
Abstract
Invasive species such as Bombus terrestris represent a severe threat that can result in the decline of local biodiversity. We examined the habitat suitability for B. terrestris invasion in East Asian countries and the risk of habitat overlap with 24 bumblebee species from different groups in China. All East Asian countries were predicted to be susceptible to invasion by B. terrestris, with the highest habitat suitability occurring in China followed by Japan, North Korea, South Korea and Mongolia. Within China, which is a global biodiversity hotspot for bumblebees, three different regions, north-west, central to south-east and north-east, were predicted as being highly suitable for invasion. In China, the group of species closely related to B. terrestris showed higher sensitivity (89%) to habitat overlap with B. terrestris than did the group of non-closely related species (73%). The risk of overlap for the three major regional bumblebees within China decreased in the order southern region, northern region and Tibetan Plateau region. Due to the sensitivity of habitat suitability and overlap, the use of the introduced European bumblebee B. terrestris for pollination services should be discouraged in regions with overlapping habitats in China, and management strategies should be implemented to conserve the vulnerable bumblebees in all East Asian countries.
Similar content being viewed by others
Introduction
Local biodiversity faces a serious threat from the introduction of alien invasive species, which can result in extinctions or population declines1,2,3,4,5. The trade of invasive species facilitates the establishment of non-native species in new areas6. In many cases, the intentional introduction of non-native species represents an important benefit to economies7. However, the trade of non-native species, e.g., Bombus terrestris, produces conflicts between the associated benefits and problems1,3,8,9,10.
In approximately 1900, a European bumblebee, Bombus terrestris (Hymenoptera: Apidae), was first introduced in New Zealand for pollination services. Currently, it is an invasive species and has spread well beyond its native range10,11,12,13. Since the 1980s, with the development of greenhouse agriculture, millions of colonies of bumblebees (B. terrestris) have been produced per year as managed pollinators to meet the demands of pollination services worldwide. Trade in bumblebee colonies has provoked concerns over the invasion of B. terrestris in regards to competitive displacement and the potential for horizontal disease transmission to local bumblebees5,14,15,16,17,18,19,20,21 via accidental escape of queens from greenhouses that establish colonies in areas where this bumblebee is non-native10. Recently, North America, Japan, Chile, and Argentina have faced challenges due to the establishment of B. terrestris and population declines of local bumblebee species3,16,22.
Among countries worldwide, China has the richest bumblebee diversity; 125 species are found in China, representing 50% of the total number of bumblebee species worldwide. Bumblebee pollinators play significant roles in agricultural and natural ecosystems23,24. Although B. terrestris is now part of the Chinese bumblebee biodiversity, it originated in Europe and has been expanding its range from Europe to Central Asia, including into northwest Xinjiang, the westernmost region of China bordering Kazakhstan25,26. Four decades ago, bumblebees collected in Xinjiang were considered B. lucorum, but recent DNA barcoding has confirmed that the B. lucorum complex in this region now includes B. terrestris27,28. The continued expansion of this species towards eastern China has been restricted by the natural barriers of the Takla Makan Desert, the Badain Jaran Desert and the high-altitude Tibetan Plateau between the western and eastern parts of China. In the past 10 years, commercial B. terrestris colonies from Europe have increasingly been introduced into China and some East Asian countries to meet the demands of crop pollination in greenhouses. This practice has raised the question of whether there are risks to the habitats of native bumblebees from this introduced species. They might pose a serious threat to native bumblebees considering the high likelihood that native bumblebees they will interact with B. terrestris around flowers29. Habitat or range contact (overlap) can enhance interspecific competition between species30, and B. terrestris introduction has already disturbed the natural mating of closely related local bumblebee species in Japan31. An analysis of habitat overlap risk is necessary to identify the areas that may have favourable conditions for interspecific competition between B. terrestris and vulnerable local bumblebee species. Such an analysis can be performed through spatial distribution modelling (SDM)32 and GIS (geographical information system) approaches. SDM techniques are considered good predictive tools and have been used previously to model the distribution of invasive or unknown species33,34. These approaches are among the most important for developing conservation strategies for declining bumblebee species19,35. These techniques combine climatic data and species occurrence records to identify the most suitable environmental conditions for population maintenance or possible species overlap. If the niches of different bumblebee species overlap, interspecific competition for food resources and habitats between invasive and local bumblebee species may occur. As overall biases and uncertainty are inherent to different prediction algorithm models, these models predict distributions differently36,37,38. Therefore, in the present study, five modelling approaches were used to identify the areas most suitable for B. terrestris; the results may guide the development of effective importation regulations for this species within East Asian countries, including China. This study attempts to answer the following questions: (1) Are all East Asian countries susceptible to B. terrestris invasion? (2) What regions of China offer potential habitats facilitating B. terrestris development and establishment? (3) Are some groups of local bumblebee species more sensitive than others to habitat overlap with B. terrestris within China?
Results
Modelling accuracy
The area-under-the-curve (AUC) values showed average to excellent model performances (0.77~0.98). However, variation in model performance from poor to excellent was observed based on the true skilled statistics (TSS) values (0~0.98). The ranges of TSS values were 0~0.86, 0.27~0.97, 0.35~0.95, 0.40~0.93 and 0~0.98 for Envelope Score, Environmental Distance, Genetic Algorithm for Rule Set Prediction (GARP), MaxEnt and Support Vector Machine (SVM), respectively (see Supplementary Table S1). The highest modelling accuracy was found with MaxEnt for almost all 25 species including B. terrestris, and the models with the next highest levels of accuracy were GARP and Environmental Distance, Envelope Score and SVM for 24, 19, 19, 7 and 1 species, respectively, under the receiver operating characteristics curves (ROC) threshold (see Supplementary Fig. S1). Different models (with TSS >0.5) were selected for each species to calculate the final “summed” distribution maps; however, for six species, B. ignitus, B. longipennis, B. lucorum, B. patagiatus, B. picipes and B. pyrosoma, only one modelling algorithm had a TSS >0.5 (see Supplementary Table S1).
Potential distributions of bumblebee species
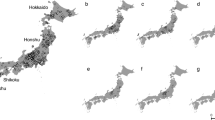
Predicted “suitable” and “highly suitable” areas for B. terrestris were detected in all East Asian countries, with the highest risk of invasion occurring in China, followed by Japan, North Korea, South Korea and Mongolia (Figs 1 & 2). However, within China, three different regions, north-west, central to south-east and north-east, were determined to be at risk; these areas cover an area of 2,001,333 km2, representing 21% of the total study area. B. terrestris expanded originally from Europe to the north-west part of China, so the central to south-east and north-east parts of China are new potential habitats for B. terrestris to become established and develop. These three zones cover 24 provinces considered to be under the threat of invasion from B. terrestris: Xinjiang, Gansu, Ningxia, Shaanxi, Shanxi, Hebei, Henan, Shandong, Chongqing, Hubei, Hunan, Guizhou, Guangxi, Guangdong, Fujian, Jiangxi, Anhui, Jiangsu, Shanghai, Zhejiang, Neimenggu, Liaoning, Jilin and Heilongjiang (Fig. 3).

Potential distribution of Bombus terrestris in East Asian countries. Red represents highly suitable habitat, yellow represents suitable habitat, and green represents unsuitable habitat for B. terrestris. The map was created with ArcGIS v 10.0 (www.arcgis.com).

Suitable habitat areas (km2) for the invasion of Bombus terrestris in all East Asian countries.

Map of the Chinese mainland showing the predicted distribution of Bombus terrestris with the names of the provinces in China. Red represents the highly suitable habitat, yellow represents suitable habitat, and green represents unsuitable habitat for B. terrestris. The map was created with ArcGIS v 10.0 (www.arcgis.com).
The modelling results revealed the potential distributions of all local bumblebee species across the different regions of China. The potential “suitable” and “highly suitable” areas for these 25 species ranged from 665,430 to 5,707,845 km2. The species with the largest distribution area, representing 59% of the total study area of China, was B. sibiricus, and B. longipennis had the smallest predicted distribution area, representing 7% of the total country area (Table 1). The optimum cut-off values for all 25 species ranged from 0.44~0.91, as shown in Table 139.
Six of seven native Tibetan bumblebee species, B. convexus, B. minshaenensis, B. rufofasciatus, B. supremus, B. ladakhensis and B. longipennis, had potential suitable and highly suitable habitats towards the Tibetan Plateau of China. However, only one species from this region, B. friseanus, had a potential distribution towards both Tibet and south-east China. All five southern species, B. bicoloratus, B. braccatus, B. breviceps, B. picipes and B. trifasciatus, had potential distributions towards the southern and south-eastern parts of the country. Unlike the other regions, the species from the northern regions had four different types of distribution trends. Two of these species, B. hypocrita and B. sporadicus, had potential distributions towards the north-east of China. Four species, B. lantschouensis, B. patagiatus, B. longipes and B. ussurensis, were present from central to north-east China. Two species, B. ignitus and B. pyrosoma, had distributions towards central to northern China. The remaining four species, B. cryptarum, B. lucorum, B. muscorum and B. sibiricus, had distributions towards north-east and north-west China (see Supplementary Fig. S2).
Habitat overlap risk analysis
There were 19 of 24 (79%) native bumblebee species overlapping with B. terrestris (Table 1). On average, the potential area of overlap between B. terrestris and native species was 464,900 ± 360,527 km2 (mean ± standard deviation). The sensitivity of habitat overlap was found to be greater (89%) for the “closely related” group (biological group) of the same subgenus, Bombus s. str., with B. terrestris compared to the “non-closely related” group (73%) of other subgenera of bumblebee species. For regional species, species from both the “southern” and “northern” regions had maximum overlap (100%) compared to the species from the “Tibetan Plateau” (29%). However, in the overlap areas, the species from the southern region had greater overlap with B. terrestris than did the species from the northern region, and only two species from the Tibetan Plateau had (slight) overlap with B. terrestris (Figs 4 and 5).

Habitat overlap risk analysis between Bombus terrestris and vulnerable native bumblebee species in China. Map was created with ArcGIS v 10.0 (www.arcgis.com).

Potential suitable areas vs overlapping habitat areas (%) between native bumblebee species and B. terrestris. The three shades of grey representing the “southern”, “northern” and “Tibetan Plateau” regions revealed that the species from the southern region have the most overlapping habitat areas with B. terrestris followed by the northern region and the Tibetan Plateau.
The spatial distributions of overlapping habitats between native and invasive species were found in all three regions with suitable habitat for B. terrestris: north-west, central to south-east and north-east within China. All five “southern” species, including B. bicoloratus, B. braccatus, B. breviceps, B. picipes and B. trifasciatus, mainly had potential overlapping habitats in central to south-east China. Six of the 12 northern species, B. cryptarum, B. hypocrita, B. lantschouensis, B. longipes, B. lucorum and B. muscorum, had potential overlapping habitats in all three regions with habitats suitable for B. terrestris. Five other “northern” species, B. ignitus, B. patagiatus, B. pyrosoma, B. sporadicus and B. ussurensis, had potential overlapping habitats towards central to north-east China. However, only one “northern” species, B. sibiricus, had a potential overlapping habitat towards central to north-west China. Similarly, two “Tibetan Plateau” species, B. minshaenensis and B. rufofasciatus, had potential overlapping habitats towards central to north-west China.
Eight of the nine species from the “closely related” group had overlapping habitats, and the range of overlapping habitat area was 114,425~776,801 km2 (1~9%). However, 11 of the 15 species from the “non-closely” related group had overlapping habitats, and the range of overlapping habitat area was 109,520~1,160,506 km2 (1~12%). Although all species in the “southern” and “northern” regional groups had overlapping habitats, the species in the “southern” group had a greater area of overlapping habitats, from 389,307~1,160,507 km2 (4~12%), than that, from 132,155 km2~776,801 km2 (1~8%), of the species in the “northern” group. However, the species from the “Tibetan Plateau” had an overlapping habitat area that ranged from 109,520~114,425 km2, which was only 1% of the whole area of China (Table 1, Fig. 5).
Discussion
In comparison to a single-model approach, summed distribution maps can provide more accurate estimations of the spatial distributions of species40. In the present study, five modelling approaches were implemented here to accurately determine the spatial distributions of B. terrestris throughout East Asian countries and of 24 local bumblebee species within China. To assess the accuracy of such models, TSS, AUC and kappa statistics values can be used. However, there are inherent limitations to using kappa statistics41, and AUC approaches do not allow the prediction of detection differences between more than two models42. The prevailing problem in kappa statistics involves their dependency on the ratio between the proportion of correctly predicted presences (sensitivity) and the proportion of correctly predicted absences (specificity)41. To avoid these problems, TSS values were calculated to assess model accuracy in the present study. The different ranges of TSS values among the different modelling approaches are due to the different mechanistic features of the models, and typically, these different features lead to model differences in predicted distributions37,43. These TSS ranges facilitated our evaluation of the modelling approaches used here. MaxEnt attained the highest performance among the modelling approaches, which is consistent with previous studies40,44,45,46,47.
In contrast to the findings of Acosta19, where East Asian countries including Japan, South Korea, North Korea and south-eastern coastal parts of China were considered suitable for B. terrestris invasion, our habitat suitability modelling approaches predicted that Mongolia as well as north-west, central to south-east and north-east parts of China (covering 24 provinces) are also suitable for invasion by B. terrestris. These regions are physiographically similar to Europe and have already been predicted to be climatically suitable for B. terrestris48,49. However, none of our models predicted that habitat susceptibility to invasion would occur only towards the south-eastern area. Our results are supported by the presence of some new records of B. terrestris in the predicted suitable habitats in China as well as in neighbouring countries surrounding China.
Interspecific mating between “closely related” invasive and native bumblebee species may occur, disturbing reproduction and reducing local bumblebee diversity. Likewise, in Japan, the invasive species B. terrestris has been found to mate with its closely related local species B. hypocrita and is responsible for the reduction in the local bumblebee population50. In addition, hybrid-mating disturbance has been found between B. terrestris males and native queens of Chinese B. lantschouensis under artificial conditions and may also occur in the field51. Although eight of the nine species from the “closely related” group (species of the same subgenus, Bombus s. str.) within China overlapped with B. terrestris, four species of the “non-closely related” group from the subtropical region of south China, B. trifasciatus, B. braccatus, B. breviceps and B. bicoloratus, had greater areas of overlapping habitat than did any of the “closely related” species” (Fig. 5). This habitat overlap between the invasive species and native species from the “non-closely related” group might be due to their ecological similarities30. More suitable similar habitats are responsible for the maximum overlap between B. terrestris and local bumblebees. More importantly, B. muscorum from the “non-closely related” group is already categorised as vulnerable in Britain52 and is vulnerable in China. Given that the foraging distance of B. muscorum (which forages mostly within 500 m of its nest) is shorter than that of B. terrestris (which forages mostly over 2 km from its nest)53, Habitat overlap with B. terrestris may be harmful for this local species because B. terrestris may be able to outcompete B. muscorum for food resources within areas close to the latter’s nests.
Alarmingly, eight species endemic to China23, B. braccatus, B. convexus, B. longipes, B. minshaenensis, B. picipes, B. pyrosoma, B. supremus and B. trifasciatus, are threatened by habitat overlap with the invasive B. terrestris. These species are important pollinators of many wild flowers and crops within China24, and endemic species are considered more important to local biodiversity than are non-native species54. Therefore, a management strategy should be implemented that prioritises the conservation of these endemic bumblebees.
Similarly, some of the overlapping species, specifically, B. braccatus, B. cryptarum, B. hypocrita, B. lucorum and B. muscorum, were “rare” in terms of abundance in this study (Table 1). Two of them, B. lucorum and B. muscorum, are also “moderately rare” in Hungry and Russia55,56,57,58. These observations highlight the potential for declines in vulnerable species within a country. However, due to species differences in food selection, the conservation strategies used for other species will not be effective for these rare species. Thus, from a conservation point of view, specific conservation planning is urgently needed to conserve these bumblebee species59,60.
The native bumblebee species from the “southern”, “northern” and “Tibetan Plateau” regions were distributed at different altitudes (Fig. 6). All the native species from the southern and northern regions at mean elevations <2500 metres had overlapping habitats. However, for the high, cold species from the Tibetan Plateau at mean elevations >2500, only two species, B. minshaenensis and B. rufofasciatus, had a very small proportion of overlapping habitats (<1% areas) with B. terrestris, and the remaining species were not overlapping (Fig. 6). This scenario might indicate that the high, cold region of the Tibetan Plateau has the least amount of suitable habitat for the invasive species or that these species have ecological dissimilarities30.

The proportion of contribution of the potential suitable habitat areas (%) of both B. terrestris and native bumblebee species that overlap at different mean elevations.
Our current findings regarding the distribution of B. terrestris and its overlapping habitats with different groups of local bumblebees showed that the local bumblebees of other East Asian countries might also be threatened by habitat overlap and loss of native biodiversity. Proper management strategies and coordinated implementation of regulations related to the introduction of B. terrestris61 should be considered for all East Asian countries to conserve the natural biodiversity of local bumblebee species within this region.
Methods
Modelled species
Records of B. terrestris were based on its natural distribution within China and surrounding countries, such as Kazakhstan, Kyrgyzstan and Russia, as well as within the areas close to China such as Japan and South Korea, where it is invasive (www.discoverlife.org) (Fig. 7). For the local bumblebees in China, we selected 24 native bumblebee species and B. terrestris with a total of 26,497 specimen records from the IAR (CAAS Institute of Apiculture Research) collection that covers the whole region of the Chinese mainland (Table 1). These species were represented in >50% of the records of the 125 bumblebee species in China and were selected by two principles, 1) representatives of two groups: species “closely related” to B. terrestris, i.e., from the subgenus Bombus s. str., all known in China (n = 10) and “non-closely related” species, i.e., species of other bumblebee subgenera (n = 14), and 2) representatives of the three major regions of China, i.e., the high cold “Tibetan Plateau” and the temperate “northern” and subtropical “southern” regions (Table 1).

Relief map of the Chinese mainland showing the sample sites of the 25 bumblebee species with the different colourful spots representing the different species in this study. The records of B. terrestris for areas surrounding China are from (www.discoverlife.org). The international boundaries are shown in grey with the names of countries in capital letters. The map was created with ArcGIS v 10.0 (www.arcgis.com).
Environmental layers and modelling procedures
In our modelling, we used 19 bioclimatic and elevation layers obtained from www.worldclim.org with a grid cell size of 5 arc-min resolution (≈10 km2) of the WGS1984 projection62 with full coverage of East Asian countries. We converted all these layers into ASCII format to use in our modelling software with ArcGIS v 10.0. We calculated Pearson’s correlation coefficients with the species distribution modelling toolbox63 of ArcGIS v 10.0 to reduce the collinearity among these bioclimatic layers, and we retained only seven bioclimatic variables together with the elevation layers, which all yielded Pearson correlation coefficient (r) values < 0.8, to avoid overfitting the model (see Supplementary Table S2). Similarly, we used a spatial rarefy tool of the SDM toolbox63 to remove all the presence points of species that have distances <10 km2 to avoid spatial autocorrelation among these occurrence records of bumblebee species during modelling (Table 1). All the occurrence records were converted into comma separated value (CSV) comma delimited and text tab delimited formats for the use in MaxEnt v 3.3.3k and OpenModeller Desktop software v 1.1.0, respectively. For MaxEnt software, we prepared a biased file of all 25 species occurrence records for background point selection using ArcGIS v 10.0.
Five modelling algorithms, MaxEnt64, Envelope Score65, Environmental Distance66, GARP67 and SVM68, were implemented to increase the reliability of the potential distributions as well as to identify areas of overlap between the 24 native bumblebee species and B. terrestris40. These models were selected mainly for their different mechanistic features69. Due to the variation in the distribution data of a species, there is error in the estimation of the spatial distribution of species by different modelling approaches. As a result, similar data used in different modelling approaches can yield very different results70. Therefore, testing more than one model is preferable for addressing the error38. All modelling approaches have inherent differences in the generation of distribution predictions37,43. By comparing their results, one can retain only those modelling approaches that yielded similar and accurate species distributions37. Among the algorithms considered in our study, the envelope score and environmental distance algorithms are simpler algorithms that predict species distributions only on the basis of presence data. In contrast, the remaining three modelling approaches, MaxEnt, SVM and GARP, are more complex and involve artificial intelligence methods. These approaches have been shown to reliably predict species occurrence43. Two software programs, MaxEnt v 3.3.3k and OpenModeller Desktop v 1.1.0, were used to model the distributions of bumblebee species71. We developed our models using 75% of our data for training and the remaining 25% for testing our models.
In contrast to the Pearson72 approach, where the lowest presence threshold (LPT) is used, in this study, only receiver operating characteristics curve (ROC) threshold values were used to cut the modelled suitability matrices. This approach determines the balance between omission and commission errors while determining the spatial distributional ranges of a species40. Furthermore, TSS values were used (which range from −1 to 1) to evaluate the accuracies of the five modelling approaches26,41. Here, values close to zero or negative mean that the distribution is not much better than random, whereas values close to one indicate almost perfect agreement between the modelling prediction and distribution. Normally, TSS values ≥0.5 are considered acceptable40. Finally, we selected only the distribution modelling outputs that yielded TSS values ≥0.5 (see Supplementary Table S1) to develop the “summed” distribution maps within ArcGIS v 10.0 that produced relatively realistic and potentially accurate spatial distributions of the species. In this study, 10,000 random pseudo-absences were used. We categorised our final “summed” potential distribution maps into three categories, 1 = unsuitable habitats, 2 = suitable habitats and 3 = high suitable habitats, based on their specific optimum threshold cut off values within ArcGIS v 10.0. (Table 1).
Assessment of overlapping habitats
To assess the habitat overlap of both invasive and local bumblebee species, the Youden Index was applied, calculated with following the formula: sensitivity + specificity −139,73. This index is used to determine the cut-off point for the models. A high Youden Index value indicates good performance of the model39. The final “summed” potential distribution maps were divided into suitable (>threshold value of Youden Index) and unsuitable (<threshold value of Youden Index) categories within ArcGIS v 10.0. The threshold value for each species is calculated as the average TSS values of all those modelling algorithms with TSS values >0.5 (see Supplementary Table S1). Then, the distribution maps of the 24 local species were overlaid with the final “summed” distribution map of B. terrestris, and the cells were reclassified into three categories using the Arc toolbox of ArcGIS v 10.0: 1 = unsuitable habitat for both species (local and invasive species), 2 = suitable habitat for one species but unsuitable for the other species, and 3 = suitable habitat for both species (Fig. 4). The optimum cut off values for all the 25 bumblebee species are shown in Table 1. All the grid cells that have values below this average threshold are considered unsuitable for species presence or vice versa.
References
Matsumura, C., Yokoyama, J. & Washitani, I. Invasion status and potential ecological impacts of an invasive alien bumblebee, Bombus terrestris L. (Hymenoptera: Apidae) naturalized in Southern Hokkaido, Japan. Glob. Environ. Res. 8, 51–66 (2004).
Clavero, M. & García-Berthou, E. Invasive species are a leading cause of animal extinctions. Trends Ecol. Evol. 20, 110 (2005).
Inari, N., Nagamitsu, T., Kenta, T., Goka, K. & Hiura, T. Spatial and temporal pattern of introduced Bombus terrestris abundance in Hokkaido, Japan, and its potential impact on native bumblebees. Popul. Ecol. 47, 77–82 (2005).
Mooney, H. A. & Cleland, E. E. The evolutionary impact of invasive species. Proc. Nat. Acad. Sci. 98, 5446–5451 (2001).
Morales, C. L., Arbetman, M. P., Cameron, S. A. & Aizen, M. A. Rapid ecological replacement of a native bumble bee by invasive species. Front. Ecol. Environ. 11, 529–534 (2013).
Perrings, C., Burgiel, S., Lonsdale, M., Mooney, H. & Williamson, M. International cooperation in the solution to trade-related invasive species risks. Ann. N. Y. Acad. Sci. 1195, 198–212 (2010).
Sandlund, O. T., Schei, P. J. & Viken, A. Introduction: the many aspects of the invasive alien species problem. Invasive species Biodivers. Manag. Kluwer Acad. Publ. Dordrecht, Netherlands 1–7 (1999).
Inoue, M. N., Yokoyama, J. & Washitani, I. Displacement of Japanese native bumblebees by the recently introduced Bombus terrestris (L.) (Hymenoptera: Apidae). J. Insect Conserv. 12, 135–146 (2008).
Molnar, J. L., Gamboa, R. L., Revenga, C. & Spalding, M. D. Assessing the global threat of invasive species to marine biodiversity. Front. Ecol. Environ. 6, 485–492 (2008).
Velthuis, H. H. W. & Doorn, A. Van. A century of advances in bumblebee domestication and the economic and environmental aspects of its commercialization for pollination. Apidologie 37, 421–451 (2006).
Goka, K., Okabe, K., Yoneda, M. & Niwas, S. Bumblebee commercialization will cause worldwide migration of parasitic mites. Mol. Ecol. 10, 2095–2099 (2001).
Byers, J. E. et al. Directing research to reduce the impacts of non-indigenous species. Conserv Biol. 16, 630–40 (2002).
Williams, P. H. Environmental change and the distributions of British bumble bees (Bombus Latr.). Bee world 67, 50–61 (1986).
Schmid-Hempel, P. & Schmid-Hempel, R. Transmission of a pathogen in Bombus terrestris, with a note on division of labor in social insects. Behav. Ecol. Sociobiol. 33, 319–327 (1993).
Ings, T. C., Ward, N. L. L. & Chittka, L. Can commercially imported bumble bees out-compete their native conspecifics? J. Appl. Ecol. 43, 940–948 (2006).
Schmid-Hempel, P., Schmid-Hempel, R., Brunner, P. C., Seeman, O. D. & Allen, G. R. Invasion success of the bumblebee, Bombus terrestris, despite a drastic genetic bottleneck. Heredity (Edinb). 99, 414–422 (2007).
Schmid-Hempel, R. et al. The invasion of southern South America by imported bumblebees and associated parasites. J. Anim. Ecol. 83, 823–837 (2014).
Arbetman, M. P., Meeus, I., Morales, C. L., Aizen, M. A. & Smagghe, G. Alien parasite hitchhikes to Patagonia on invasive bumblebee. Biol. Invasions 15, 489–494 (2013).
Acosta, A. L., Giannini, T. C., Imperatriz-Fonseca, V. L. & Saraiva, A. M. Worldwide alien invasion: A methodological approach to forecast the potential spread of a highly invasive pollinator. PLoS ONE 11, 1–25 (2016).
Polidori, C. & Nieves-Aldrey, J. L. Comparative flight morphology in queens of invasive and native Patagonian bumblebees (Hymenoptera: Bombus). C. R. Biol. 338, 126–133 (2015).
Krosby, M. et al. Climate-induced range overlap among closely related species. Nat. Clim. Chang. 5, 883–886 (2015).
Goulson, D. Conservation of Bumblebees. Species Manag. Challenges Solut. 21st Century 25, 1–16 (2010).
Huang, J. & An, J. Species diversity, pollination application and strategy for conservation of the bumblebees of China. Biodivers. Sci. 26, 486–497 (2018).
An, J. et al. The bumblebees of North China (Apidae, Bombus Latreille). Zootaxa 3830, 1–89 (2014).
Williams, P. H. et al. Cryptic bumblebee species: Consequences for conservation and the trade in greenhouse pollinators. PLoS ONE 7, e32992 (2012).
Liu, C., White, M. & Newell, G. Measuring and comparing the accuracy of species distribution models with presence–absence data. Ecography 34, 232–243 (2011).
Wang, S. & Yao, J. Apidae-Bombus. In: Organisms of the Tuomuerfeng region of Tianshan, Sinkiang Renmin publishing house, Urumchi, China. 160–165 (1985).
Liu, P., Huang, J., An, J., He, S. & Wu, J. Molecular identification and distribution characteristics of the bumblebee Bombus lucorum complex in China. Acta Entomol. Sin. 57, 235–243 (2014).
Goulson, D., Lye, G. C. & Darvill, B. Decline and conservation of bumble bees. Annu. Rev. Entomol. 53, 191–210 (2008).
Jankowski, J. E., Robinson, S. K. & Levey, D. J. Squeezed at the top: Interspecific aggression may constrain elevational ranges in tropical birds. Ecology 91, 1877–1884 (2010).
Kanbe, Y., Okada, I., Yoneda, M., Goka, K. & Tsuchida, K. Interspecific mating of the introduced bumblebee Bombus terrestris and the native Japanese bumblebee Bombus hypocrita sapporoensis results in inviable hybrids. Naturwissenschaften 95, 1003–1008 (2008).
Guisan, A. & Zimmermann, N. E. Predictive habitat distribution models in ecology. Ecol. Modell. 135, 147–186 (2000).
Araújo, M. B. & Peterson, A. T. Uses and misuses of bioclimatic envelope modeling. Ecology 93, 1527–1539 (2012).
Jiménez-Valverde, A. et al. Use of niche models in invasive species risk assessments. Biol. Invasions 13, 2785–2797 (2011).
Eken, G. et al. Key Biodiversity areas as site conservation targets. Bioscince 54, 1110–1118 (2004).
Barry, S. & Elith, J. Error and uncertainty in habitat models. J. Appl. Ecol. 43, 413–423 (2006).
Diniz-Filho, J. A. F. et al. Partitioning and mapping uncertainties in ensembles of forecasts of species turnover under climate change. Ecography 32, 897–906 (2009).
Rocchini, D. et al. Accounting for uncertainty when mapping species distributions: the need for maps of ignorance. Prog. Phys. Geogr. 35, 211–226 (2011).
Wu, W., Li, Y. & Hu, Y. Simulation of potential habitat overlap between red deer (Cervus elaphus) and roe deer (Capreolus capreolus) in northeastern China. Peer J. 4, e1756 (2016).
Rangel, T. F. et al. Seeking the flowers for the bees: Integrating biotic interactions into niche models to assess the distribution of the exotic bee species Lithurgus huberi in South America. Ecol. Modell. 273, 200–209 (2014).
Allouche, O., Tsoar, A. & Kadmon, R. Assessing the accuracy of species distribution models: prevalence, kappa and the true skill statistic (TSS). J. Appl. Ecol. 43, 1223–1232 (2006).
Ray, D., Behera, M. D. & Jacob, J. Evaluating ecological niche models: A comparison between Maxent and GARP for predicting distribution of Hevea brasiliensis in India. Proc. Natl. Acad. Sci. India Sect. B Biol. Sci. https://doi.org/10.1007/s40011-017-0869-5 (2017).
Rangel, T. F. & Loyola, R. D. Labeling ecological niche models. Nat. Conserv. 10, 119–126 (2012).
Wang, W., Lo, N., Chang, W. & Huang, K. Modeling spatial distribution of a rare and endangered plant species (Brainea insignis) in central Taiwan. ISPRS Arch. 241–246 (2012).
Padalia, H., Srivastava, V. & Kushwaha, S. P. S. Modeling potential invasion range of alien invasive species, Hyptis suaveolens (L.) Poit. in India: Comparison of MaxEnt and GARP. Ecol. Inform. 22, 36–43 (2014).
Alahmed, A. M., Naeem, M., Kheir, S. M. & Sallam, M. F. Ecological distribution modeling of two malaria mosquito vectors using geographical information system in Al-Baha Province, Kingdom of Saudi Arabia. Pak. J. Zool. 47, 1797–1806 (2015).
Naeem, M., Alahmed, A. M., Kheir, S. M. & Sallam, M. F. Spatial distribution modeling of Stegomyia aegypti and Culex tritaeniorhynchus (Diptera: Culicidae) in Al-Bahah Province, Kingdom of Saudi Arabia. Trop. Biomed. 33, 295–310 (2016).
Williams, P. H. An Annotated checklist of bumble bees with an analysis of patterns of description (Hymenoptera: Apidae, Bombini). Bull. Nat. Hist. Museum London 67, 79–152 (1998).
Lecocq, T., Rasmont, P., Harpke, A. & Schweiger, O. Improving international trade regulation by considering intraspecific variation for invasion risk assessment of commercially traded species: The Bombus terrestris case. Conserv. Lett. 9, 281–289 (2016).
Kondo, N. I. et al. Reproductive disturbance of Japanese bumblebees by the introduced European bumblebee Bombus terrestris. Naturwissenschaften 96, 467–475 (2009).
Yuan, X. et al. Evaluation of reproductive disturbance of Chinese bumblebees by European bumblebee Bombus terrestris (Hymenoptera: Apidae). Acta Entomol. Sin. 61, 348–359 (2018).
Nieto, A. et al. European red list of bees. in Publication office of the European Union (2014).
Walther-Hellwig, K. & Frankl, R. Foraging distances of Bombus muscorum, Bombus lapidarius, and Bombus terrestris (Hymenoptera, Apidae). J. Insect Behav. 13, 239–246 (2000).
Chitale, V. S., Behera, M. D. & Roy, P. S. Future of endemic flora of biodiversity hotspots in India. PLoS ONE 9, 1–15 (2014).
Walther-Hellwig, K. & Frankl, R. Foraging habitats and foraging distances of bumblebees, Bombus spp. (Hym., Apidae), in an agricultural landscape. J. Appl. Entomol. 124, 299–306 (2000).
Sárospataki, M., Novák, J. & Molnár, V. Assessing the threatened status of bumble bee species (Hymenoptera: Apidae) in Hungary, Central Europe. Biodivers. Conserv. 14, 2437–2446 (2005).
Darvill, B., Ellis, J. S., Lye, G. C. & Goulson, D. Population structure and inbreeding in a rare and declining bumblebee, Bombus muscorum (Hymenoptera: Apidae). Mol. Ecol. 15, 601–611 (2006).
Goulson, D., Hanley, M. E., Darvill, B. & Ellis, J. S. Biotope associations and the decline of bumblebees (Bombus spp.). J. Insect Conserv. 10, 95–103 (2006).
Goulson, D. et al. Causes of rarity in bumblebees. Biol. Conserv. 122, 1–8 (2005).
Dicks, L.V. et al. Ten policies for pollinators. Science 354, 975–976 (2016).
Aizen, M. A. et al. Coordinated species importation policies are needed to reduce serious invasions globally: The case of alien bumblebees in South America. J. Appl. Ecol. https://doi.org/10.1111/1365-2664.13121 (2018).
Hijmans, R. J., Cameron, S. E., Parra, J. L., Jones, P. G. & Jarvis, A. Very high resolution interpolated climate surfaces for global land areas. Int. J. Climatol. 25, 1965–1978 (2005).
Brown, B. J. L., City, T. & York, N. SDMtoolbox User Guide (2014).
Phillips, S. J., Anderson, R. P. & Schapire, R. E. Maximum entropy modeling of species geographic distributions. Ecol. Modell. 190, 231–259 (2006).
Pineiro, R., Aguilar, J. F., Munt, D. D. & Feliner, G. N. Ecology matters: Atlantic–Mediterranean disjunction in the sand-dune shrub Armeria pungens (Plumbaginaceae). Mol. Ecol. 16, 2155–2171 (2007).
Farber, O. & Kadmon, R. Assessment of alternative approaches for bioclimatic modeling with special emphasis on the Mahalanobis distance. Ecol. Modell. 160, 115–130 (2003).
Stockwell, D. & Peters, D. The GARP modelling system: problems and solutions to automated spatial prediction. Int. J. Geogr. Inf. Sci. 13, 143–158 (1999).
Tax, D. M. J. & Duin, R. P. W. Support vector data description. Mach. Learn. 54, 45–66 (2004).
Silva, D. P., Macêdo, A. C. B. A., Ascher, J. S. & De Marco, P. Range increase of a Neotropical orchid bee under future scenarios of climate change. J. Insect Conserv. 19, 901–910 (2015).
Araújo, M. B., Cabeza, M., Thuiller, W., Hannah, L. & Williams, P. H. Would climate change drive species out of reserves? An assessment of existing reserve-selection methods. Glob. Chang. Biol. 10, 1618–1626 (2004).
de Souza Muñoz, M. E. et al. OpenModeller: A generic approach to species’ potential distribution modelling. Geoinformatica:https://doi.org/10.1007/s10707-009-0090-7 (2009).
Pearson, R. G., Raxworthy, C. J., Nakamura, M. & Peterson, A. T. Predicting species distributions from small numbers of occurrence records: a test case using cryptic geckos in Madagascar. J. Biogeogr. 34, 102–117 (2007).
Garcia, C. V., Woodard, P. M., Titus, S. J., Adamowicz, W. L. & Lee, B. S. A Logit Model for predicting the daily occurrence of human caused forest fires. Int. J. Wildl. Fire 5, 101–111 (1995).
Acknowledgements
We are very thankful to Dr. Paul H. Williams for his help in identifying the bumblebees and to Dr. Jianghong Li for comments on the manuscript. This work was supported by the Agricultural Science and Technology Innovation Program (CAAS-ASTIP-2015-IAR) and the Natural Science Foundation of China (31672500, U1603108).
Author information
Authors and Affiliations
Contributions
J.A. and J.H. conceived and designed this research; M.N., X.Y., J.H. and J.A. reviewed the studied specimens and the data of the IAR bumblebee collection. M.N., J.H. and J.A. analysed the data and wrote the paper. All authors read and approved the final manuscript.
Corresponding authors
Ethics declarations
Competing Interests
The authors declare no competing interests.
Additional information
Publisher's note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Naeem, M., Yuan, X., Huang, J. et al. Habitat suitability for the invasion of Bombus terrestris in East Asian countries: A case study of spatial overlap with local Chinese bumblebees. Sci Rep 8, 11035 (2018). https://doi.org/10.1038/s41598-018-29414-6
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-018-29414-6
This article is cited by
-
Impact of climate change on potential distribution of Dickeya zeae causal agent of stalk rot of maize in Sialkot district Pakistan
Scientific Reports (2024)
-
Macroecological perspectives on the competition between the native and invasive bumblebees in southern South America under climate change
Biological Invasions (2024)
-
Ecosystem health and malfunctions: an organisational perspective
Biology & Philosophy (2023)
-
The cuckoo bumble bee, Bombus chinensis, has a fragmented habitat, as revealed using the maximum entropy approach (Hymenoptera: Apidae)
Apidologie (2022)
-
Colour patterns, distribution and food plants of the Asian bumblebee Bombus bicoloratus (Hymenoptera: Apidae)
Apidologie (2019)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
