribution95, poisson lognormal96, neutral model97, negative binomial98, and gamma distribution99. We used Akaike Information Criteria (AIC) for ranking the models in descending order. We examined SADs for terrestrial lizards in the Andaman Islands (AND) which is a metacommunity, and seven local communities (individual islands) – LAND, SAND, RUT, NAND, NEIL, CAM, and GNI (for abbreviations, see Annexure 1). These analyses were conducted in programme R, using the package ‘SADs’88,89.
Ordered, log transformed body mass data was used to examine whether lizard communities exhibited a constant size ratio. We used the variance in size ratios (V-ratio) as a metric to examine the distribution of body sizes among species in 23 local communities (individual islands)100. We looked for non-random size ratios in these communities by comparing them with expected V-ratios in null communities having equal species richness. To create null communities, we first defined a source pool consisting of all species of insectivorous lizards in the ANI. We added an arbitrary five hypothetical species to this source pool to ensure that the source pool community always had more species than all local communities did. We kept the maximum body size in the source community identical to the largest body size exhibited by the real community. We drew null communities from this source pool and compared the V-ratios against observed local communities. We used Standardized Effect Size (SES) to evaluate the significance of difference between observed and simulated V-ratios. These analyses were performed using the R package EcosimR88,101.
References
Hutchinson, G. E. Homage to Santa Rosalia or Why are there so many kinds of Animals. Am. Nat. 93, 145–159 (1959).
Price, T. D. et al. Niche filling slows the diversification of Himalayan songbirds. Nature 509, 222–5 (2014).
Caswell, H. Community Structure: A Neutral Model Analysis. Ecol. Monogr. 46, 327–354 (1976).
Chesson, P. L. & Warner, R. R. Environmental variability promotes coexistence in lottery competitive systems. Am. Nat. 117, 923–943 (1981).
Connor, E. F. & Simberloff, D. The Assembly of species communities: Chance or Competition? Ecology 60, 1132–1140 (1979).
Gravel, D., Canham, C. D., Beaudet, M. & Messier, C. Reconciling niche and neutrality: the continuum hypothesis. Ecol. Lett. 9, 399–409 (2006).
Fisher, C. K. & Mehta, P. The transition between the niche and neutral regimes in ecology. Proc. Natl. Acad. Sci. 111, 13111–13116 (2014).
Hubbell, S. P. A unified theory of biogeography and relative species abundance and its application to tropical rain forests and coral reefs. Coral Reefs 16, S9–S21 (1997).
Hubbell, S. P. The Unified Neutral Theory of Biodiversity and Biogeography. Monographs in Population Biology 17, (Princeton University Press, 2001).
Abrams, P. A. A world without competition. Nature 412, 858–859 (2001).
Rosindell, J., Hubbell, S. P. & Etienne, R. S. The Unified Neutral Theory of Biodiversity and Biogeography at Age Ten. Trends Ecol. Evol. 26, 340–348 (2011).
Alonso, D., Etienne, R. S. & McKane, A. J. The merits of neutral theory. Trends Ecol. Evol. 21, 451–457 (2006).
Holt, R. D. Bringing the Hutchinsonian niche into the 21st century: Ecological and evolutionary perspectives. Proc. Natl. Acad. Sci. 106, 19659–19665 (2009).
Kraft, N. J. B., Valencia, R. & Ackerly, D. D. Functional traits and niche-based tree community assembly in an Amazonian forest. Science (80-.). 322, 580–2 (2009).
Letten, A. D., Ke, P.-J. & Fukami, T. Linking modern coexistence theory and contemporary niche theory. Ecol. Monogr. 87, 161–177 (2017).
Gotelli, N. J., Buckley, N. J. & Wiens, J. A. Co-Occurrence of Australian Land Birds: Diamond’s Assembly Rules Revisited. Oikos 80, 311–324 (1997).
Borthagaray, A. I., Arim, M. & Marquet, P. A. Inferring species roles in metacommunity structure from species co-occurrence networks. Proc. Biol. Sci. 281, 20141425 (2014).
Diamond, J., Pimm, S. L. & Sanderson, J. G. The checkered history of checkerboard distributions: comment. Ecology 96, 3386–3388 (2015).
Connor, E. F., Collins, M. D. & Simberloff, D. The checkered history of checkerboard distributions: reply. Ecology 96, 3388–3389 (2015).
Hastings, A. Can Competition Be Detected Using Species Co-Occurrence Data? Ecology 68, 117–123 (1987).
Gilpin, M. E. & Diamond, J. M. Factors contributing to non-randomness in species co-occurrences on islands. Oecologia 52, 75–84 (1982).
Pitta, E., Giokas, S. & Sfenthourakis, S. Significant pairwise co-occurrence patterns are not the rule in the majority of biotic communities. Diversity 4, 179–193 (2012).
Diamond, J. M. In Ecology and Evolution of Communities (eds Cody, M. L. & Diamond, J. M.) 342–444 (Harvard University Press, 1975).
Ulrich, W. Species co-occurrences and neutral models: reassessing J. M. Diamond’s assembly rules. Oikos 107, 603–609 (2004).
Stone, L. & Roberts, A. Competitive exclusion, or species aggregation? Oecologia 91, 419–424 (1992).
Heino, J. & Grönroos, M. Does environmental heterogeneity affect species co-occurrence in ecological guilds across stream macroinvertebrate metacommunities? Ecography (Cop.). 36, 926–936 (2013).
Connor, E. F., Collins, M. D. & Simberloff, D. The checkered history of checkerboard distributions. Ecology 94, 2403–2414 (2013).
Wiens, J. A. On size ratios and sequences in ecological communities: Are there no rules? Ann. Zool. Fennici 19, 297–308 (1982).
Tokeshi, M. Ninche apportionment or random assortment: species abundance patterns revisited. J. Anim. Ecol. 59, 1129–1146 (1990).
MacArthur, R. H. On the relative abundance of bird species. Zoölogy 43, 293–295 (1957).
MacArthur, R. On the relative abundance of species. Am. Nat. 94, 25–36 (1960).
Tokeshi, M. In Advances in Ecological Research 24, 111–186 (1993).
Ulrich, W., Ollik, M. & Ugland, K. I. A meta-analysis of species-abundance distributions. Oikos 119, 1149–1155 (2010).
Roth, V. L. Constancy in the Size Ratios of Sympatric Species. Am. Nat. 118, 394–404 (1981).
Kiltie, R. A. Size ratios among sympatric neotropical cats. Oecologia 61, 411–416 (1984).
Pagel, M. D. & Greenough, J. A. Explaining Species Size Ratios in Nature. Trends Ecol. Evol. 2, 114–115 (1987).
Sfenthourakis, S., Tzanatos, E. & Giokas, S. Species co-occurrence: The case of congeneric species and a causal approach to patterns of species association. Glob. Ecol. Biogeogr. 15, 39–49 (2006).
Rodolfo, K. S. Bathymetry and Marine Geology of the Andaman Basin, and Tectonic Implications for Southeast Asia. Geol. Soc. Am. Bull. 80, 1203–1230 (1969).
Voris, H. K. Maps of Pleistocene sea levels in Southeast Asia: shorelines, river systems and time durations. J. Biogeogr. 27, 1153–1167 (2000).
Lee, T. & Lawver, L. A. Cenozoic plate reconstruction of Southeast Asia. Tectonophysics 251, 85–138 (1995).
Karunakaran, C., Ray, K. K. & Saha, S. S. A revision of the stratigraphy of Andaman and Nicobar Islands, India. Bull. Natl. Inst. Sci. 38, 436–441 (1968).
Cavender-Bares, J., Kozak, K. H., Fine, P. V. A. & Kembel, S. W. The merging of community ecology and phylogenetic biology. Ecol. Lett. 12, 693–715 (2009).
Webb, C. O., Ackerly, D. D., McPeek, M. A. & Donoghue, M. J. Phylogenies and community ecology. Annu. Rev. Ecol. Syst. 33, 475–505 (2002).
Das, I. Biogeography of the amphibians and reptiles of the Andaman and Nicobar Islands, India. In Tropical Island Herpetofauna. Origin, Current Diversity, and Conservation (ed. Ota, H.) 43–77 (Elsevier Science B. V., 1999).
Luiselli, L. & Filippi, E. Null models, co-occurrence patterns, and ecological modelling of a Mediterranean community of snakes. Amphibia-Reptilia 27, 325–337 (2006).
França, F. G. R. & Araújo, A. F. B. Are there co-occurrence patterns that structure snake communities in Central Brazil? Braz. J. Biol. 67, 33–40 (2007).
Luiselli, L., Capula, M., Rugiero, L., Salvi, D. & Akani, G. C. Does interspecific competition with a stronger competitor explain the rarity of an endangered snake on a Mediterranean island? Ecol. Res. 27, 649–655 (2012).
Gotelli, N. J. & McCabe, D. J. Species co-occurrence: a meta-analysis of J. M. Diamond’s assembly rules model. Ecology 83, 2091–2096 (2002).
Cardillo, M. & Meijaard, E. Phylogeny and co-occurrence of mammal species on Southeast Asian islands. Glob. Ecol. Biogeogr. 19, 465–474 (2010).
MacKenzie, D. I. & Kendall, W. C. How should detection probability be incorporated into estimates of relative abundance? Ecology 83, 2387–2393 (2002).
Preston, F. W. The Commonness, And Rarity, of Species. Ecology 29, 254–283 (1948).
Preston, F. W. The Canonical distribution of commonness and rarity: Part I. Ecology 43, 185–215 (1962).
Nee, S., Harvey, P. H. & May, R. M. Lifting the Veil on Abundance Patterns. Proc. R. Soc. B Biol. Sci. 243, 161–163 (1991).
Harikrishnan, S. & Vasudevan, K. The devil is in the detail: estimating species richness, density, and relative abundance of tropical island herpetofauna. BMC Ecol. 2015, 18 (2015).
Rodda, G. H., Campbell, E. W. I. & Fritts, T. H. A high validity census technique for herpetofaunal assemblages. Herpetol. Rev. 32, 24–30 (2001).
Rodda, G. H. & Campbell, E. W. Distance Sampling of Forest Snakes and Lizards. Herpetol. Rev. 33, 271–274 (2002).
Smolensky, N. L. & Fitzgerald, L. A. Distance Sampling Underestimates Population Densities of Dune-Dwelling Lizards. J. Herpetol. 44, 372–381 (2010).
Preston, F. W. Pseudo-lognormal distributions. Ecology 62, 355–364 (1981).
Preston, F. W. Gas laws and wealth laws. Sci. Mon. 71, 309–311 (1950).
Nekola, J. C. & Brown, J. H. The wealth of species: Ecological communities, complex systems and the legacy of Frank Preston. Ecol. Lett. 10, 188–196 (2007).
Golestani, A. & Gras, R. A New Species Abundance Distribution Model Based on Model Combination. Int. J. Biostat. 9, 33–48 (2013).
Taper, M. L. & Case, T. J. In Oxford Surveys in Evolutionary Biology (eds Futuyma, D. & Antonovics, J.) 63–110 (Oxford University Press, 1992).
Simberloff, D. & Boecklen, W. Santa Rosalia Reconsidered: Size Ratios and Competition. Evolution (N. Y). 35, 1206–1228 (1981).
Losos, J. B., Naeem, S. & Colwell, R. K. Hutchinsonian Ratios and Statistical Power. Evolution (N. Y). 43, 1820 (1989).
Bowers, M. A. & Brown, J. H. Body size and coexistnce in desert rodents: chance or community structure? Ecology 63, 391–400 (1982).
Gotelli, N. & Ellison, A. Assembly rules for New England ant assemblages. Oikos 99, 591–599 (2002).
Simberloff, D. S. Using Island Biogeographic Distributions to Determine if Colonization is Stochastic. Am. Nat. 112, 713–726 (1978).
Simberloff, D. Competition Theory, Hypothesis-Testing, and Other Community Ecological Buzzwords. Am. Nat. 122, 626–635 (1983).
Connor, E. F. & Simberloff, D. S. Interspecific competition and species co-occurence patterns on islands: null models and the evaluation of evidence. Oikos 41, 455–465 (1983).
Roll, U. et al. The global distribution of tetrapods reveals a need for targeted reptile conservation. Nat. Ecol. Evol. 1, 1677–1682 (2017).
Mohanty, N. P., Harikrishnan, S., Sivakumar, K. & Vasudevan, K. Impact of invasive spotted deer (Axis axis) on tropical island lizard communities in the Andaman archipelago. Biol. Invasions 18, 9–15 (2016).
Ripley, S. D. & Beehler, B. M. Ornithogeographic affinities of the Andaman and Nicobar islands. J. Biogeogr. 16, 323–332 (1989).
Champion, H. G. & Seth, S. K. A Revised Survey of the Forest Types of India. (Natraj Publishers, Publication Division, 1968).
Myers, N., Mittermeier, R. A., Mittermeier, C. G., Da Fonseca, G. A. & Kent, J. Biodiversity hotspots for conservation priorities. Nature 403, 853–858 (2000).
Smith, M. A. The herpetology of the Andaman and Nicobar Islands. Proc. Linn. Soc. London 1940, 150–158 (1940).
Harikrishnan, S., Chandramouli, S. R. & Vasudevan, K. A survey of herpetofauna on Long Island, Andaman and Nicobar Islands, India. Herpetol. Bull. 2012, 19–28 (2012).
Das, I. The validity of Dibamus nicobaricum (Fitzinger in Steindachner, 1867) (Squamata: Sauria: Dibamidae). Russ. J. Herpetol. 3, 157–162 (1996).
Das, I. Rediscovery of Lipinia macrotympanum (Stoliczka, 1873) from the Nicobar Islands, India. Asiat. Herpetol. Res. 7, 23–26 (1997).
Biswas, S. & Sanyal, D. P. A new species of skink of the genus Dasia Gray 1889 (Reptilia: Scincidae) from Car Nicobar Islands, India. J. Bombay Nat. Hist. Soc. 74, 133–136 (1977).
Tiwari, K. K. & Biswas, S. Two new reptiles from the Great Nicobar Island. J. Zool. Soc. India 25, 57–63 (1973).
Biswas, S. & Sanyal, D. P. A report on the reptilia fauna of Andaman and Nicobars Islands in the collection of Zoological Survey of India. Rec. Zool. Surv. India 77, 255–292 (1980).
Heatwole, H. & Stuart, B. L. High Densities of a “ Rare “ Skink. Herpetol. Rev. 39, 169–170 (2008).
Steindachner, F. Reise der Osterreichischen Frigatte “Novara” um die Erde, in den Jahren 1857-58-59, unter den Befehlen des Commodore B von Wüllerstorf-Urbair. Zoologischer Theil. Erster Band. Reptilien., https://doi.org/10.1038/001602a0 (Kaiserlich-Königlischen, 1867).
MacArthur, R. H. & Wilson, E. O. The Theory of Island Biogeography. (Princeton University Press, 1967).
Veech, J. A. A probabilistic model for analysing species co-occurrence. Glob. Ecol. Biogeogr. 22, 252–260 (2013).
Veech, J. A. The pairwise approach to analysing species co-occurrence. J. Biogeogr. 41, 1029–1035 (2014).
Griffith, D. M., Veech, J. A. & Marsh, C. J. cooccur: Probabilistic Species Co-Occurrence Analysis in R. J. Stat. Softw. 69, 1–17, https://cran.r-project.org/web/packages/cooccur/ (2016).
R Core Team. R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. www.R-project.org/ (2017).
Prado, P. I. & Miranda, M. D. Package ‘ sads’. https://cran.r-project.org/web/packages/sads/ (2017).
Johnson, N. L., Kotz, S. & Balakrishnan, N. Continuous Univariate Distributions, volume 2. (Wiley, 1995).
Motomura, I. On the statistical treatment of communities. Zool. Mag. 44, 379–383 (1932).
Fisher, A. G., Corbet, S. A. & Williams, S. A. The relation between the number of species and the number of individuals in a random sample of an animal population. J. Anim. Ecol. 12, 42–58 (1943).
Weibull, W. A statistical distribution function of wide applicability. J. Appl. Mech. 18, 293–297 (1951).
Magurran, A. E. & McGill, B. J. Biological Diversity - Frontiers in measurement and assessment. (Oxford University Press, 2011).
Alonso, D. & McKane, A. J. Sampling Hubbell’s neutral model of biodiversity. Ecol. Lett. 7, 901–910 (2004).
Engen, S., Lande, R., Walla, T. & DeVries, P. J. Analyzing spatial structure of communities using the two-dimensional Poisson lognormal species abundance model. Am. Nat. 160, 60–73 (2002).
Volkov, I., Banavar, J. R., Hubbell, S. P. & Maritan, A. Neutral theory and relative species abundance in ecology. Nature 424, 1035–1037 (2003).
Pileou, E. Mathematical Ecology. (Wiley-Blackwell, 1977).
Engen, S. & Lande, R. Population dynamic models generating the lognormal species abundance distribution. Math. Biosci. 132, 169–183 (1996).
Schluter, D. A variance test for detecting species associations with some example applications. Ecology 65, 998–1005 (1984).
Gotelli, N. J., Hart, E. M. & Ellison, A. M. EcoSimR: Null model analysis for ecological data. R package version 0.1.0. https://CRAN.R-project.org/package=EcoSimR (2015).
Acknowledgements
This work was supported by Department of Science and Technology (DST), Government of India (Grant No. SR/SO/AS-08/2009). We thank Department of Forests and Wildlife, Andaman and Nicobar Islands, for providing necessary permits to conduct fieldwork in the Andaman and Nicobar Islands (File No. CWLW/WL/134/161). We are grateful to the Andaman and Nicobar Environmental Team (ANET) for logistical support. For help in data collection in the Andaman Islands we thank researchers Nitya Prakash Mohanty, S. R. Chandramouli, Chetan Rao and our field assistants Saw Glen, Sebe Horo, Anand James Tirkey, and Sudhir Kumar Ekka.
Author information
Authors and Affiliations
Contributions
This study was conceptualized by K.V. and S.H. Field data collection and analysis were done by S.H. The manuscript was co-written by S.H. and K.V.
Corresponding author
Ethics declarations
Competing Interests
The authors declare no competing interests.
Additional information
Publisher's note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Harikrishnan, S., Vasudevan, K. Niche dissociated assembly drives insular lizard community organization. Sci Rep 8, 11978 (2018). https://doi.org/10.1038/s41598-018-30427-4
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-018-30427-4
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
