
Abstract
Formaldehyde-based feed additives are approved in the US for Salmonella control and reducing bacterial contamination in animal feed. However, we hypothesize formaldehyde inclusion in swine diets may influence gut microbial composition due to its antimicrobial properties which might negatively influence microbial populations and pig growth performance. Also, formaldehyde inclusion in diets is known to reduce the dietary availability of amino acids. Therefore, our study was conducted to characterize if the effects of feed formaldehyde-treatment are due to influences on microbial population or diet amino acid (AA) sources. Dietary treatments were arranged in a (2 × 2) + 1 factorial with formaldehyde treatment (none vs. 1000 ppm formaldehyde) and crystalline AA inclusion (low vs. high) with deficient AA content plus a positive control diet to contain adequate AA content without dietary formaldehyde. Treating diets with formaldehyde reduced growth rate (P = 0.001) while the AA inclusion had no evidence of impact. Formaldehyde reduced feed bacterial content and altered fecal microbial communities (P < 0.05). Therefore, we conclude that the negative influence on growth was due to the impact on the fecal microbial community. Implications are that strategies for feed pathogen control need to take into account potential negative impacts on the gut microbial community.
Similar content being viewed by others


Introduction
In the swine industry, in-feed antibiotics have been used to control enteric infections and improve growth performance1. The use of growth promoting antibiotics has led to increased antibiotic resistance in gut bacteria2. In swine production systems there are considerable interest and effort in identifying feeding and management practices that maintain and improve production efficiency without the use of in-feed antibiotics. Formaldehyde-based feed additives can be used to reduce and or prevent bacterial contamination in animal feed3. According to the Food and Drug Administration’s federal register4 (#21 CFR 573.460), the food additive formaldehyde can be included in animal feed or ingredients to maintain complete feed and ingredients as Salmonella negative for up to 21 days. Formaldehyde-based products are widely used in the poultry industry for Salmonella control in feed5. Since the emergence of porcine epidemic diarrhea virus (PEDV) in the United States and demonstration that feed can be a vector for the transmission of the disease6 formaldehyde-based products have received attention as a potential method to reduce the risk of PEDV transmission in feed. Research demonstrating formaldehyde use in reducing PEDV infectivity in contaminated feed and ingredients has been successful7,8 and has led to an increased use of formaldehyde in pig feed. However, formaldehyde is known to produce reactions with numerous groups of amino acid residues of proteins that can lead to the formation of methylol groups, Schiff-bases, and methylene bridges amongst these residues9. Thus, inclusion in diets may reduce the small intestinal availability of dietary amino acids (AA) for pigs, which may negatively influence growth performance and nutrient utilization10,11,12. Likewise, information is lacking regarding formaldehyde inclusion in the diet of pigs and changes in gut microbiota which contributes to nutrient utilization. Therefore, a study was conducted in nursery pigs to determine if formaldehyde treatment would interact with dietary Lysine concentration and crystalline AA concentration and affect nursery pig growth performance, feed bacteria concentration, and fecal microbiota.
Results
Feed bacterial concentration
For diet bacterial concentrations, there was no evidence of difference (P > 0.10) for a crystalline AA × Formaldehyde interaction for any bacterial plate counts evaluated (Table 1). There was also no evidence of difference (P > 0.10) in bacterial plate counts evaluated between the low and high crystalline amino acid diets. Analysis of feed samples revealed that the control diet had greater (P = 0.027) aerobic plate counts compared to the other treatment diets and marginally significant greater (P = 0.062) Enterobacteriaceae counts. Formaldehyde inclusion in diets reduced (P < 0.05) Enterobacteriaceae and total coliform counts compared to diets that did not include formaldehyde.
Lysine analysis of the feed
As expected, the positive control diet contained a greater (1.51 ± 0.02; P = 0.001) amount of total Lys than the low or high crystalline amino acid diets with or without formaldehyde confirming that diets were formulated correctly (Table 1). Free lysine analysis confirmed that the crystalline AA were added correctly to the diets. A crystalline AA × Formaldehyde interaction was observed (P < 0.05) in analyzed diets for total Lys and total Lys percent difference from calculated values. This interaction occurred because formaldehyde inclusion in low crystalline AA diets reduced total Lys and had greater percent difference from calculated total Lys values than in the high crystalline AA diets. There was no evidence of difference between total Lys and percent difference from calculated total Lys values with or without the inclusion of formaldehyde in high crystalline AA diets.
Growth performance of piglets
Overall (d 0 to 28), pigs fed the control diet had improved (P < 0.05) average daily gain (ADG), ending body weight (BW), and gain to feed ratio (G:F) compared to those fed other diets containing reduced Lys (Table 2). This confirmed that Lys was limiting in the diets for pigs fed below their Lys requirement. The application of formaldehyde to diets reduced (P < 0.05) ADG (513 ± 7.30 vs 519 ± 7.30 g/d) and ending BW (26.8 ± 0.280, 26.7 ± 0.280 kg) compared to not treating diets with formaldehyde. A Crystalline AA × formaldehyde interaction (P < 0.05) was observed for average daily feed intake (ADFI) and G:F. The interaction for ADFI occurred because treating diets with formaldehyde decreased ADFI for pigs fed diets with high crystalline AA inclusions, but did not influence ADFI for pigs fed low crystalline AA diets. The interaction for G:F was observed because treating diets with formaldehyde decreased G:F for pigs fed low crystalline AA diets but did not influence G:F for pigs fed high crystalline AA diets.
Bacterial community structure
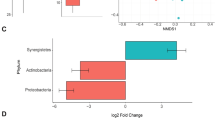
At the phylum level, the most dominant phyla were Firmicutes (74.8%) and Bacteroidetes (15.6%); however, there were no effects of dietary treatment at the phylum level. At the family level, 49 families were identified, among which 12 families having greater than 1% of core sequences. There was no evidence of differences (P > 0.10) in bacterial abundances amongst the dietary treatments for Methanobacteriaceae, Prevotellaceae, Lachnospiraceae, or Spirochaetaceae. A crystalline AA × formaldehyde interaction (P = 0.003) was observed for Streptococcaceae abundances in the bacterial community of the gut, because pigs fed low crystalline AA diets had a more dramatic reduction in abundance when treated with formaldehyde compared to the high crystalline AA diets (Fig. 1). The treatment of diets with formaldehyde decreased (0.69 ± 0.17, 0.48 ± 0.17; P < 0.05) bacterial abundance for Paraprevotellaceae and Lactobacillaceae species, while formaldehyde treatment increased (27.5 ± 2.65, 35.5 ± 2.65; P < 0.05) Clostridiaceae and Erysipelotrichaceae species within the bacterial community of the gut. Pigs fed formaldehyde-treated diets had marginal significance (3.75 ± 0.53, 2.94 ± 0.53; P = 0.074) for lower percentages of S24-7 bacteria species than pigs fed non-formaldehyde treated diets. Pigs fed low crystalline AA diets had increased (P < 0.05) abundance of Paraprevotellaceae, Lactobacilliaceae, Ruminococcaceae, and Veillonellaceae bacterial species compared to high crystalline AA diets. Pigs fed high crystalline AA diets had increased (25.9 ± 2.65, 35.5 ± 2.65; P = 0.007) Clostridiaceae and tended (2.51 ± 0.33, 3.19 ± 0.33; P = 0.080) to have increased Erysipelotrichaceae bacterial species compared to pigs fed low crystalline AA diets. Treatment diets fed to lower lysine levels than the control had increased (P = 0.009) Clostridiaceae bacterial species, while Paraprevotellaceae species were marginally (P = 0.091) lower in these diets compared to the positive control.

Bacterial abundance analysis among treatments groups evaluated at the family level. The graph exhibits the percent of each bacterial species that was detected.
Discussion
Our results are in agreement with earlier research using formaldehyde to reduce bacterial load in complete feeds for poultry13 and swine14,15. The addition of formaldehyde to poultry diets reduces contamination from specific bacterial pathogens such as Salmonella13. This is consistent with our results that indicated formaldehyde reduced Enterobacteriaceae and total coliform count in feed.
The use of formaldehyde and the subsequent effect on animal performance has been more extensively investigated in poultry compared to swine. For example, one study evaluated the addition of a commercially available formaldehyde additive to broiler diets and it improved feed conversion ratio by 4.8%16. However in another study, chickens received 1 of 4 diets treated with either 0, 630, 1580 or 6,300 mg formaldehyde/kg feed17. A reduction in body weight was observed for chickens fed formaldehyde treated diets, regardless of level, compared to the untreated feed. In swine, more limited research has been conducted evaluating the effects of formaldehyde on single ingredients or the complete diet on bacterial concentrations and performance. One study evaluated the application of formaldehyde to spray-dried animal plasma prior to complete diet manufacturing, which reduced the bacterial concentration14. Pigs fed diets with 6% formaldehyde-treated animal plasma had improved ADG and ADFI compared to pigs fed control diets with untreated animal plasma. In a follow up study these researchers observed that formaldehyde application to the complete diet reduced bacterial concentration of the total diet compared to those not treated with formaldehyde, but growth performance was not improved14. This study is consistent with our study suggesting that treating the complete diet reduces growth performance. We believe that formaldehyde treatment of an ingredient that was a relatively low proportion of the diet (6%) did not affect the microbial diversity of the feed, however treating the complete diet did affect the microbial content of the feed and led to reduced growth performance.
Formaldehyde has the ability to produce reactions with numerous groups of AA, including Lys9,18,19. Thus, we hypothesized the form of AA supplementation (crystalline AA or intact AA in protein) maybe affected by formaldehyde may be affected differently as reported previously20. The reactions between formaldehyde and AA, especially Lys, could render these AA unavailable which could possibly alter growth performance. Our results suggest the effect was greater in the low crystalline AA diets which contained more intact protein as a source of amino acids. The reactions that occur between formaldehyde and Lys residues in proteins may explain why formaldehyde is reducing the amount of total and available Lys in the current studies and could alter protein utilization, thus explaining the reduction in performance. The growth performance data suggests that formaldehyde is affecting the growth performance of the pigs both through the nutritive content of the feed and by affecting the microbial community within the feed.
The gastrointestinal tract of pigs harbors beneficial commensal bacteria that play a role in regulating gene expression that can influence gut health and immune responses21. Advancements in sequencing techniques have allowed researchers to focus on sequencing specific regions within the 16S rRNA gene to reveal patterns in microbial composition of the pig gastrointestinal tract22. Holman et al.23 utilized a meta-analysis to determine the major types of commensal bacteria species that inhabit the gastrointestinal tract of pigs. The researchers observed that Prevotella, Clostridium, Ruminococcus, and Lactobacillus species were found in greater than 90% of fecal samples collected in those studies, which would be similar to the findings of the current study. However, little data has been generated in pigs documenting the effects dietary manipulations have on gut microbial diversity. One study had demonstrated dietary effects on weaned pig fecal microbial diversity24 and other researchers have evaluated macro minerals and in-feed antimicrobials and their effects on gut microbiota changes25,26. When looking at the relationship between gut microbiota and nutrient utilization, there is an established possible link in piglets where feed efficiency is directly related to gut microbial compositional differences with a higher relative abundance of beneficial bacteria such as Clostridiales and Bacteroidetes10. However, our study is the first to observe the effects of formaldehyde treatment of diets on nursery gut microbiota. More uniquely we have demonstrated a decrease in lactic acid bacteria species, specifically Lactobacillaceae, and the increase of Clostridiaceae species in fecal samples obtained from pigs fed formaldehyde-treated diets. This warrants further investigation to determine the short- and long-term effects this shift in commensal bacterial species has on gut health. It has been well documented that antimicrobial therapy plays a pivotal role in the pathogenesis of Clostridia infection in humans, particularly Clostridium difficile infection. Antimicrobials known to disrupt the indigenous microbiota of the hindgut (colon), allowing Clostridia to grow in high concentrations27. Thus, in our study we shifted the microbial population by reducing more desirable species and increasing species with potential negative side effects.
In summary, as expected the inclusion of formaldehyde reduced bacterial concentration in nursery pig feed. However, these studies have provided evidence that in nursery pigs the inclusion of formaldehyde in complete feeds has a negative impact on growth performance. In addition, the inclusion of formaldehyde altered fecal microbial populations with a reduction in Lactobacillaceae species but increase in Clostridiaceae species. Thus, implications of this research are that the effect of pathogen control needs to be balanced against potential negative influences on microbial populations.
Materials and Methods
Animals and study design
The Kansas State University Institutional Animal Care and Use Committee (IACUC # 3529) approved the protocol used in this experiment and all experiments were performed in accordance with the relevant guidelines and regulations. A total of 1,235 pigs (PIC 359× Gentiporc F25 initially 12.2 ± 0.12 kg) were used in a 28-d study. Pens of pigs were allotted to 1 of 5 dietary treatments based on average BW and location within barn with 19 to 22 pigs per pen (with a similar number of barrows and gilts in each pen) and 12 replications per treatment in a randomized complete block design. Dietary treatments were arranged in a (2 × 2) + 1 factorial. The diet formaldehyde (1000 ppm) treatment was done using a commercially available product (3.2 kg/tonne, Sal CURB, Kemin Industries, Inc., Des Moines, IA) and crystalline AA inclusion were low and high. A positive control was used in the experiment to represent diets that met the assumed standardized ileal digestible (SID) Lys requirement estimate for pigs used in this study. Other treatment diets were formulated to be 80% of the SID Lys concentration in the positive control diet. Pigs and feeders were weighed to determine average daily gain (ADG), average daily feed intake (ADFI), and gain to feed ratio (G:F). The study was conducted at typical standard pig production facility in Iowa. Each pen (2.44 × 5.64 m) was equipped with a 4-hole, dry-self feeder and a pan waterer to provide ad libitum access to feed and water.
Feed sample collection
Feed samples were collected directly from each individual batch of feed in 5 spaced sub-samples by passing sterile Whirl-Pak (Nasco, Ft. Atkinson, WI) through the stream of feed as it was emptied from load-out bin into the feed delivery truck. Feed samples were also collected directly from 6 different feeders for each dietary treatment and placed in sterile Whirl-Pak bags to represent farm samples. Both mill and farm samples were pooled within collection location and transported to the Department of Diagnostic Medicine/Pathobiology, College of Veterinary Medicine, Kansas State University for feed bacterial enumeration analysis.
Feed bacteria enumeration
Feed samples were evaluated for bacterial counts using specialized plates (3 M Petri film 3 M Microbiology, St. Paul, MN) with each plate selecting for certain organisms. The specific organisms being detected for this experiment were: total coliforms (TC), aerobic plate counts (APC), and Enterobacteriaceae (EB). One gram of feed sample was diluted in 10 mL of phosphate buffered saline (PBS) tube and vortexed to make a uniform suspension before serially diluting the feed suspension to achieve 100, 10−1, 10−2, and 10−3 concentrations. With a sterile pipette, 1 mL of feed sample at each dilution was placed in the center of the Petrifilm in triplicates for each plate type. Dilutions were vortexed before plating to ensure equal distribution of feed inoculum. The sample inoculum was uniformly distributed over a 20 cm2 circular plate area. Total coliform, Enterobacteriaceae, and Aerobic plates were incubated for 48, 24, and 48 hrs, respectively. After incubation, colony number, colony morphology, gas production, and acidification for each plate was performed using a commercial plate reader (3 M Petrifilm, St. Paul, MN). Counting ranges were 50 to 150 colonies for coliform plates, 15 to 100 colonies for Enterobacteriaceae plates, and 25 to 250 colonies aerobic count plates. Colony counts were expressed as colony forming units per g of feed sample (cfu/g) and bacterial counts were expressed as an average of 2 separate runs ran in duplicate with a different feed sample.
Fecal sample collection
Fecal samples were collected via rectal massage into individual sterile bags from 6 randomly selected pigs on d 0 and from 3 randomly selected pigs per pen on d 28. Samples were stored at 4 °C and transported to Kansas State University where d 28 samples were pooled within pen into individual samples to represent 6 baseline samples and 12 samples per treatment. Samples were stored at −80 °C until transportation to the University of Nebraska-Lincoln for bacterial community analysis.
DNA Extraction and PCR Amplification
Fecal DNA (6 from baseline, 12 per treatment) were isolated from approximately 100 mg samples using a commercially available kit (Mag-Bind Soil DNA Kit; Omega Bio-tek, Inc., Norcross, GA) according to the manufacturer’s protocol with the following modifications: the raw sample was transferred to a 2 mL sterile Safe-Lock tube (Eppendorf, Hauppauge, NY) with 0.3 g of acid washed beads (Scientific Asset Management, Basking Ridge, NJ) and 300 µL of SLX-Mlus Buffer, followed by bead-beating at frequency of 20 for 10 min (QIAGEN Inc., Valencia, CA); after samples were mixed with RNase A and DS Buffer, samples were incubated in a 90 °C water bath for 8 min and occasionally vortexed; the samples were centrifuged at a speed of 5,000 \(\times \) g for 10 min and the supernatant was transferred; finally, DNA was eluted with 130 µL of elution buffer at the last step.
The V4 region of the 16S rRNA gene specific to the eubacterial communities was amplified from the extracted DNA samples28. A PCR reaction (20 µL) consisted of 2 µL of template DNA, 0.5 µL each of both forward and reverse 16S rRNA V4 primers (final concentration 10.0 µM; Integrated DNA Technologies, Coralville, IA), 0.5 µL of Terra PCR Direct Polymerase Mix (Clontech Laboratories Inc., Mountain View, CA), 10 µL of 2× Terra PCR Direct Buffer (Clontech Laboratories Inc., Mountain View, CA) and 6.5 µL of nuclease-free water (Hoefer Inc., Holliston, MA). Using a Veriti 96-well thermocycler (Life Technologies, Carlsbad, CA), the amplification was performed at 98 °C for 3 min, followed by 25 cycles of 30 s at 98 °C, 30 s at 55 °C, and 45 s at 68 °C, with a final elongation stage of 4 min at 68 °C. The amplified PCR products were tested by agarose gel electrophoresis (5 µL PCR product, 1.5% agarose gel) at 100 V for 60 min for size verification and to confirm amplification.
Preparation of 16S rRNA Library and sequencing
From each sample, 10 μL of the PCR product was pooled together and mixed. The pooled 16S rRNA gene library was column purified using PCR cleanup procedure (DNA, RNA, and protein purification; Clontech Laboratories, Mountain View, CA) and eluted into 40 µL. Subsequently, the Pippin Prep (Sage Science, Beverly, MA) was used to remove any spurious PCR fragments from the purified concentrated library. Finally, sequencing was performed using the Illumina MiSeq platform (Illumina, Santa Clara, CA) according to the manufacturer’s protocol.
Bacterial Community Analysis
The raw reads were demultiplexed using the Quantitative Insights Into Microbial Ecology program (QIIME29) and run through a quality control described by Anderson et al.30. Reads were trimmed to a fixed length of 251 bp using Mothur31 and the FASTX-TOOLKIT32, followed by identification of operational taxonomic unit (OTU) based on 97% similarity using UPARSE pipeline33. The generated OTU sequences were aligned using Ribosomal Database Project (http://pyro.cme.msu.edu). Taxonomic classification was performed using QIIME and the Green Genes database32 (version 13_8). The OTUs belonging to the phylum Cyanobacteria were removed as these are from dietary source. Single sequences that may be generated from sequencing error, were removed from the data. A core set of 854 OTUs (42.3% of the original OTU table), presenting in at least 75% of the samples, was identified for further analysis.
Fecal Microbial Diversity
Abundance in different taxonomy levels in bacterial community structure (β-diversity) were evaluated using normalized unweighted UniFrac distance matrices, with an OUT defined at an identity cut-off at 97% (14,163). The unweighted UniFrac distance matrices were used as input for a multivariate analysis using the Fathom Toolbox for MATLAB34.
Statistical Analyses
The AA content as represented as the Lysine concentration of the feed was evaluated using the analyzed value and the percent difference of analyzed values from expected formulated values. Feed bacterial concentration was analyzed by converting these values to log10. Pig growth performance as represented by ADG, ADFI and G:F were analyzed as a randomized complete block design using analysis of variance based on the assumption of normal distribution and heterogeneous variance. Pen was considered the experimental unit for growth performance and bacterial community analysis. Bacterial community analysis data were analyzed as OTU abundances at the family level. Fecal microbial diversity and the observed core OTU, Chao 1 and Shannon index from 10 subsampling events per fecal sample. Treatment was considered the fixed effect and a random effect of block. Residuals for each response criteria were evaluated for evidence of lack of normality and heterogeneous variance. Feed bacterial concentration had evidence of lack of normality and thus was log transformed and heterogeneous variance accounted for in the statistical model. No evidence for heterogeneous variance was noted for other criteria and thus variance was reported as pooled standard error of the mean. Pre-planned contrasts were utilized to compare the interaction between crystalline AA level and formaldehyde inclusion, the main effects of formaldehyde inclusion or crystalline AA level, and crystalline AA inclusion compared to the positive control. All data were analyzed using the GLIMMIX procedure of SAS 9.435. Results are reported as lsmeans +/− the standard error of the mean and were considered significant at P ≤ 0.05 and marginally significant at P > 0.05 and P ≤ 0.10.
References
Muhl, A. & Liebert, F. Growth and parameters of microflora in intestinal and fecal samples of piglets due to application of a phytogenic feed additive. J. Anim. Physiol. Anim. Nutr. 91, 411–418 (2007).
Zeyner, A. & Boldt, E. Effects of probiotic Enterococcus faecium strain supplemented from birth to weaning on diarrhea patterns and performance of piglets. J. Anim. Physiol. Anim. Nutr. 90, 25–31 (2006).
Anderson, K. E., Sheldon, B. W. & Richardson, K. E. Effect of termin-8 compound on the growth of commercial white and brown egg type pullets and environmental microbiological populations. Poult. Sci. 80, 88 (2001).
Food and Drug Administration (FDA). Code of Federal Regulations. 2015. Formaldehyde. 21CFR573.460 (2015).
Moller, J. Treating feeds with formaldehyde to protect protein. Feedstuffs 30, 12–13 (1983).
Dee, S. et al. An evaluation of porcine epidemic diarrhea virus survival in individual feed ingredients in the presence or absence of a liquid antimicrobial. Porcine Health Management. 1, 9, https://doi.org/10.1186/s40813-015-0003-0 (2015a).
Dee, S. et al. An evaluation of a liquid antimicrobial (Sal CURB®) for reducing the risk of porcine epidemic diarrhea virus infection of naïve pigs during consumption of contaminated feed. BMC Veterinary Research. 10, 220, https://doi.org/10.1186/s12917-014-0220-9 (2015b).
Cochrane, R. A. et al. Evaluating chemical mitigation of Porcine Epidemic Diarrhea virus in swine feed and ingredients. Proc. ADSA-ASAS 2015 Midwest Meeting (2015).
Metz, B. et al. Identification of formaldehyde-induced modifications in proteins reactions with model peptides. J. Bio. Chem. 279, 6235–6243, https://doi.org/10.1074/jbc.M310752200 (2004).
McCormack, U. M., et al. Exploring a possible link between the intestinal microbiota and feed efficiency in pigs. Appl. Environ. Microbiol. 83 (15), e00380-17, https://doi.org/10.1128/AEM.00380-17 (2017).
Lamendella, R. et al. Comparative fecal metagenomics unveils unique functional capacity of the swine gut. BMC Microbiol. 11, 103, https://doi.org/10.1186/1471-2180-11-103 (2011).
Vigors, S. et al. Pigs that are divergent in feed efficiency, differ in intestinal enzyme and nutrient transporter gene expression, nutrient digestibility and microbial activity. Animal 10, 1848–1855, https://doi.org/10.1017/S1751731116000847 (2016).
Carrique-Mas, J. J., Bedford, S. & Davies, R. H. Organic acid and formaldehyde treatment of animal feeds to control Salmonella: efficacy and masking during culture. J. App. Micro. 103, 88–96, https://doi.org/10.1111/j.1365-2672.2006.03233.x (2007).
DeRouchey, J. M. et al. Evaluation of methods to reduce bacteria concentrations in spray-dried animal plasma and its effects on nursery pig performance. J. Anim. Sci. 82, 250–261, https://doi.org/10.2527/2004.821250x (2004).
Sbardella, M. et al. Effects of a dietary added formaldehyde-propionic acid blend on feed enterobacteria counts and on growing pig performance and fecal formaldehyde excretion. Ciência Rural. 45, 474–479, https://doi.org/10.1590/0103-8478cr20131660 (2015).
Rowghani, E., Arab, M. & Akbarian, A. Effects of a probiotic and other feed additives on performance and immune response of broiler chicks. Int. J. of Poult. Sci. 6, 261–265, https://doi.org/10.3923/ijps.2007.261.265 (2007).
European Food Safety Authority (EFSA). Scientific opinion on the safety and efficacy of formaldehyde for all animal species based on a dossier submitted by Regal BV. EFSA Journal 12, 3561 (2014).
French, D. & Edsall., J. T. The reactions of formaldehyde with amino acids and proteins. Adv. Pro. Chem. 2, 277–335, https://doi.org/10.1016/S0065-3233(08)60627-0 (1945).
Rude, C., Mellick, D., Lamptey, A. & Bienhoff, M. Evaluation of the effects of a formaldehyde-based feed additive on free lysine. J. Anim. Sci. 94, 135, https://doi.org/10.2527/msasas2016-288 (2016).
Ochoa, L. et al. Effect of feeding formaldehyde-treated feed to pigs throughout the growing period on amino acid utilization from crystalline lysine or protein sources. J. Anim. Sci. 95, 142, https://doi.org/10.2527/asasmw.2017.294 (2017).
Brestoff, J. R. & Artis, D. Commensal bacteria at the interface of host metabolism and the immune system. Nat. Imm. 14, 676–684, https://doi.org/10.1038/ni.2640 (2013).
Adams, R. I., Bateman, A. C., Bik, H. M. & Meadow, J. F. Microbiota of the indoor environment: a meta-analysis. Micro 3, 49, https://doi.org/10.1186/s40168-015-0108-3 (2015).
Holman, D. B., Brunelle, B. W., Trachsel, J. & Allen, H. K. Meta-analysis To Define a Core Microbiota in the Swine Gut. mSys. 2, e00004–17, https://doi.org/10.1128/mSystems.00004-17 (2017).
Levesque, C. L., Hooda, S., Swanson, K. S. & de Lange, K. Alterations in ileal mucosa bacteria related to diet complexity and growth performance in young pigs. PloS. 9, e108472, https://doi.org/10.1371/journal.pone.0108472 (2014).
Looft, T. et al. Bacteria, phages and pigs: the effects of in-feed antibiotics on the microbiome at different gut locations. ISME. 8, 1566, https://doi.org/10.1028/ismej.2014.12 (2014).
Mann, E. et al. Mucosa-associated bacterial microbiome of the gastrointestinal tract of weaned pigs and dynamics linked to dietary calcium-phosphorus. PLoS. 9, e86950, https://doi.org/10.1371/journal.pone.0086905 (2014).
Owens, R. C. Jr. et al. Antimicrobial-associated risk factors for Clostridium difficileinfection. Clin. Inf. Dis. 46, S19–S31, https://doi.org/10.1086/521859 (2008).
Kozich, J. J. et al. Development of a dual-index sequencing strategy and curation pipeline for analyzing amplicon sequence data on the MiSeq Illumina sequencing platform. Appl. Environ. Microbiol. 79(15), 5112–5120, https://doi.org/10.1128/AEM.01043-13 (2013).
Caporaso, J. G. et al. Ultra-high-throughput microbial community analysis on the illumina HiSeq and MiSeq platforms. ISME J. 6, 1621–1624, https://doi.org/10.1038/ismej.2012.8 (2012).
Anderson, C. L., Schneider, C., Erickson, G., MacDonald, J. & Fernando, S. C. Rumen bacterial communities can be acclimated faster to high concentrate diets than currently implemented feedlot programs. J. Appl. Microbiol. 120, 588–599, https://doi.org/10.1111/jam.13039 (2016).
Schloss, P. D. et al. Introducing mothur: Open-source, platform-independent, community-supported software for describing and comparing microbial communities. Appl. Environ. Microbiol. 75, 7537–7541, https://doi.org/10.1128/AEM.01541-09 (2009).
Edgar, R. C. UPARSE: Highly accurate OTU sequences from microbial amplicon reads. Nature Methods. 10, 996–998, https://doi.org/10.1038/nmeth.2604 (2013).
McDonald, D. et al. An improved green genes taxonomy with explicit ranks for ecological and evolutionary analyses of bacteria and archaea. ISME J. 6, 610–618, https://doi.org/10.1038/ismej.2011.139 (2012).
Jones, D. The fathom toolbox for MATLAB: Software for multivariate ecological and oceanographic data analysis. College of Marine Science, University of South Florida, Petersburg, FL (2015).
SAS Institute Inc., Cary, NC, USA (2018).
Acknowledgements
We thank Kemin Industries (Des Moines, IA) and PIC (Hendersonville, TN) for technical and financial support towards the experiment. Appreciation is expressed to Pillen Family Farms (Columbus, NE) for assistance in manufacturing of diets. Authors also thank Mal Hoover for her help with the image preparations.
Author information
Authors and Affiliations
Contributions
H.E.W., J.C.W., J.M.D., S.S.D., M.D.T., C.K.J., R.G.A., and R.D.B. conceived and designed the study. H.E.W. and R.A.C. oversaw the diet design and feed manufacturing and animal work. S.C.F., T.E.B. and S.L. performed microbiome analyses. R.G.A. performed the bacterial enumeration. All authors contributed to the manuscript writing and approved the final version to be published.
Corresponding author
Ethics declarations
Competing Interests
The authors declare no competing interests.
Additional information
Publisher's note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Williams, H.E., Cochrane, R.A., Woodworth, J.C. et al. Effects of dietary supplementation of formaldehyde and crystalline amino acids on gut microbial composition of nursery pigs. Sci Rep 8, 8164 (2018). https://doi.org/10.1038/s41598-018-26540-z
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-018-26540-z
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
