
Abstract
Feeding preference of fossil herbivorous mammals, concerning the coevolution of mammalian and floral ecosystems, has become of key research interest. In this paper, phytoliths in dental calculus from two gomphotheriid proboscideans of the middle Miocene Junggar Basin, Central Asia, have been identified, suggesting that Gomphotherium connexum was a mixed feeder, while the phytoliths from G. steinheimense indicates grazing preference. This is the earliest-known proboscidean with a predominantly grazing habit. These results are further confirmed by microwear and isotope analyses. Pollen record reveals an open steppic environment with few trees, indicating an early aridity phase in the Asian interior during the Mid-Miocene Climate Optimum, which might urge a diet remodeling of G. steinheimense. Morphological and cladistic analyses show that G. steinheimense comprises the sister taxon of tetralophodont gomphotheres, which were believed to be the general ancestral stock of derived “true elephantids”; whereas G. connexum represents a more conservative lineage in both feeding behavior and tooth morphology, which subsequently became completely extinct. Therefore, grazing by G. steinheimense may have acted as a behavior preadaptive for aridity, and allowing its lineage evolving new morphological features for surviving later in time. This study displays an interesting example of behavioral adaptation prior to morphological modification.
Similar content being viewed by others

Introduction
The coevolution of plants and mammals amid dramatic Cenozoic climactic perturbations has been the focus of intensive research1,2. There is now abundant evidence to suggest that grassland ecosystems dominated by the family Gramineae expanded markely in the Neogene, and promoted significant adaptive evolutionary responses in various herbivorous mammal lineages2. For example, proboscideans, the largest Neogene herbivores which were ‘super-keystone species’ in their ecosystems3, modified their grinding teeth from low-crowned bunolophodont (“mastodont-like”) molars with few loph(id)s (enamel cones fused in a line), to high-crowned and multi-plated (“elephant-like”) teeth allowing them to consume more abrasive foods4. At least for proboscideans, several analytical approaches have been adopted to track this process, such as enamel microwear3,5 and stable isotope studies6. However, any single given palaeodietary proxy per se is insufficient to provide extensively detailed inferences about the feeding ecology of fossil organisms. Therefore, a multi-proxy approach is essential for performing palaeodietary investigations.
Dental calculus is the calcium phosphate deposited on teeth, and captures a large number of food particles that provide crucial information about the food of ancient animals7,8. Phytoliths sealed within the calculus, in particular, provide an extensive dietary record9,10,11. Due to their strong resistance to decay and to mechanical or biological decomposition, phytoliths can be well preserved for a long time12. Phytoliths have been extracted from long-dead animals that span a great temporal scope, from ancient humans8 to an Early Cretaceous dinosaur13. A wide range of phytoliths have also recently been recovered from the dental calculus of Pleistocene proboscideans, such as Notiomastodon14 and Mammut15. Because of their consistent shape within different plant taxa, phytoliths therefore provide a very strong taxonomic signal12,16. With diagnosability to the family, genus or even species level, phytoliths thus provide a reliable basis to explore the feeding ecology and palaeoenvironments of ancient animals, beyond the level of resolution available from other palaeodietary proxies such as dental microwear and stable isotopes, which generally provide broad ‘C3/C4’ inferences (which themselves act as proxy for ‘browsing/grazing’) for herbivores, although these proxies provide useful complementary data17.
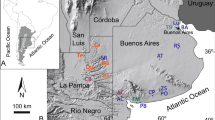
In the present article, we examine the feeding ecology of two species of gomphotheriid proboscideans, Gomphotherium connexum and G. steinheimense, based on phytoliths from their dental calculus, complemented by dental stereomicrowear and stable isotopic data. Our studied samples are discovered from the Miocene Halamagai Formation, northern Junggar Basin, Xinjiang, China (Figs 1, S1). Previous studies and our pollen data have shown that the Halamagai Formation covers the Mid-Miocene Climate Optimum (MMCO)18 and records the major palaeoenvironmental perturbation crises from a more humid Middle Miocene environment to the comparatively more arid Late Miocene ecosystem19,20 (SI 1.1). This palaeoenvironmental transition had a profound impact on the evolution of terrestrial floras and faunas in Central Asia and elsewhere. Our gomphothere dietary reconstruction indicates the disparate feeding habits of the two Gomphotherium species from Halamagai Formation. Furthermore, cladistic analysis also indicates the distinct phylogenetic affinities of the two species (Fig. 1C, SI 1.2, 1.3). Therefore, our research is of importance not only in understanding palaeoenvironmental changes in the middle-latitude zone of inland Asia during the Middle to Late Miocene, but also holds broader significance in understanding the coevolution of plant communities and mammals in terms of habitats and feeding ecology.

Geography, geology, and phylogeny in relation to the study material. (A) The location of the study area (black star), details are shown in Fig. S1. The map was generated by GTOPO309 (http://www1.gsi.go.jp/geowww/globalmap-gsi/gtopo30/gtopo30.html) using Globalmapper (v10) (http://www.bluemarblegeo.com/products/global-mapper.php). (B) Stratigraphic column and polarity with palaeomagnetic age (redrawn after refs18,19), also denoting the horizon of study material in the strata (in light yellow). (C) The 50% majority consensus tree from 29 maximum parsimonious trees showing the phylogenetic position of the Gomphotherium species and Tetralophodon longirostris (tree length = 81, CI = 0.617; RI = 0.744, data set in Table S1), the number at each node representing the support value calculated by majority rules (percentages of supported MPTs in the total MPTs, which are always larger than 50%) and the orange frame indicating the sister-taxon relationship of G. steinheimense and T. longirostris. (D) G. steinheimense, IVPP V23283, right m3. (E) G. connexum, IVPP V8576, left M3.
Results
Phytoliths analysis
Our method for extracting phytoliths from dental calculus was successful for four molars of G. connexum and two of G. steinheimense (see the Materials and Methods section). The samples contain grass phytolith morphotypes including several types of short cells and long cells (Table 1 and Fig. 2A–K). The main forms of grass short cell phytoliths are bilobate short cells, bulliform, rondel, scutiform-bulliform and square/rectangular. Meanwhile, other morphotypes including hair cell/hair base and irregular multifaceted phytoliths were also extracted; these are found primarily in eudicots leaf based on the available phytolith literature12,21. In addition, other phytolith types without taxonomic significance, such as smooth elongate and elongate echinate, are also present. Besides, some sponge spicules were also found in the Gomphotherium specimens (Fig. 2L, SI 1.4).

Phytolith and sponge spicules, as well as their statistics in Gomphotherium dental calculus. (A–L) Morphology of various phytolith, including bilobates short cell (V8573) (A), bilobates short cell (V23283) (B), bulliform (V8575) (C), bulliform (V23283) (D), rondel (V23283) (E), rectangular (V8576) (F), reed-type bulliform (V23283) (G), long cell (V8575) (H), hair cell (V8576) (I), irregular multifaceted (V8573) (J), elongate (V8575) (K) sponge spicules (V8576) (L). (M) Percentages of grass phytoliths, eudicots phytoliths, unclassed phytoliths and sponge spicules occurring in Gomphotherium dental calculus. (N) Example of dental calculus sampling (the tooth is V8576).
Phytolith types exacted from the dental calculus of four studied G. connexum specimens are consistent. They have relatively high grass phytolith concentration (40–50%). Meanwhile eudicot phytoliths (28–34%) are also consistently present in these four samples (Fig. 2M). Eudicots produce considerably fewer phytoliths on average than grasses12, so the presence of eudicot leaf phytoliths very likely indicate that eudicot foliage was a staple dietary component for G. connexum from our studied area. In other words, our phytolith analysis indicates that G. connexum was a mixed-feeding generalist that fed on both eudicot foliage and grass, or even primarily a browser.
By contrast, the percentage of grass phytoliths (around 85% including several types of short cells and long cells) in the dental calculus of G. steinheimense is much higher. Bilobate short cell phytoliths from panicoid grasses (subfamily Panicoideae) are the most common type of short cell12,21, comprising around 25% of total phytoliths. Other grass phytoliths include abundant common bulliforms (around 20%) and square/rectangular (around 15%), rondels from pooid grasses12,21 (subfamily Pooideae) (around 9%). Long cells (around 4%) were exacted in lesser quantities (Table 1 and Fig. 2M). Large quantities of grass phytoliths among this sample suggest that grass occupied a significant dietary component in the two examined individual of G. steinheimense from our present study. Moreover, no eudicot phytoliths are present in this sample (Fig. 2M). The result indicates that the G. steinheimense individuals examined in this study were grazers, rather than browsers.
Microwear analysis
A stereomicrowear analysis of gomphothere dental enamel was performed to further validate the phytolith results. Consistently, the G. connexum sample displays relatively sparse configuration of thick scratches with scattered pits of irregular size, through micrographs of the molar shearing surfaces (Fig. 3A–C). On the other hand, the G. steinheimense sample shows dense thin scratches with two (or sometimes more) predominant orientations, which shadow the pits (Fig. 3D). This corroborates the disparity of the feeding habits in the two Gomphotherium species indicated by the aforementioned phytolith analyses. The average number of scratches in the studied G. steinheimense molars are significantly larger than any of the G. connexum specimens examined in this study (Table S2). The G. steinheimense samples fall into the area of grazers, whereas G. connexum from Halamagai is indicated as browser or mixed feeder (Figs 3, S2). This result draws consistent interpretations with respect to the aforementioned phytolith analyses.

Bivariate plot of the average scratch versus average pit counts in enamel microwear of G. connexum, G. steinheimense, and extant elephants (redrawn after ref.38). Oval outlines = Gaussian confidence ellipses (p = 0.95) on the centroid of the comparative extant grazer and browser samples adjusted by sample size. The insert panels represent photomicrographs of enamel surfaces of G. connexum (A–C) and G. steinheimense (D).
Stable isotope analysis
The δ13C values of these six samples range from −10.9‰ to −8.3‰, whereas δ18O values are between −13.1‰ and −7.5‰ (Fig. 4, Table S3). Considering the isotopic enrichment (14.1‰) from the diet to bioapatite in large herbivores6, the δ13C values of all Gomphotherium individuals here, averaged by −8.8 ± 0.3‰ (n = 6), show that they consumed mainly C3 plants. The relatively small variation of carbon isotope values shows that the Gomphotherium species may have had broadly similar habitat preferences, and inhabited a largely homogeneous ecosystem. The relatively high mean δ13C value (−8.8‰) of these animals suggests that they probably inhabited a considerably open environment where the C3 grasses with higher δ13C values were grown22. A greater large variability of δ18O values, averaged by −10.2 ± 2.2‰ (n = 6), are detected (Fig. 4), suggesting that G. connexum and G. steinheimense from the Halamagai Formation may have exploited different sources of water intake, or fed upon plants with different δ18O values22,23,24,25. Nevertheless, considering the fact that the fossiliferous horizon which yielded the Gomphotherium fossils here is probably the product of gradual sedimentation over a very long period, during which large climatic fluctuations could have occurred. On the other hand, given the fact that no other animals were used here as isotopic baseline for habitat differentiation, it is not meaningful to discuss the oxygen isotopic variation alone between the two species with unequal sample numbers.

Scatter plot of carbon and oxygen isotope values of the Gomphotherium samples.
Cladistic Analysis
A cladistic analysis was performed to investigate the possible phylogenetic relationships of different Gomphotherium species. The data matrix contains 52 binary characters and 19 terminals, including three outgroups (Table S1, SI 6.1). Characters 0–49 used follow those of Wang et al.26, whereas characters 50 and 51 are novel characters incorporated in the present study (Table S1). Cladograms were obtained from a maximum parsimony analysis carried out using the TNT1.1 program with the ‘traditional research’ option27. The reported results and node supports were calculated from a 50% majority consensus tree (Fig. 1C).
Pollen analysis
We analysed a total of 34 pollen samples, among which 21 samples had a relative high pollen concentration, and 13 samples failed to obtain 50 pollen grains or more. A total of 2278 pollen grains were obtained from the 21 samples, 108 in average per sample, consisting of 48 genera and species were counted and identified (Fig. S1 and SI 6.2).
Discussion
Gomphotherium plays an essential role in exploring the evolution of elephantiform proboscideans, as the genus has long been considered to represent the broad distant ancestral stock to more derived elephantiforms, including the extant elephantids or “true elephants”, via the intermediate “tetralophodont gomphothere” grade4,28. Tetralophodonts possess at least one more loph(id) on each molar than the trilophodont Gomphotherium, an possible adaptation for processing more abrasive foodstuff4. Further comparison of enamel stereomicrowear from G. steinheimense and several tetralophodont gomphotheres (Tetralophodon xiaolongtanensis, Parateralophodon sp., and Anancus sinensis) underpins this hypothesis (Fig. S2). An exception can be found in the stegodontid Stegolophodon stegodontoides, which appears to be a mixed feeder (Fig. S2). The stegodontid family are another fully lophodont radiation of proboscideans that originated from the tetralophodonts, in parallel with elephantids28. Stegolophodon is believed to be ancestral to the Plio-Pleistocene Stegodon, both genera characterised by brachyodont (low-crowned) molars29, and thought to have fed upon softer foodstuff than the contemporary true elephantids, as a means of niche partition5.
Within Gomphotherium, our cladistic analysis suggests that G. steinheimense is the likely sister taxon to Tetralophodon longirostris, the archetypical tetralophodont gomphothere. Whereas G. connexum, the other trilophodont gomphothere from the Halaimagai Formation, is nested as the sister species to the type species G. angustidens (Fig. 1C). Palaeomagnetic studies indicate that the Halamagai Formation approximately covers the 17–15 Ma age range, corresponding to the Mid-Miocene Climatic Optimum (MMCO)18,19,20 (Fig. 1B and SI 1.1). Subsequently, a decrease of gomphothere diversity took place in northern China during the Late Miocene30, which saw the complete replacement of trilophodont gomphotheres, including Gomphotherium, by tetralophodont gomphotheres (tri-/tetralophodont replacement event). To a considerable extent, this major turnover has been attributed to the severe aridification of the Asian interior31. The Halamagai Formation thus provides a remarkable opportunity to understand the palaeoecological backdrop to this major proboscidean turnover. Our phytolith and microwear results strongly indicate that the two Gomphotherium species from the Halamagai Formation show a potential partitioning of feeding ecology. The appreciable presence of eudicot phytoliths from our G. connexum samples suggests it was an obligate browser or mixed feeder, similar to other Gomphotherium species from Europe and North America32,33. Whereas more unexpectedly, the two studied specimens of G. steinheimense may have had a more grass-dominated feeding preference.
The MMCO has previously been detected in deposits from the southern part of the Junggar Basin. Pollen data from the Jingou River section suggests a rapid recovery of regional woodland during 17.3–16.2 Ma34. However, our pollen record from the Gomphotherium-bearing horizon (the lower part of the Halamagai Formation) reveals that the environment was a grass- and Artemisia-dominated prairie accompanied by few arboreal taxa (Fig. S3, SI 2.5, 6.2). Furthermore, in the upper part of the Halamagai Formation and the immediately overlying Kekemaideng Formation, an increase of drought-tolerance shrubs, such as Ephedra and Tamarix, reveals that prairie had been replaced by a goose-foot-dominated semi-arid savannah (Fig. S3 and SI 6.2). These pollen data thus indicate probable aridity in the northern Junggar Basin during MMCO. This establishment of a grassland-dominated ecosystem may have driven some large herbivores such as G. steinheimense to modify their feeding behaviour, and become opportunistic to obligate grazers.
The end of the Middle Miocene saw the near-complete extinction of Gomphotherium in northern China and the rest of Central Asia30, with the exception of G. steinheimense. Unfortunately, the prerequisite circumstances available for the present study restricts the sample size of G. steinheimense materials analysed hereby to two specimens, thereby precluding more detailed inferences about the feeding ecology of this species in Middle Miocene Halamagai. However, it is worth noting that G. steinheimense from the Middle/Late Miocene of Southern Germany has been demonstrated to be a catholic mixed-feeder of both grass and foliage33. The modern elephants have long been held as considerably more committed grazer than the gomphotheres, with their molar loph(id)s modified into numerous tightly packed lamellae and considerably higher molar crown (hypsodonty)4,35. Nonetheless, empirical studies of elephant feeding behaviour have instead shown the living elephant species as highly generalist megaherbivores, with wild populations typically feeding on over 100 different plant species which span the whole browsing-grazing spectrum, depending on local habitat and seasonality36.
As aforementioned, G. steinheimense quite possibly represents an ancestral lineage to tetralophodont gomphotheres (Fig. 1C), which in turn form the immediate stem grade to elephantids4,28,36. Combining this phylogenetic framework with our understanding of past and present proboscidean feeding ecology, it is hereby reasonable to hypothesise that the first evolutionary steps towards the strongly eurytopic feeding ecology seen in modern elephants36 was first made by G. steinheimense, a behavioural tendency which facilitates this species to better exploit a newly abundant food source in the form of grass. On the other hand, more evolutionarily conservative gomphotheres, such as G. connexum, may have had less flexible feeding preferences to adapt to the significant vegetational turnover in the Middle/Late Miocene, which resulted in the expansion of more open, grassy habitats.
A classic narrative in the evolution of large herbivorous mammals during the Cenozoic is that hypsodonty appeared in various herbivore lineages in response to the spread of grasslands towards the late Neogene, in order to feed on more abrasive vegetation17,37. However, Lister4,37 noted the prominent temporal lag between a major shift from browsing to grazing in African proboscideans (these include putative direct ancestors of elephantids) (10 Ma) and a significant evolutionary burst towards hypsodonty (5–0.5 Ma). Our Middle Miocene (17–15 Ma) record of a predominantly grazing Gomphotherium thus further extends this lag, and supports the hypothesis that within the tetralophodont-elephantid lineage, other modifications to dental morphology (increase in the number of molar loph[id]s and multiplication in the number of conelets on the occlusal facet of each loph[id]) were taking place as a subsequent adaptive response during the lag (Fig. S2)4,37. Furthermore, it is worth noting that the fossil record points to the likely origin of tetralophodont gomphotheres in Central Asia at the 18–16 Ma interval28, the geochronological context of our study on the feeding ecology of Gomphotherium from Halamagai is thus also in immediate correspondence with the origin of the tetralophodont-elephantid lineage, in addition to phylogenetic aspect.
In sum, our present study reveals that the marked Neogene dietary transition from browsing to grazing in the elephantid stem lineage4,37 is a process that may be linked back to G. steinheimense from the MMCO of Central Asia (Fig. S2), at the origin of the tetralophodont-elephantid line. Reconciliation with our current understanding of dietary evolution in elephantimorphs suggests grazing quite probably began through a combination of eurytopic foraging habits35,36 and the global expansion of grasslands which began towards the Late Miocene22,34, which then led to the suite of grazing-related morphological adaptations which dominated the later evolution and radiation of proboscideans4,5,28,35,36. Our study thus further enforces the emergence of grazing in proboscideans and their subsequent adaptive morphological evolution as a crucial case for demonstrating the importance of decoupling the origination of behavioural and phenetic novelties in palaeobiology4,37.
Materials and Methods
Data availability
The specimens on which this study is based is housed in the collections of the Institute Vertebrate Paleontology and Paleoanthropology (IVPP), Chinese Academy of Sciences. The txt format file for the maximum parsimony analysis (SI 6.1) and xlsx format file for the pollen data (SI 6.2) are available on DRYAD.
Specimens
Six molars of Gomphotherium have been examined, including five specimens of G. connexum (IVPP V8573–8576, and V18701) (Figs 1E and S4A–C), and one complete m3 of G. steinheimense (IVPP V23283) (Figs 1D and S4D). Another tooth fragment was referred to as? G. steinheimense (IVPP V24891), because it displays the diagnostic morphology of G. steinheimense, details were shown in SI 1.1 (Fig. S4E,F). These specimens are the total Gomphotherium sample so far from the study area. Therefore our sample includes most of the available data at the present stage.
Methods
Dental calculus was carefully removed from the tooth valleys of the study molars, preventing contamination (Figs 2N, S3 and SI 2.1). Phytolith extraction, stereomicrowear analysis, stable isotope study, and pollen extraction were performed using standard methods (SI 2.2, 2.3, 2.4). Cladistic analysis was carried out using TNT1.1 program.
References
Fortelius, M. et al. Evolution of Neogene mammals in Eurasia: environmental forcing and biotic interactions. Ann. Rev. Earth Pl. Sc. 42, 579–604 (2014).
MacFadden, B. J. Cenozoic mammalian herbivores from the Americas: Reconstructing ancient diets and terrestrial communities. Ann. Rev. Ecol. System 31, 33–59 (2000).
Calandra, I., Göhlich, U. B. & Merceron, G. How could sympatric megaherbivores coexist? Example of niche partitioning within a proboscidean community from the Miocene of Europe. Naturwissenschafen 95, 831–838 (2008).
Lister, A. M. The role of behaviour in adaptive morphological evolution of African proboscideans. Nature 500, 331–334 (2013).
Zhang, H., Wang, Y., Janis, C. M., Goodall, R. H. & Purnell, M. A. An examination of feeding ecology in Pleistocene proboscideans from southern China (Sinomastodon, Stegodon, Elephas), by means of dental microwear texture analysis. Quat. Int. 445, 60–70 (2017).
Cerling, T. E., Harris, J. M. & Leakey, M. G. Browsing and grazing in elephants: the isotope record of modern and fossil proboscideans. Oecologia 120, 364–374 (1999).
Lieverse, A. R. Diet and theaetiology of dental calculus. Internat. J. Osteoarchaeol. 9, 219–232 (1999).
Henry, A. G., Brooks, A. S. & Piperno, D. R. Microfossils in calculus demonstrate consumption of plants and cooked foods in Neanderthal diets (Shanidar III, Iraq; Spy I and II, Belgium). Proc. Natl. Acad. Sci. USA 108, 486–491 (2011).
Ciochon, R. L., Piperno, D. R. & Thompson, R. G. Opal phytoliths found on the teeth of the extinct ape Gigantopithecus blacki: Implications for paleodietary studies. Proc. Natl. Acad. Sci. USA 87, 8120–8124 (1990).
Hardy, K. et al. Neanderthal medics? Evidence for food, cooking, and medicinal plants entrapped in dental calculus. Naturwissenschaften 99, 617–626 (2012).
Hardy, K. et al. Diet and environment 1.2 million years ago revealed through analysis of dental calculus fromEurope’s oldest hominin at Sima del Elefante, Spain. Sci. Nat. 104, 1–5 (2016).
Piperno, D. R. Phytoliths: A Comprehensive Guide for Archaeologists and Paleoecologists (Alta Mira Press, Lanham, 2006).
Wu, Y., You, H. & Li, X. Dinosaur-associated Poaceae epidermis and phytoliths from the Early Cretaceous of China. Natl. Sci. Rev. https://doi.org/10.1093/nsr/nwx145 (2017).
Asevedo, L., Winck, G. R., Mothé, D. & Avilla, L. S. Ancient diet of the Pleistocene gomphothere Notiomastodon platensis (Mammalia, Proboscidea, Gomphotheriidae) from lowland mid-latitudes of South America: Stereomicrowear and tooth calculus analyses combined. Quat. Int. 255, 42–52 (2012).
Gobetz, K. E. & Bozarth, S. R. Implications for late Pleistocene mastodon diet from opal phytoliths in tooth calculus. Quat. Res. 55, 115–122 (2001).
Bozarth, S. R. In Phytolith Systematics (eds Rapp, Jr G. & Mulhollandand, S. C.) 193–214 (Plenum, New York, 1992).
Damuth, J. & Janis, C. M. On the relationship between hypsodonty and feeding ecology in ungulate mammals, and its utility in palaeoecology. Biol. Rev. 86, 733–758 (2011).
Sun, J.-M. et al. Late Oligocene–Miocene mid-latitude aridification and wind patterns in the Asian interior. Geology 38, 515–518 (2010).
Ye, J. et al. The Duolebulejin Section of northern Junggar Basin and its stratigraphic and environmental implication. Sci. China Earth Sci. 42, 1523–1532 (2012).
Zachos, J., Pagani, M., Sloan, L., Thomas, E. & Billups, K. Trends, rhythms, and aberrations in global climate 65 Ma to present. Science 292, 686–693 (2001).
Lu, H. Y. et al. Phytoliths as quantitative indicators for the reconstruction of past environmental conditions in China, I: Phytolith-based transfer functions. Quat. Sci. Rev. 25, 945–959 (2006).
Cerling, T. E. & Harris, J. M. Carbon isotope fractionation between diet and bioapatite in ungulate mammals and implications for ecological and paleoecological studies. Oecologia 120, 347–363 (1999).
Quade, J., Cerling, T. E., Andrews, P. & Alpagut, B. Paleodietary reconstruction of Miocene faunas from Paşalar, Turkey, using stable carbon and oxygen isotopes of fossil tooth enamel. J. Human Evol. 28, 373–384 (1995).
Sponheimer, M. & Lee-Thorp, J. A. Oxygen isotopes in enamel carbonate and their ecological significance. J. Archaeolog. Sci. 26, 723–728 (1999).
Koch, P. L., Fisher, D. C. & Dettman, D. Oxygen isotope variation in the tusks of extinct proboscideans: A measure of season of death and seasonality. Geology 17, 515–519 (1989).
Wang, S.-Q. et al. A new species of Gomphotherium (Proboscidea, Mammalia) from China and the evolution of Gomphotherium in Eurasia. J. Vertebr. Paleontol. 37, 1–15 (2017).
Goloboff, P. A., Farris, J. S. & Nixon, K. C. TNT, a free program for phylogenetic analysis. Cladistics 24, 774–786 (2008).
Gheerbrant, E. & Tassy, P. L’origine et l'évolution des éléphants. C. R. Palevol. 8, 281–294 (2009).
Saegusa, H., Thasod, Y. & Ratanasthien, B. Notes on Asian stegodontids. Quat. Int. 126, 31–48 (2005).
Wang, S.-Q. et al. Yunnan, a refuge for trilophodont proboscideans during the late Miocene aridification of East Asia. Palaeogeogr. Palaeoclimatol. Palaeoecol. http://dx.doi.org/10.1016/j.palaeo.2017.07.034 (2017).
An, Z. S., Kutzbach, J. E., Prell, W. L. & Porter, S. C. Evolution of Asian monsoons and phased uplift of the Himalaya-Tibetan plateau since Late Miocene times. Nature 411, 62–66 (2001).
Fox, D. L. & Fisher, D. C. Dietary reconstruction of Miocene Gomphotherium (Mammalia, Proboscidea) from the Great Plains region, USA, based on the carbon isotope composition of tusk and molar enamel. Palaeogeogr. Palaeoclimatol. Palaeoecol. 206, 311–335 (2004).
Calandra, I., Göhlich, U. B. & Merceron, G. Feeding preferences of Gomphotherium subtapiroideum (Proboscidea, Mammalia) from the Miocene of Sandelzhausen (Northern Alpine Foreland Basin, southern Germany) through life and geological time: Evidence from dental microwear analysis. Paläontol. Z. 84, 205–215 (2010).
Tang, Z. et al. Late Cenozoic central Asian drying inferred from a palynological record from the northern Tian Shan. Earth Planet Sci. Lett. 302, 439–447 (2011).
Savage, R. J. G. & Long, M. R. Mammal Evolution: an Illustrated Guide (Facts On File Publications, Oxford, 1986).
Sukumar, R. The Living Elephants: Evolutionary Ecology, Behaviour, and Conservation. (Oxford University Press, Oxford, 2003).
Lister, A. M. Behavioural leads in evolution: evidence from the fossil record. Biol. J. Linnean Soc. 112, 315–331 (2014).
Semprebon, G. M., Deng, T., Hasjanova, J. & Solounias, N. An examination of the dietary habits of Platybelodon grangeri from the Linxia Basin of China: Evidence from dental microwear of molar teeth and tusks. Palaeogeogr. Palaeoclimatol. Palaeoecol. 457, 109–116 (2016).
Acknowledgements
We thank A. M. Lister (The Natural History Museum, London, UK) for many kindly suggestions on this manuscript, and P. Dios (Museo Nacional de Ciencias Naturales Madrid, Spain) for preparation of specimens of Tetralophodon longirostris. We also thank M Fortelius (University of Helsinki) and an anonymous reviewer for the review of the manuscript. This work was funded by the National Natural Science Foundation of China (41472145, 41372001, 41625005 and 41430102), the Strategic Priority Cultivating Research Program, CAS (Grant No. XDA 20070203, XDPB05), National Basic Research Program of China (2015CB953803), and the Youth Innovation Promotion Association of Chinese Academy of Sciences (2018099) supported this project.
Author information
Authors and Affiliations
Contributions
Y.W., T.D. and S.W. designed research. Y.W., Y. H., J.M. X.Z., J.Y., L. M. and S.W. conducted research and analysed data. Y.W., H.Z. and S.W. prepared the paper.
Corresponding author
Ethics declarations
Competing Interests
The authors declare no competing interests.
Additional information
Publisher's note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Wu, Y., Deng, T., Hu, Y. et al. A grazing Gomphotherium in Middle Miocene Central Asia, 10 million years prior to the origin of the Elephantidae. Sci Rep 8, 7640 (2018). https://doi.org/10.1038/s41598-018-25909-4
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-018-25909-4
This article is cited by
-
Fluctuating climate and dietary innovation drove ratcheted evolution of proboscidean dental traits
Nature Ecology & Evolution (2023)
-
The anthracotheres from northern Junggar Basin and their palaeoclimatic significance in relation to the Tibetan Plateau
Palaeobiodiversity and Palaeoenvironments (2021)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
